
Abstract
Oligonucleotides containing CpG motifs (cytosine-phosphate-guanosine oligodeoxynucleotide [CpG ODN]) display strong immunostimulatory effects, and polycations have been previously reported as cellular delivery system. In the present study, we investigated the adjuvant properties of combinations of a CpG ODN with various polycations (poly-arginine, poly-lysine, poly-histidine, or chitosan) in an ovalbumin vaccination model. We showed that, when combined to CpG ODN, poly-arginine and poly-histidine, but not poly-lysine or chitosan, enhanced efficiently both the IgG antibody production and the number of splenocytes secreting interferon-gamma after stimulation with a CD8+ T cell-restricted peptide. Interestingly, CpG ODN-poly-arginine, which was the most efficient, compared favorably to the complete Freund's adjuvant and aluminium salts and induced no local toxicity, making this combination a very attractive adjuvant for vaccines.
Introduction
V
Cytosine-phosphate-guanosine oligodeoxynucleotides (CpG ODN) are short single-stranded synthetic DNA molecules that contain unmethylated CpG dinucleotides similar to those commonly found in bacterial DNA. They are taken up by cells via adsorptive endocytosis and bind to the toll-like receptor 9 (TLR9) within the endosomes. In humans, TLR9 is selectively expressed by B cells and plasmacytoid dendritic cells (pDCs), whereas the expression in mice also includes myeloid cells (Barchet et al., 2008). The binding of CpG ODN to its receptor triggers an immunostimulatory cascade inducing the activation and/or proliferation of multiple immune cells, including B and T lymphocytes, natural killer cells, and pDCs that produce IL-6, IL-12, IFN-gamma, IFN-alpha, or TNF-alpha (Klinman et al., 1996; Yi et al., 1996; Hartmann et al., 1999; Bauer et al., 2001a; Kadowaki et al., 2001). Several in vivo experiments have shown that adding CpG ODN to conventional protein antigens boosts antibody production, induces a Th1 shift in the antibody isotype, and increases the number of spleen cells secreting IFN-gamma (Chu et al., 1997; Davis et al., 1998; Tighe et al., 2000; Zhang et al., 2007).
Interestingly, enhancement of a specific immune response against an MHC-class I epitope can be achieved when poly-arginine is combined with a CpG ODN (Lingnau et al., 2002). Poly-arginine noncovalently interacts with negatively charged proteins or nucleic acids and has been reported to enhance antigen uptake into APCs in vitro (Buschle et al., 1997). Actually, several other polycationic compounds (such as poly-lysine, poly-histidine, chitosan, or others) have been found to enhance antigen uptake into cells in vitro and successful immunizations have been described with some of these associations (Buschle et al., 1997; Schmidt et al., 1997; Borges et al., 2008; Takemoto et al., 2009). However, it remains to be determined which type of polycations can most efficiently enhance immune responses, as no study has formally compared poly-arginine to other polycations. This point is of great interest, given the potent clinical development of these fully synthetic adjuvants.
Therefore, the objective of this study was to identify the best association of CpG ODN and polycation in a full protein antigen vaccine model. We tested the ability of different polycations, with or without CpG ODN, to elicit antibodies and an MHC-class I-restricted cellular response. We then compared the optimal formulation to the commonly used adjuvants: Alum, CFA, and CpG ODN emulsified in incomplete Freund's adjuvant (IFA).
Materials and Methods
Cells
Splenocytes were cultured at 37°C in a 5% CO2 humidified incubator in a complete medium consisting of Dulbecco's modified Eagle's medium (DMEM; Lonza, Verviers, Belgium) supplemented with 10% heat inactivated horse serum (Invitrogen, Cergy Pontoise, France), 2 mM L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin (Lonza).
Reagents
EndoGrade OVA purified from chicken egg white was purchased from Hyglos (Regensburg, Germany). Immunodominant H-2b OVA epitope (OVA257–264, amino acid sequence SIINFEKL) and irrelevant H-2b-restricted peptide (amino acid sequence SIYRYYGL) were synthesized by Proteogenix (Oberhausbergen, France).
Poly-arginine hydrochloride (polyR, mol wt 5000–15,000), poly-lysine hydrochloride (polyK, mol wt>30,000), poly-histidine hydrochloride (polyH, mol wt≥5000), and chitosan (low molecular weight, deacetylation≥75.0%) were obtained from Sigma-Aldrich (Saint Quentin Fallavier, France).
The phosphorothioate CpG ODN used in this study (CpG-28, 5′-TAAACGTTATAACGTTATGACGTCAT) and the control ODN (5′-TAAA
CFA and IFA were obtained from Sigma-Aldrich and Alum (Aluminium Hydroxyde Gel, Alhydrogel 2%) from Superfos Biosector (Vedbaek, Denmark).
Horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG, IgG1, and IgG2b antibodies were purchased from Alpha Diagnostic International (Cortecnet, Voisins-Le-Bretonneux, France).
