
Abstract
Flu vaccines are partially protective in infants and elder people. New adjuvants such as immunostimulatory oligonucleotides (ODNs) are strong candidates to solve this problem, because a combination with several antigens has demonstrated effectiveness. Here, we report that IMT504, the prototype of a major class of immunostimulatory ODNs, is a potent adjuvant of the influenza vaccine in young adult and elderly rats. Flu vaccines that use virosomes or whole viral particles as antigens were combined with IMT504 and injected in rats. Young adult and elderly animals vaccinated with IMT504-adjuvated preparations reached antibody titers 20-fold and 15-fold higher than controls, respectively. Antibody titers remained high throughout a 120 day-period. Animals injected with the IMT504-adjuvated vaccine showed expansion of the anti-hemagglutinin antibody repertoire and a significant increase in the antibody titer with hemagglutination inhibition capacity when confronted to viral strains included or not in the vaccine. This indicates that the addition of IMT504 in flu vaccines may contribute to the development of significant cross-protective immune response against shifted or drifted flu strains.
Introduction
Immunostimulatory ODNs that are active in human cells are grouped into 2 major classes: (a) CpG ODNs, characterized by the presence of at least one active site bearing an unmethylated CpG in a given context (Krieg, 2002), and (b) PyNTTTTGT ODNs, which have at least one active site bearing the sequence PyNTTTTGT, in which Py is C or T and N is A, T, C, or G (Elías et al., 2003). Among other effects, both kinds of ODNs induce in vitro expression of co-stimulatory molecules acting on plasmacytoid dendritic cells. However, they show some differences: CpG ODNs induce the secretion of IFN alpha (Krieg, 2002), whereas PyNTTTTGT ODNs do not (Elías et al., 2003); PyNTTTTGT ODNs induce the release of GM-CSF acting on human NK and NKT cells in the presence of IL2, whereas CpG ODNs do not (our unpublished data); PyNTTTTGT ODNs stimulate the expansion of mesenchymal stem cell precursors both in vitro and in vivo, whereas CpG ODNs do not (Hernando-Insua et al., 2007). In vivo, both CpG and PyNTTTTGT ODNs act as potent adjuvants in vaccines by promoting cellular and humoral immune responses (Gupta and Cooper, 2008; Elias et al., 2005; Zhao et al., 2009). We have now found that IMT504, the prototype of PyNTTTTGT ODNs, is a potent adjuvant of the flu vaccine in young and elderly rats, which highly increases the diversity of the antibody (Ab) response against the flu virus strains both included and not included in the vaccine.
Materials and Methods
Animals
Female Sprague Dawley 8–12-week-old rats (considered young adults) or 20–22-month-old rats (considered elder animals) were obtained from FUCAL Laboratories and housed in the animal facility at Immunotech S.A. Animal care and use were according to NIH guidelines.
Oligodeoxynucleotides
Desalted phosphorothioate (PS) ODNs were purchased from Oligos ETC. ODNs were suspended in depyrogenated water, assayed for lipopolysaccharide (LPS) contamination using the Limulus test, and kept at −20°C until used. Purity was assessed by high performance liquid chromatography and polyacrylamide gel electrophoresis assays. ODN preparations were used if purity was >97% and LPS were undetectable. The ODN IMT504 sequence is as follows: 5’-TCATCATTTTGT CATTTTGTCATT-3’ (Immunotech S.A.). Also, a PS ODN with no action on the immune system (IMT022) was tested as a control: 5’-TGCTGCAAAAGAGCAAAAGAGCAA-3’.
Vaccines
The study was performed with a virosomal influenza vaccine (Inflexal V; Berna Biotech Ltd.), 2002 season for the Southern Hemisphere, containing 15 μg of HA of each of the following flu virus strains: A/New Caledonia/2099 H1N1, A/Moscow/10/99 H3N2, and B/Sichuan/379/99. A comparative study was performed using an equivalent antigen dose of a whole virus inactivated vaccine Fluarix (Glaxo SmithKlain-GSK, United Kingdom, 2004 season for the Southern Hemisphere) containing the following flu strains: A/New Caledonia/20/99 (H1N1), A/California/7/2004 (H3N2), and B/Malaysia/2506/2004. Commercial vaccines were diluted with saline to obtain the desired concentration and combined with a fixed amount of 50 μg of IMT504 in a total volume of 150 μL.