Microparticle preparation
Solutions of ovalbumin (1 mg/mL), polyR, polyK, and polyH (10 mg/mL) were prepared in distilled water, aliquoted, and stored at −20°C. Solutions (10 mg/mL) of chitosan were prepared extemporaneously as previously described (Han et al., 2008). Stock solutions (10 mg/mL) of CpG ODN dissolved in NaCl 0.9% were aliquoted and stored at −20°C.
Microparticles were prepared by mixing 10 μg of CpG ODN and 20 μg of OVA. Polycations were dissolved in distilled water and slowly added under mild agitation in a final volume of 100 μL.
Electrophoretic mobility shift assay
The formation of CpG ODN complexes was observed by electrophoretic mobility shift assay. Ten micrograms CpG ODN was combined with increasing doses (1–70 μg) of polyR, polyK, polyH, or chitosan in a final volume of 100 μL, and incubated for 30 minutes at room temperature. Aliquots of samples (10 μL) were loaded in 10% glycerol (Sigma-Aldrich) with 0.01% bromophenol blue (Sigma-Aldrich), and run for 30–60 minutes at 80 V on 1% agarose containing 0.5 μg/mL ethidium bromide (Bio-Rad, Marnes-la-Coquette, France) in 1×TBE (45 mM Tris-borate, 1 mM EDTA, pH 8.0; Bio-Rad) running buffer.
Particle size analysis
CpG ODN (10 μg), OVA (20 μg), and polyR (30 μg) were mixed in a final volume of 100 μL. The solutions were diluted 5 times in distilled H2O before analysis by the granulometer VASCO (Cordouan Technologies, Pessac, France). Measurements are based on dynamic light scattering of backscattered light.
Stability assessment of the CpG ODN-OVA-polyR particles
Particles containing CpG ODN (10 μg), OVA (20 μg), and polyR (70 μg) were dispersed in phosphate-buffered saline (PBS) in several tubes. The samples were stored at 37°C for 3 weeks. At predetermined intervals, one tube was centrifuged at 16,000 g (Eppendorf Centrifuge 5415 R). In the supernatants, the amount of CpG ODN released from the particles was quantified spectrophotometrically at 260 nm. The amount of CpG ODN contained in the pellet was measured as previously described, but after trypsin digestion (Invitrogen).
Determination of the OVA content in the microparticles
Formulations containing CpG ODN (10 μg), OVA (20 μg), and various polycations (70 μg) were prepared in distilled water in a final volume of 100 μL. The suspensions containing the microparticles were centrifugated at 16,000 g (Eppendorf Centrifuge 5415 R) at 20°C. The resulting pellets, which contained the microparticles, were resuspended in 10% sodium dodecyl sulfate (SDS) and incubated for 1 hour at 60°C under mild agitation. The ovalbumin content of the microparticles was observed by standard polyacrylamide gel electrophoresis (PAGE) under denaturing condition (SDS-PAGE) followed by Coomassie staining.
Mice immunization
Five- to 6-week-old female BALB/cByJ (H-2d) or C57BL/6J (H-2b) mice (Charles River Laboratories, L'Abresle, France) were immunized or immunized and boosted subcutaneously at the left flank at a 14 days' interval with 20 μg OVA alone or formulated with 10 μg CpG ODN and/or 70 μg polycation (100 μL injected per mouse, used 30 to 45 minutes after preparation), or with 20 μg OVA combined with CFA (v/v) or Alum (500 μg), or with the mix CpG ODN-OVA (10–20 μg) emulsified in IFA (v/v). The mice were euthanized 14 days after the last immunization. Spleens and blood were collected. Cell-mediated immune responses were analyzed by ELISPOT assay and antigen-specific antibody titres were measured by ELISA.
Animal use and handling were performed according to the French laws for animal experimentation.
Determination of anti-OVA antibody levels in serum by ELISA
Ninety-six-well ELISA plates (ATGC, Marne La Vallée, France) were coated with 50 μL/well of OVA antigen (50 μg/mL in PBS, pH 7.4), and incubated at 4°C overnight. Plates were washed with PBS containing 0.05% Tween-20 (PBS-T) and blocked by incubation with 100 μL/well of 1% bovine serum albumin (BSA) in PBS for 1 hour at 37°C. Plates were washed with PBS-T. Serum samples (100 μL), diluted 1:100 (for detection of IgG and IgG1) or 1:50 (for detection of IgG2b), were incubated for 1 hour at room temperature. Plates were washed with PBS-T. Diluted HRP-conjugated goat anti-mouse IgG (1:1000), IgG1 (1:10,000), or IgG2b (1:5000) was added to detect the bound antibodies. As described in Schellack et al. (2006), the Th1/Th2 balance was assessed by the IgG2b/IgG1 ratio, since the IgG2a encoding gene is deleted in C57BL/6 mice. One hour after incubation at room temperature, plates were washed with PBS-T. The reaction was developed with TMB substrate-developing solution (BD Bioscience, Le Pont de Claix, France) and stopped with 2N H2SO4. The absorbance was measured at 450 nm. The amount of mouse IgG, IgG1, or IgG2b in samples was calculated relative to mouse anti-ovalbumin reference calibrators composed of dilutions of ascites from mice immunized with ovalbumin. Values were assigned as arbitrary anti-ovalbumin activity units (U/mL).
Evaluation of the relative numbers of IFN-gamma secreting cells by ELISPOT
Splenocytes were prepared according to standard procedures. Briefly, each spleen was transferred onto a 70 μm cell strainer. The cells were then gently forced through the meshes with a syringe pestle and collected in cold PBS containing 1% BSA. Pelleted cells were resuspended into 2 mL of erythrocyte lysis buffer (0.15M NH4Cl, 10 mM KHCO3, and 0.1 mM Na2EDTA, pH 7.2) and incubated for 2 minutes on ice. The reaction was stopped by addition of 12 mL of cold PBS containing 1% BSA. Pelleted cells were resuspended in 4 mL of complete medium consisting of DMEM (Lonza) supplemented with 10% heat-inactivated horse serum (Invitrogen), 2 mM L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin (Lonza).
Anti-mouse IFN-gamma mAb-precoated 96 PVDF-bottomed-well plates (Diaclone, Besançon, France) were washed with PBS. Splenocytes were added to triplicate wells at a concentration of 5×105 cells/well, and cultured at 37°C in the presence of 10 μg/mL MHC class I-restricted peptides (immunodominant H-2b OVA epitope or irrelevant H-2b-restricted peptide), or 100 ng/mL phorbol myristate acetate (PMA) and 10 μg/mL ionomycin (both from Sigma-Aldrich), or complete medium alone for 18–20 hours. The plates were then washed with PBS containing 0.1% Tween-20 and incubated with biotinylated anti-mouse IFN-gamma antibody diluted 1:100. Bound antibodies were detected using streptavidin-alkaline phosphatase diluted 1:5000, and observed using 5-bromo-4-chloro-3-indolylphosphate/nitroblue tetrazolium chloride substrate buffer. Positive controls included cells stimulated with PMA and ionomycin. Stained spots were counted on a Cellular Technology Ltd. system using the ImmunoSpot 5.0.3 software version (Bonn, Germany), and results were expressed as the numbers of IFN-gamma spot forming cells per 5×105 cells.
Statistical analysis
All data are presented as mean±standard deviation. Differences between groups were determined using Student's t-test. Results were considered significant at a P value of <0.05.
Results
Physical characteristics of CpG ODN and OVA when combined with different polycations