Immunization and sample collection
Animals received a single i.m. vaccine injection in the tibialis anterior muscles. Control groups received the vaccine alone, whereas the treated groups received the vaccine plus IMT504. At the indicated time points, rats were bled for antibody (Ab) assays in serum.
Antibody assays
Specific antibody titers were determined by end-point dilution ELISA. Briefly, ELISA plates (Maxisorb; Nunc) were coated overnight with 2 μg/mL of the commercial vaccine (Inflexal or Fluarix). Residual protein-binding sites were blocked with carbonate-bicarbonate buffer containing 8% nonfat milk. Samples were serially diluted with 0.05% Tween-20 in phosphate-buffered saline, and plates were incubated for 2 h at 37°C. Total IgG was evaluated using Sigma A5795 Ab conjugated with horseradish peroxidase. O-phenylendiamine dichloride solution (1 mg/mL) was finally added to the wells. End-point titers were defined as the highest serum dilution that resulted in an absorbance value 3-fold greater than that of nonimmune serum with a cut-off value of 0.15 O.D. at 490 nm.
Antibody binding assay on cellulose membranes
Several 15-amino acid (aa)-long peptides with a 3-aa displacement covering the total sequence of HA, corresponding to the flu strains A/New Caledonia/20/1999 (H1N1), A/Fujian/411/2002 (H3N2), and A/Viet Nam/1203/2004 (H5N1), were synthesized as an array on cellulose membranes by SPOT-synthesis as previously described (Frank, 1992; Beutling et al., 2008). Spots containing random peptide sequence (negative controls) were also synthesized on the probes. Five independent pools, each containing sera from 10 animals immunized with or without IMT504, were diluted 1:100 and assayed for peptide-binding capacity by incubation with the arrays for 4 h at room temperature. Then, membranes were incubated with AP-conjugated secondary Ab goat anti-rat IgG (START71; Serotec) and revealed by incubation with 5-bromo-4-chloro-3-indolphosphate p-toluidine dissolved in DMF combined with nitro blue tetrazolium. After color development, membrane probes were digitally recorded.
Signals were scanned and quantified using Image-Pro Plus software. Dye units were defined as being directly proportional to the amount of dye precipitated on the spots as determined through calibration of the scanning device. The reaction for a given spot was calculated as the intensity of the stain in the spot minus the membrane background surrounding each spot. Each membrane probe was separately analyzed. For statistical analysis, dye units of a given spot from each probe were grouped together, and Media and Standard Deviation were calculated. Control spots containing irrelevant sequences were considered negative. Differences with regard to negative spots were evaluated by Student's t-test and considered significant (“Positive”) when P<0.05.
Hemagglutination inhibition assay
The assay was performed as recommended by WHO [World Health Organization, 2002). Briefly, sera from 10 independent animals injected with the vaccine alone or in combination with the adjuvant were mixed with the receptor-destroying enzyme to remove nonspecific HA and inhibitors. Samples were incubated overnight at 37°C, inactivated at 56°C for 30 min, and adsorbed with packed red blood cells at 4°C. Treated sera were serially diluted 2-fold, and 8 HA units (an HA unit is defined as the highest virus dilution that yields complete hemagglutination) of the test virus were added. The reaction mix was then incubated at room temperature for 30 min. After this, a 0.5% suspension of turkey red blood cells was added, and hemagglutination patterns were examined. The lowest dilution of serum that inhibited hemagglutination was considered the hemagglutination inhibition (HI) titer.
Statistical analysis
Statistical significances of differences were evaluated by t-test on log-transformed data. If not stated, differences were considered significant when P<0.001.
Results
IMT504 greatly increases the antibody response to the flu vaccine in young adult animals
To study the effect of IMT504 on the efficiency of the flu vaccine, rats were inoculated with 15 μg of HA of the virosomal influenza vaccine (Inflexal V; Berna), with or without the addition of 50 μg of IMT504. This dose of adjuvant was chosen according to our previous studies using the recombinant Hepatitis B vaccine (Elias et al., 2005).
Thirty days postimmunization (d.p.i.), animals were bled; and Ab was measured by ELISA. Figure 1A shows that addition of IMT504 resulted in an about 20-fold increase in the Ab titer relative to that obtained without the addition of IMT504 (M±SEM with IMT504: 172,216±102,400; without IMT504: 8,109±8,635).