The ability of polyR, polyK, polyH, and chitosan to form complexes with CpG ODN was investigated by agarose gel electrophoresis, combining CpG ODN with increasing doses of polycation (Fig. 1a). As expected, CpG ODN alone migrated into the gel, whereas the addition of polycation peptides completely abolished the migration of 10 μg of CpG ODN for doses above 10 μg for polyR, 20 μg for polyK, and 30 μg for polyH. Chitosan bound very weakly to CpG ODN, as chitosan was unable to inhibit migration of CpG ODN, even with doses as high as 600 μg (data not shown). For subsequent experiments (ie, in vivo applications), a mass ratio 10:70 of CpG ODN:polycation was selected to be sure that the positive charges of polycation exceed the negative charges of both the CpG ODN and the OVA antigen.
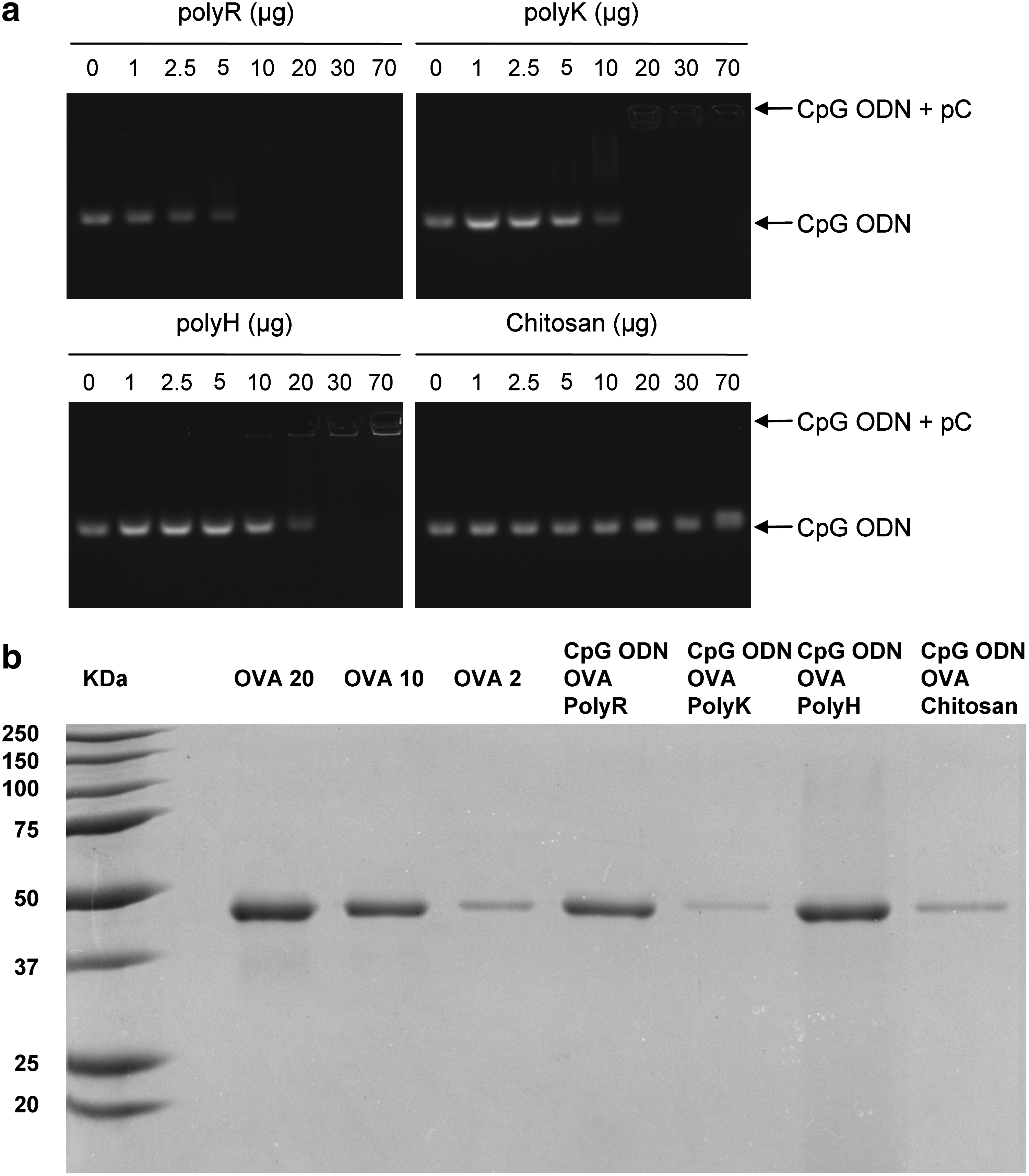
Physicochemical characteristics of the different CpG ODN-OVA-polycations formulations.
Polycations spontaneously built complexes with CpG ODN and OVA, which increased turbidity of the solutions. The different formulations of CpG ODN-OVA-polycation were centrifuged and the pellets were analyzed for their OVA content by SDS-PAGE. PolyR and polyH-precipitates entrapped significantly more OVA than their polyK and chitosan counterparts, which contained <10% of the total OVA antigen (Fig. 1b).
The formulation of polyR with CpG ODN and OVA was further characterized by a particle size analyzer. The diameter of particles slowly increased with time, starting from about 400 nm and reaching 1 μm after 10 hours (Fig. 2). For in vivo applications, the CpG ODN-OVA-polycation formulations were injected 30 to 45 minutes after preparation.

Size analysis of the CpG ODN-OVA-polyR particles CpG ODN was mixed with OVA before addition of polyR. Particles' size were analyzed for a 20-hour period after preparation (VASCO granulometer).
To evaluate the stability of these microparticles, CpG ODN-OVA-polyR particles were incubated at 37°C for 3 weeks in saline buffer. Over a 3-week incubation period, the dissociation of CpG ODN from particles was limited (≤4%), as assessed by UV spectrophotometry at 260 nm. All the remaining CpG ODN could be retrieved from these particles after digestion of their protein content with trypsin.
PolyR compares favorably to other polycations to enhance both humoral and cellular immune responses when combined with CpG ODN
In this set of experiments designed to compare the different polycations (polyR, polyK, polyH, and chitosan), we first observed that the addition of a polycation to the OVA antigen was able to induce anti-OVA IgG antibodies independently of the nature of the polycations, as early as 2 weeks after a single immunization (Fig. 3a). Immunization with 20 μg OVA alone elicited no significant antibody titers (data not shown).