Antibody titers in young adult rats immunized with the influenza vaccine (Inflexal V; Berna Biotech). Animals were i.m. injected with the experimental vaccine containing 15 μg of HA with 50 μg of IMT504 (■), IMT022 () or saline (□), and saline plus IMT504 ( ). Bars represent the geometric mean ± the standard error of the mean (M ± SEM) for specific IgG titers as determined by end-point-dilution ELISA
). Bars represent the geometric mean ± the standard error of the mean (M ± SEM) for specific IgG titers as determined by end-point-dilution ELISA
To study the relevance of the specific nucleotide sequence in the generation of the response, a PS ODN (IMT022) with no activity in the immune system was tested. Addition of IMT022 to the vaccine did not show statistical difference when compared with the vaccine alone (Fig. 1A), and the animals immunized with IMT504 in the absence of the vaccine (Fig. 1A) behaved as the preimmune controls (not shown).
Anti-HA antibody titers remain high for at least 120 days in animals immunized with the IMT504-adjuvated vaccine
To investigate the extent of Ab response, rats inoculated with the vaccine containing 15 μg of antigen with or without the addition of IMT504 were periodically tested for Ab titers from 15 to 120 d.p.i. Figure 1B shows that the kinetics of both responses are similar. The antibody titer in the animals vaccinated with the addition of IMT504 was maximum at 30 d.p.i. and remained higher than that of the animals vaccinated without the addition of IMT504 throughout the 120-day period analyzed. Differences were statistically significant at all time points analyzed.
The flu vaccine adjuvated with IMT504 is also very effective in elderly animals
Elderly people, particularly those with cardiac or respiratory disease, are at highest risk of morbility and mortality associated with epidemics of influenza A or B, which occur almost every winter (Ilyinskii et al., 2008; Kumar and Burns, 2008). Therefore, we decided to study the effect of the flu vaccine adjuvated with IMT504 in elderly rats. A total of 40 rats aged 20–22 months old were injected i.m. with a vaccine containing 15 μg of HA either with or without the addition of IMT504 and the specific IgG titer measured at 30 d.p.i. Figure 2A shows that vaccination with the addition of IMT504 resulted in a 15-fold increase as compared with vaccination without addition of IMT504 (M±SEM with IMT504: 41,366±14,693, without IMT504: 2,541±6,123). It is worth noting that vaccination with the addition of IMT504 in elderly rats resulted in an antibody response about 5-fold higher (41,366 vs. 8,109) than that reached by an injection of the vaccine without addition of IMT504 in young adult animals (Fig. 1A). On the other hand, the specific IgG titers obtained with animals injected with the vaccine without IMT504 addition showed a widespread distribution from 200 to 25,600. However, in elderly animals vaccinated with the addition of IMT504, the Ab titer spread was clearly lower (from 25,600 to 102,400). Therefore, the addition of IMT504 to the vaccine not only enhances the antibody response but also homogenizes it. Finally, we observed that the kinetics of the responses with and without ODN were similar, but that the specific antibody titers in animals vaccinated with the addition of IMT504 remained high for at least 120 days after inoculation (Fig. 2B).
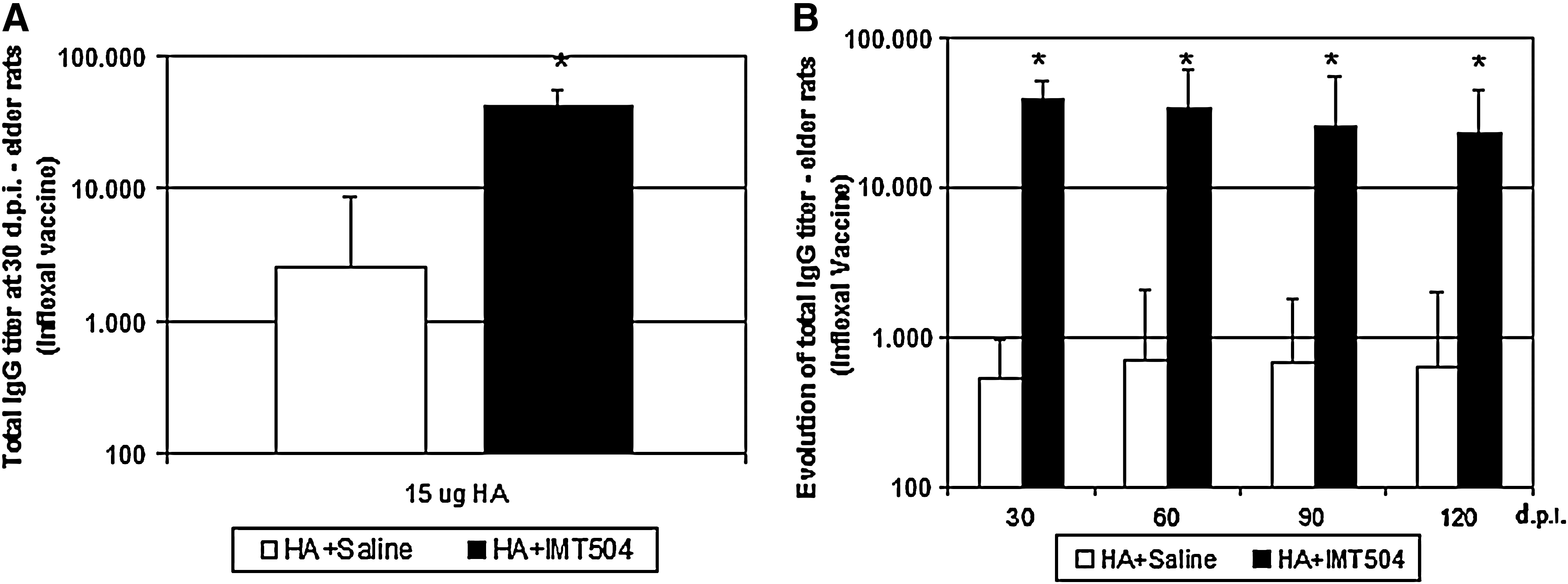
Antibody titers in elder rats immunized with the influenza vaccine (Inflexal V; Berna Biotech). Animals were i.m. injected with the experimental vaccine containing 15 μg of HA with either 50 μg of IMT504 (■) or saline (□). Bars represent the geometric mean±standard error of the mean (M ± SEM) for specific IgG titers as determined by end-point-dilution ELISA
IMT504 also increases the antibody response when combined with a whole virus inactivated flu vaccine
The results just described were obtained using a virosomal vaccine containing the HA and NA proteins of the influenza virus as antigens. A complementary study was performed using a whole virus inactivated vaccine (Fluarix, GSK) to assay the influence of the antigen molecular arrangement in the response generated by addition of IMT504 to the vaccine. To this end, the whole virus inactivated vaccine containing 15 μg of HA with or without addition of IMT504 was injected in rats. The response achieved with IMT504 was 15-fold higher than that with the nonadjuvated vaccine (M±SEM with IMT504: 58,813±28,672, without IMT504: 3,675±4,456) (Fig. 3). Therefore, the addition of IMT504 was effective in both the virosomal and the whole virus inactivated vaccines assayed in this study, which represent the main classes of current commercial flu vaccines.