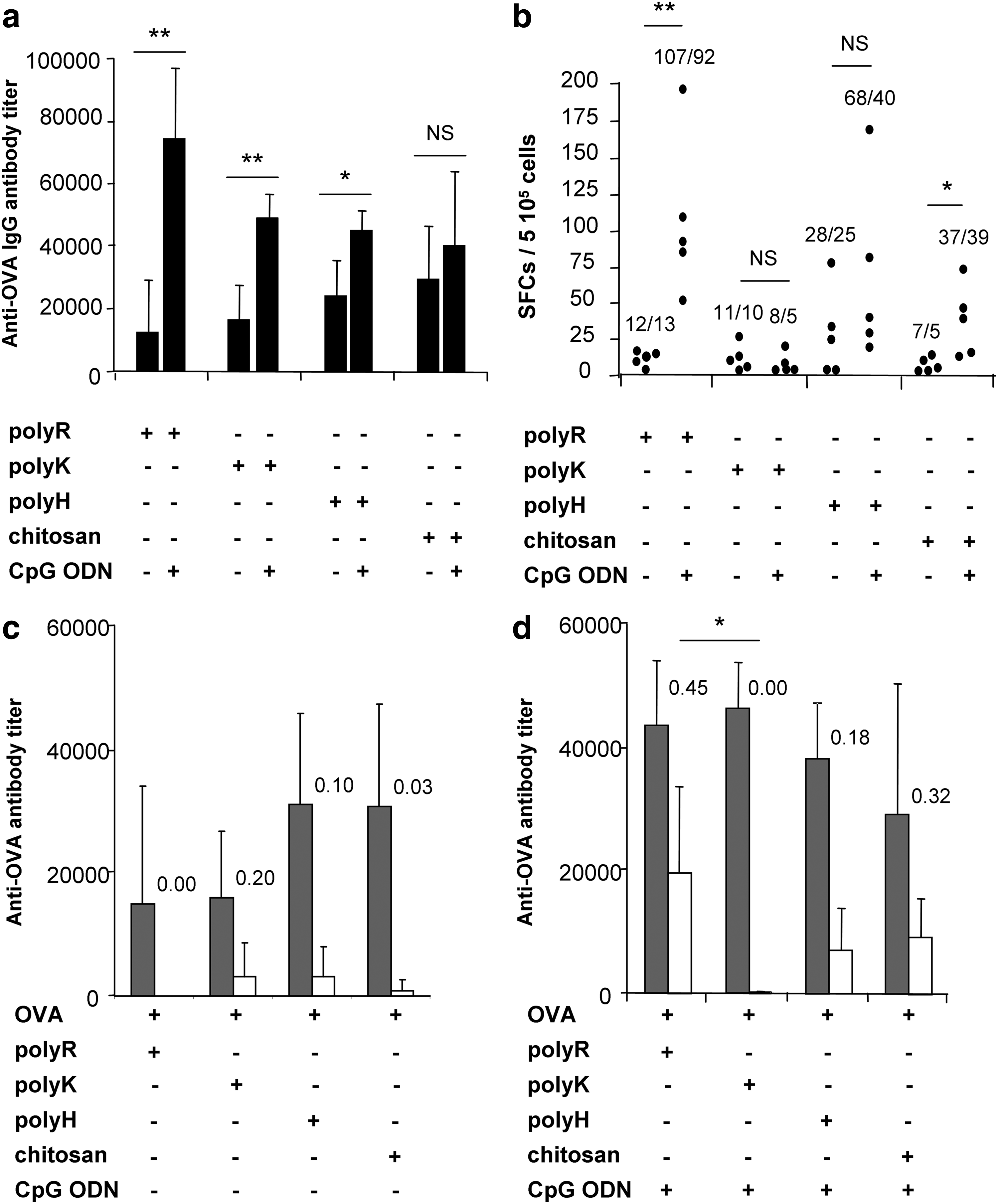
OVA-specific immune responses in mice immunized with OVA co-formulated with CpG ODN and various polycations C57BL/6J mice (n=5/group) were immunized against OVA (20 μg) with combinations of CpG ODN (10 μg) and various polycations (70 μg). OVA-specific antibodies titers, 14 days after one single immunization, were measured by ELISA:
Interestingly, the combination of CpG ODN with the various polycations further enhanced the anti-OVA IgG immune response, especially when polyR was used (CpG ODN-OVA-polyR vs. OVA-polyR: P<0.01) (Fig. 3a). This effect was not observed when an ODN devoid of any CpG motif was used, demonstrating that the enhancement of the immune responses induced by the CpG ODN was CpG-specific (CpG ODN-OVA-polyR vs. non-CpG ODN-OVA-polyR: 90,150±15,860 U/mL vs. 42,840±22,520 U/mL; n=6/group; P=0.002).
In spite of their abilities to trigger an IgG response, all formulations were not equivalent with respect to their ability to induce a Th1-biased immune response, which was clearly enhanced by CpG ODN. Indeed, polycations alone only poorly induced CD8+ T lymphocytes (Fig. 3b). When combined to CpG ODN, the different polycations tested induced an OVA-specific immune response with variable efficiencies. Hence, combinations of CpG ODN with polyR induced the highest numbers of IFN-gamma secreting-splenocytes (P<0.01 for CpG ODN-polyR when compared to the polyR alone), whereas chitosan and polyH were less efficient and polyK inefficient, either alone or in combination with CpG ODN.
When polycations were used alone, the IgG2/IgG1 ratio of the humoral immune response remained below 0.20 (Fig. 3c). The IgG2b/IgG1 ratio was variable among the different CpG ODN-polycation-based compositions, polyR being the most efficient formulation for inducing IgG2 antibodies (IgG2/IgG1 ratio: 0.45) and polyK being the least efficient (Fig. 3d).