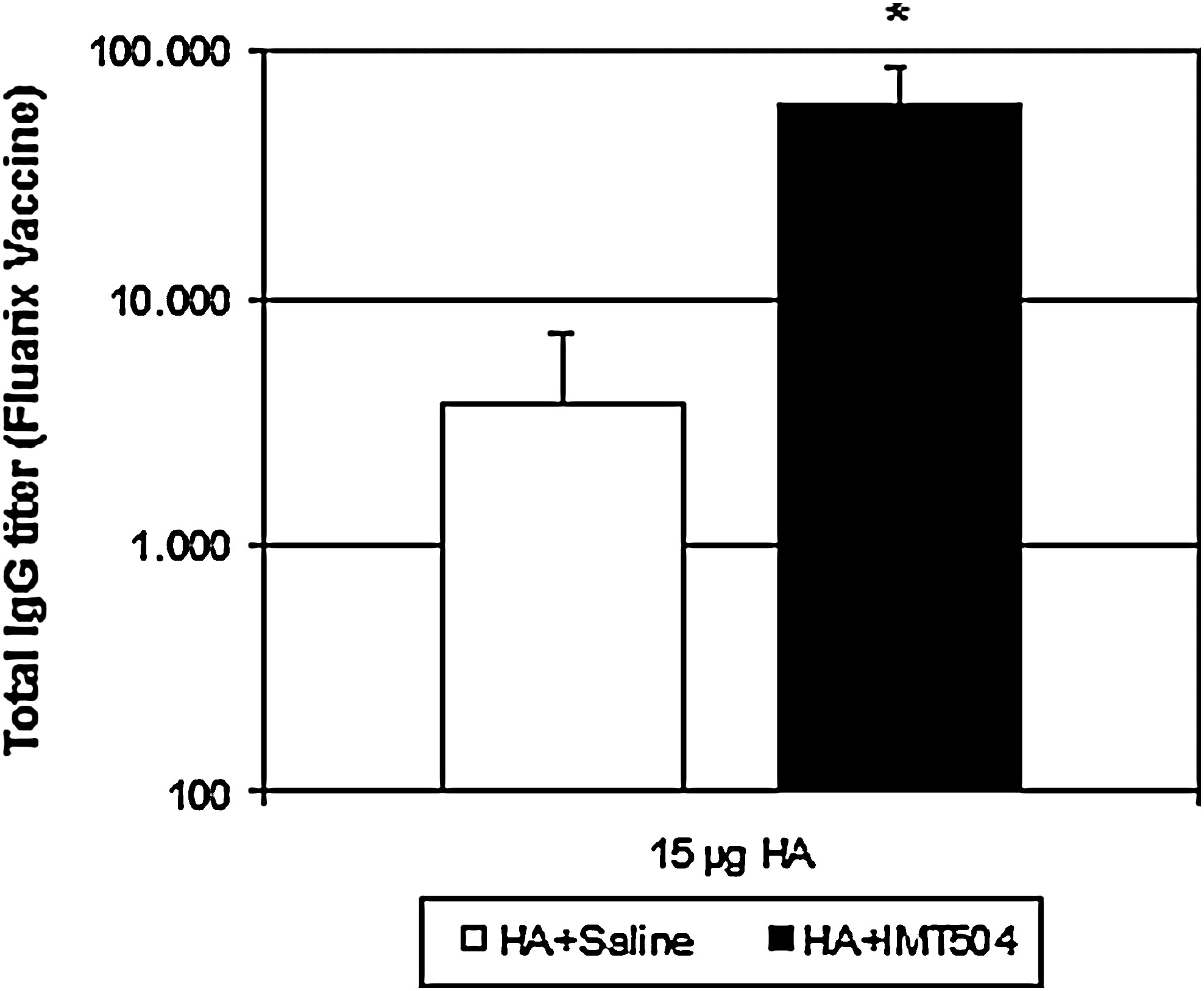
Antibody titers in elder rats immunized with the influenza vaccine (Fluarix; Glaxo SmithKline) plus ODN IMT504. Rats (10 per group) were i.m. injected with the experimental vaccine containing 15 μg of HA either with 50 μg of IMT504 (■) or without it (□). Each bar represents the geometric mean (M ± SEM) for anti-HA titers at 30 d.p.i. as determined by end-point-dilution ELISA. Asterisks indicate statistically significant difference compared to controls (P < 0.001).
Vaccination of animals with the flu vaccine containing IMT504 results in expansion of the repertoire of anti-HA antibodies
The diversity of antibodies produced during the immune response to the flu vaccine and to the flu vaccine adjuvated with IMT504 was studied using an array of peptides (15 aa long) dotted by synthesis on a membrane probe. This array spans the HA sequence of both flu virus strains included in the vaccine (A/New Caledonia/20/1999 (H1N1) or not included in the vaccine (A/Viet Nam/1203/200 and A/Fujian/411/2002). Five independent pools of sera from 10 animals each, vaccinated with the vaccine with saline or IMT504, were incubated with the membranes just described; and reactive dots were developed and analyzed. To our surprise, the intensity of the main spots present in the membranes incubated with the pool of sera from the animals injected with the nonadjuvated vaccine was not very different from those present in the membranes incubated with the pool of sera from the animals injected with the IMT504-adjuvated vaccine, although the titer in this last group was about 20-fold higher (Fig. 4). However, there were a significant large number of spots clearly detectable only in the membranes incubated with the pool of sera from the animals injected with the vaccine containing IMT504. This indicates that the higher Ab titer obtained after injection with the adjuvated vaccine was mainly attributable to the presence of antibodies reacting with epitopes poorly reactive during regular vaccination or, in other words, to a larger repertoire of anti-HA antibodies produced during vaccination in the presence of IMT504. A positive effect on antibody diversity has also been recently observed in a study of a flu vaccine supplemented with the oil-in-water MF59 adjuvant (Khurana et al., 2010).