The CpG ODN-polyR-based vaccine compares favorably to other well-known adjuvants
Next, we compared the efficiency of the CpG ODN-polyR adjuvant with well-known adjuvants such as CFA, Alum, and a combination of CpG ODN and IFA. As shown in Fig. 4, all these adjuvants significantly enhanced the production of IgG antibodies over CpG ODN alone (P<0.001 for all 3 adjuvants vs. CpG ODN alone). However, the CpG ODN co-formulated with polyR compared favorably to Alum and CpG ODN-IFA and the IgG2/IgG1 ratio was higher with this formulation (>0.47 for CpG ODN-OVA-polyR, 0.41 for CpG ODN-OVA-IFA, 0.28 for CFA-OVA, and 0.20 for Alum-OVA) (Fig. 4c, d).
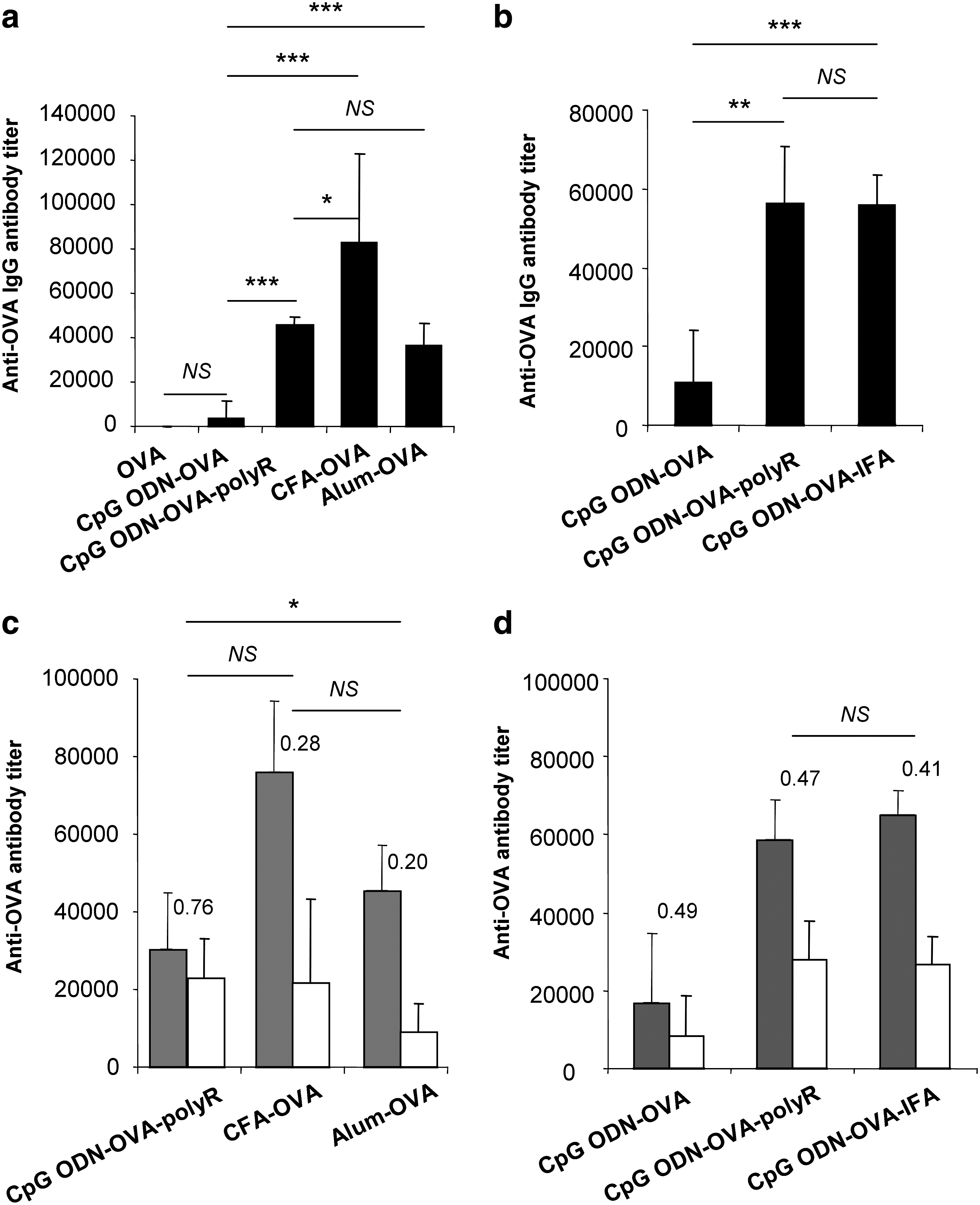
Comparison of different adjuvants on OVA-specific humoral immune responses
Most interestingly, the CpG ODN-polyR-based vaccine was significantly more efficient than Alum or CpG ODN-IFA, or even CFA for the induction of cell-mediated immune responses (Fig. 5).

Role of adjuvants on OVA-specific cellular immune responses C57BL/6J mice (n=6/group) were immunized against OVA (20 μg) combined with CpG ODN (10 μg), polyR (70 μg), Alum (500 μg), IFA (v/v), or CFA (v/v). Cell-mediated immune response, 14 days after one single immunization, was analyzed by ELISPOT assay. Splenocytes from immunized mice were re-stimulated in vitro with the OVA-derived peptide SIINFEKL and the numbers of IFN-gamma-SFCs were measured. Values for individual mice (dots) and mean/median value for each group are shown. Results are representative of 3 independent experiments (NS: not significant, *P<0.05, **P<0.01, ***P<0.001).
Finally, the injection sites were inspected 2 weeks after the primary immunization. As shown in Fig. 6, no noticeable inflammatory reaction was observed following the injection of OVA, CpG ODN-OVA, or CpG ODN-OVA-polyR. In contrast, the injection of OVA combined with CFA or CpG ODN-OVA emulsified in IFA (not shown) led to a major local inflammatory reaction.
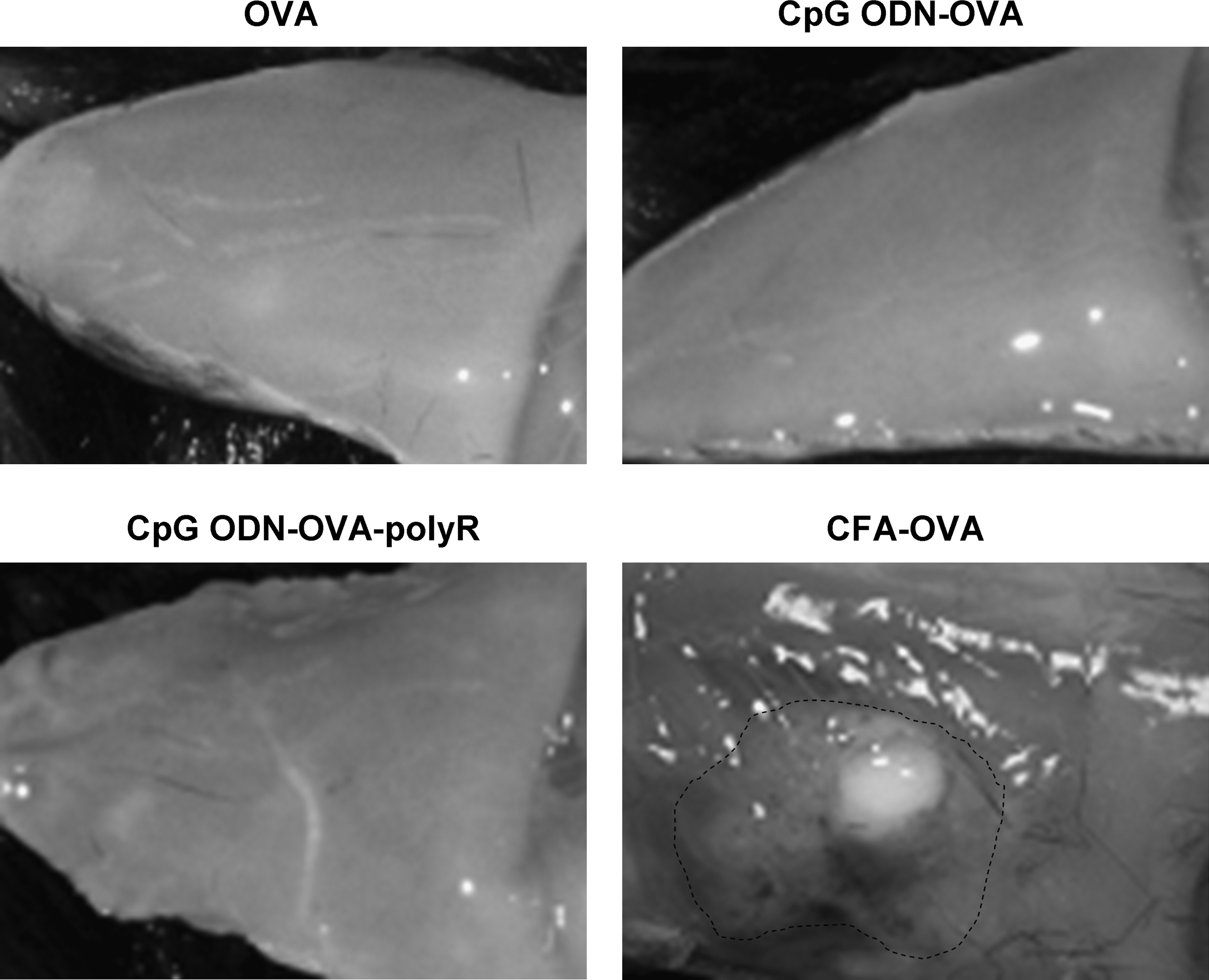
Macroscopic aspect of the injection sites in mice immunized with OVA co-formulated with CpG ODN, polyR, or CFA. At day 14 after vaccination with OVA (20 μg) combined with CpG ODN (10 μg), polyR (70 μg), or CFA (v/v), the injection sites were inspected for the presence of an inflammatory reaction. While injections with OVA, CpG ODN-OVA, and CpG ODN-OVA-polyR did not show noticeable changes, CFA induced a marked inflammatory reaction. Representative pictures from 1 out of 5 mice/group.