Representative array on a cellulose membrane of 15-amino acid-long peptides spanning the HA sequence belonging to the A/Viet Nam/1203/2004, A/New Caledonia/20/1999, and A/Fujian/411/2002 flu virus strain incubated with five independent pools of sera of 10 animals each, injected with the vaccine containing 15 μg of HA plus saline or IMT504. White arrowheads (Δ) point to some of the main spots in the membrane incubated with the pool of sera from the animals injected with the nonadjuvated vaccine (nS) that show an intensity similar to the corresponding spots in the membrane incubated with the pool of sera from the animals injected with the IMT504-adjuvated vaccine (nO). The first peptide corresponding to the N-terminal region of the HA was dotted on the right edge of the membrane and the following peptides in the direction pointed by the black arrowhead (
Next, we studied the reactivity of antibodies present in sera from animals vaccinated with the 2 preparations just described to the array of peptides representing the HA1 or HA2 segment of the HA belonging to the flu virus strains present or not in the vaccine used in this study. HA1 contains the globular membrane-distal highly variable receptor binding domain of the HA and is the main target of neutralizing antibodies that are classically detected by the HI assay (Wilson and Cox, 1990). Figure 5 schematically shows that the antibodies present in the sera from the animals injected with the IMT504-adjuvated vaccine widely cover the HA1 region of the flu virus strain present (A/New Caledonia/20/1999) or absent (A/Fujian/411/2002 and A/Vietnam/1203/2004) in the vaccine and that the coverage of antibodies present in the sera from the animals vaccinated with the nonadjuvated vaccine is clearly less extended.

Schematic representation of the epitope coverage of the HA1 region of the HA belonging to three flu virus strains, one present (A/New Caledonia/20/1999) and two absent (A/Fujian/411/2002 and A/Vietnam/1203/2004), of one of the vaccines used in this study (Inflexal V; Berna Biotech, season 2002). Antibody presence in sera from animals injected with the nonadjuvated vaccine or the IMT504-adjuvated vaccine was detected by arranges of 15-amino acid-long peptides covering in this case only the HA1 region belonging to the above-mentioned strains as described in Fig. 4. Spots were scanned and analyzed statistically as described in the Materials and Methods section. Gray bar, vaccine; black bar, vaccine+IMT504.
On the other hand, the HA2 segment is the main component of the stem domain of HA that is conserved among Influenza A virus strains and is the main target of antibodies with a wide cross-protection range (Webster et al., 1992; Corti et al., 2010). Figure 6 shows that the antibodies present in the sera from the animals injected with the IMT504-adjuvated vaccine widely cover the HA2 region belonging to 2 flu virus strains, one present (A/New Caledonia/20/1999) and one absent (A/Vietnam/1203/2004) in the vaccine, and that the coverage of the antibodies present in the sera from the animals vaccinated with the nonadjuvated vaccine was markedly poor.

Schematic representation of the epitope coverage of the HA2 region of the HA belonging to two flu virus strains, one present (A/New Caledonia/20/1999) and one absent (A/Vietnam/1203/2004), of one of the vaccines used in this study (Inflexal V; Berna Biotech, season 2002). Antibody presence in sera from animals injected with the nonadjuvated vaccine or the IMT504-adjuvated vaccine was detected by arrays of 15-amino acid-long peptides as described in Fig. 4, covering in this case the HA2 region belonging to the above-mentioned flu virus strains. Spots were scanned and analyzed. Gray bar, vaccine; black bar, vaccine +IMT504.
Sera from animals immunized with the IMT504-adjuvated vaccine have, in general, better performance in HI assays than sera from animals immunized with the nonadjuvated vaccine
Table 1 shows that immunization of young adult animals with the IMT504-adjuvated virosomal (Berna Biotech Ltd.) vaccine resulted in a 3- or 4-fold increase in the titer of antibodies with HI capacity regarding the virus strain assayed. However, in elderly animals, titers were the same for the B and A (H1N1) strains assayed. Both of these strains were included in the vaccine. In contrast, there was a 4-fold increase in the titer for the A (H3N2) strain included in the vaccine (Moscow 10/99) in sera from elderly animals immunized with the IMT504-adjuvated vaccine and for an A (H3N2) strain not included in the vaccine (Fujian 411/02). This flu virus strain caused a severe 2003–2004 flu season due to a reassortment event which provided a HA gene that codifies for a protein nonrecognizable by the antibodies raised by the seasonal vaccine used in that period, which included the A (H3N2) strain Moscow 10/99 present in the experimental vaccine here assayed (Ghedin et al., 2005).
HI assay of flu virus strains against 2-fold serial dilutions of sera from rats immunized with the IMT504-adjuvated vaccine or sera from rats immunized with the nonadjuvated vaccine. Each test was performed in triplicate with sera from 10 different animals per group immunized with 15 μg of Inflexal vaccine (Berna Biotech) alone or in combination with 50 μg of ODN IMT504. The HA titer is expressed as the reciprocal of the highest dilution showing complete HA. Reference sera from ferrets immunized at CDC with the specific strain showed the following HI titer: Sichuan, 370±47; Caledonia, 338±36; Moscow, 736±96; Fujian, 1152±85. Figures represent M±SEM.
Differences were considered significant when P<0.001.
ODN, oligonucleotide; HA, hemagglutinin; HI, hemagglutination inhibition; M±SEM, mean±standard error of the mean.
The HI assay was also performed with sera from young animals vaccinated with the inactivated GSK vaccine (Table 2). Once again, there was a 3-fold increase in the titer for the A (H1N1) strain included in the vaccine (New Caledonia) in the sera from the animals immunized with the IMT504-adjuvated vaccine. We also observed a 4-fold increase for the A (H3N2) strains not included in the vaccine (Moscow and Fujian) when compared with the nonadjuvated vaccine.
HI assay of flu virus strains against 2-fold serial dilutions of sera from rats immunized with the IMT504-adjuvated vaccine or sera from rats immunized with the nonadjuvated vaccine. Each test was performed in triplicate with sera from 10 different animals per group immunized with 15 μg of Fluarix vaccine (GSK) alone or in combination with 50 μg of ODN IMT504. The HA titer is expressed as the reciprocal of the highest dilution showing complete HA. Reference sera from ferrets immunized at CDC with the specific strain showed the following HI titer: Caledonia, 352±32; Moscow, 1152±85; Fujian, 1344±149. Figures represent M±SEM.
Differences were considered significant when P<0.001.
Discussion
Ideal flu vaccines should elicit an effective immune response against the virus strains included in the vaccine as well as against antigenic drifted strains that can reduce the vaccine performance and cause epidemic outbreaks (Webster et al., 1992). Ideal vaccines should also be effective in elderly subjects whose protection rates could be very low (Kumar and Burns, 2008). Both these issues were addressed in the present study. First, we established that IMT504, the prototype of a major class of immunostimulatory ODNs (Elías et al., 2003; Rodriguez et al., 2006), is a strong adjuvant if combined with seasonal commercial flu vaccines representing virosomal and whole virus inactivated technologies. Specific antibody titers in sera from young adult animals injected with the IMT504-adjuvated vaccine were about 20-fold higher than in sera from animals injected with the nonadjuvated vaccine, and titers remained stable for at least 120 days postvaccination. On the other hand, the IMT504-adjuvated vaccine was very effective in elderly animals, as the specific antibody titers in sera were about 15-fold higher than those in sera from elderly animals injected with the nonadjuvated vaccine and about 5-times higher than those in sera from young adult animals injected with the nonadjuvated vaccine. Also, similar to that observed in young adult animals, the specific antibody titers in elderly animals remained stable for at least 120 days postvaccination. Further, it is important to point out that the antibody immune response in elderly animals was clearly less widespread in the group injected with the IMT504-adjuvated vaccine, an important fact concerning the prediction of vaccination outcome at the individual level. Also, a comparison of the quality of the antibody immune response between the regular vaccine and the vaccine containing IMT504 as adjuvant was performed. For this, arrays of 15-aa peptides dotted by synthesis on a membrane, spanning the HA sequence belonging to several flu virus strains, were incubated with sera from animals vaccinated with different vaccine preparations; and reactive spots were developed and analyzed. Using this technique, we established that the sera from the animals injected with the regular vaccine or adjuvated with IMT504 possessed a variety of antibodies specific for different portions of the HA sequence corresponding to the viral strains present or not in the vaccine. Moreover, we observed that the number of positive spots in the membranes incubated with the pool of sera from the animals injected with the IMT504-adjuvated vaccine was significantly larger than the positive spots observed in the membranes incubated with the pool of sera immunized with the nonadjuvated vaccine. However, we also observed that the intensity of the spots generated by incubation with sera from animals injected with the nonadjuvated vaccine was, in general, quite similar to that of the spots generated by incubation with sera from animals injected with the IMT504-adjuvated vaccine (Fig. 4). This indicates that a large portion of the antibody excess produced by an injection of the adjuvated vaccine could be attributable to anti-HA antibodies with specificity for regions of the HA sequence which behave as poor antigenic regions during the immune response to the nonadjuvated vaccine. In other words, in the presence of IMT504, the vaccine is able to elicit greater variability in the anti-HA antibodies. A similar effect has also been recently observed for a flu vaccine supplemented with the oil-in-water adjuvant MF59 (Khurana et al., 2010).
The HA viral protein is post-translationally cleaved by host enzymes that generate 2 polypeptides (HA1 and HA2) which remain linked by a disulfide bond. HA1 (∼300 aa long) forms a membrane-distal globular domain that contains the receptor-binding site, and most determinants are recognized by virus-neutralizing antibodies which are classically detected by the HI assay. The larger antibody coverage of the HA1 segment by the antibodies present in the sera from the animals inoculated with the IMT504-adjuvated vaccine observed during the present study (Fig. 5) was accompanied by a significant increase in the titer of antibodies with capacity for HI when confronted with several flu virus strains (Table 1). As previously shown, IMT504 increases total Ab titers by 20-fold, whereas the latter indicates an 8-fold increase. This apparent discrepancy may be explained by the increase in antibody variability. In this context, many of the new antibodies generated in the presence of IMT504 did not react with those epitopes that are relevant to the inhibition of hemagglutination. Particularly interesting was the result obtained with the sera from elderly rats inoculated with the nonadjuvated vaccine, which was negative in the HI assay when confronted with a H3N2 strain (Fujian A/F/411/02) but clearly reactive if the animals were previously inoculated with the IMT504-adjuvated vaccine. This flu virus strain caused a severe 2003–2004 flu season due to a reassortment event that provided a HA gene which codifies for a protein nonrecognizable by the neutralizing anti-HA antibodies raised by the seasonal vaccine used in that period (Ghedin et al., 2005). Therefore, our results suggest that an IMT504-adjuvated flu virus vaccine may be useful to protect against, at least, some emerging drifted strains.
HA2 (180 aa long) contains the most conserved region, mainly near the HA cleavage site, among all influenza A subtypes (Gerhard et al., 2006). The protective potential of antibodies with specificity for some HA2 epitopes has been demonstrated in several studies (Gocnik et al., 2007; Prabhu et al., 2009; Corti et al., 2010). Thus, HA2, or portions of it, has been postulated as a possible antigen candidate for a prospective universal flu vaccine (Gerhard et al., 2006; Steel et al., 2010). Therefore, of special significance is the observation that the antibodies present in the sera from animals inoculated with the IMT504-adjuvated vaccine are able to react with many epitopes present in HA2 and that the antibodies in the sera from animals inoculated with the nonadjuvated vaccine are not (Fig. 6). Once more, this suggests that current seasonal flu vaccines in combination with IMT504 may considerably increase their field of protection, covering new emerging drifted or shifted viral strains. Therefore, antibody diversity should be investigated when performing clinical trials using a flu-vaccine adjuvated with IMT504 to evaluate possible cross-protection.
In conclusion, addition of IMT504 to current seasonal flu vaccines results in higher HA specific antibody titers in young adult and elderly rats and expansion of the anti-HA (anti-HA1 and anti-HA2) antibody repertoire.
Footnotes
Acknowledgment
We thank S. Daenicke for help in the peptide spot synthesis.
Author Disclosure Statement
None of the authors have competing financial interests.