Discussion
In the present study, we investigated the adjuvant effects of various combinations of CpG ODN and polycations (polyR, polyK, polyH, and chitosan) on OVA-specific humoral and cellular immune responses. We showed that all polycations were not equally efficient when combined to CpG ODN. PolyR appeared to be the most powerful polycation for inducing both humoral and CD8+ T-cell immune responses. Most importantly, such formulation with polyR compared favorably to commonly used adjuvants such as CFA.
Several reports have shown that adding CpG ODN to protein-based vaccines can boost antibody production (Klinman et al., 1999; Weeratna et al., 2003; Bourquin et al., 2008) and increase the number of spleen cells actively secreting IFN-gamma in vivo (Klinman et al., 1999). In our model, we did not observe such a clear adjuvant effect of CpG ODN alone after a single immunization. The more limited impact of CpG ODN in our model might be related to the dose used in our experiments (10 μg), which is lower than those applied in other studies (50–100 μg) (Klinman et al., 1999; Bourquin et al., 2008). Our data are actually in line with another report in which the adjuvant effect of CpG ODN also appeared more limited when used at the same dose, that is, 10 μg (Kovacs-Nolan et al., 2009). This relatively low CpG ODN dosage (0.5 mg/kg in mice), was chosen to fit with the doses commonly used in clinical trials (<0.3 mg/kg per subcutaneous injections), and to minimize the potential immunological bias created by systemic diffusion of CpG ODN in our mice.
Polycations can act as adjuvants on their own. Indeed, the addition of a polycation alone to the OVA antigen was able to induce an antibody response after a single immunization, independently of the nature of the polycation. This is in agreement with other publications showing that cationic peptides can enhance the delivery of antigens to cells and APCs (Buschle et al., 1997; Mitchell et al., 2000; Futaki et al., 2001; Fritz et al., 2004). Successful immunizations in vivo have been achieved with different polycationic formulations (Schmidt et al., 1997; Lührs et al., 2002; Borges et al., 2008). However, in spite of their abilities to trigger an IgG response, polycations alone only poorly induced CD8+ T lymphocytes or a high IgG2:IgG1 ratio in our experiments.
The association of polycations and CpG ODN further enhanced the antibody response, especially when polyR was used. Moreover, when combined to CpG ODN, polyR and, to a slightly lesser extent, polyH significantly increased the number of IFN-gamma-secreting splenocytes, whereas the other polycations were less, or not efficient. This successful combination of CpG ODN and polyR is in line with a publication using an MHC class I-restricted peptide (Lingnau et al., 2002). Our study further extends this latter study, showing that CpG ODN-polyR complex allows a full protein antigen, and not only a predesigned MHC class I-peptide, to be processed into APCs and to be presented on both MHC class I and class II molecules. In addition, we showed that polyH is also efficient.
The significant difference in the ability of the various polycations to induce a cellular immune response when combined to CpG ODN was unexpected. Some studies have shown that all polycations do not share the same ability to cross the cell membranes (FUTAKI, 2005). Peptides containing basic amino-acids have been reported to have translocation activity, especially those rich in arginine. The number of arginine residues should be higher than 5 for optimal translocation (Futaki et al., 2001; Mitchell et al., 2000). Polyarginine containing 6 or more amino acids entered cells more effectively than corresponding polymers of lysine, ornithine, and histidine (MITCHELL, 2000). To date, no specific receptor for polyR has been identified so far, and the mechanisms of internalization are still unclear but seem to rely on endocytosis and macropinocytosis (Nakase et al., 2004). Buschle et al. (1997) suggested that the differences observed in peptide uptake by dendritic cells between polyR and polyK could be explained by their means of entry into the cells, that is, endocytosis and transient permeabilization, respectively. In addition, it was reported that polyK, especially high-molecular-weight forms, may elicit apoptosis in antigen-presenting cells—APC (Buschle et al., 1997)—thus accounting for a weaker immune response. Surprisingly, we found that the type of polycation also strongly interferes with the ability of the particles to entrap the OVA antigen. Most of the OVA antigen was found in the particles with polyR or polyH, but remained in the supernatant with polyK or chitosan. These data were unexpected, and the underlying mechanisms are unclear. One explanation could be that the simple ammonium residues, as encountered in chitosan and polyK, are not sufficient to efficiently bind ovalbumin, whereas in polyR and PolyH, the positive charges are delocalized, enabling the formation of multiple H-bonds. It is well known that immune responses are improved by chemically coupling CpG ODN to the antigen (Cho et al., 2000; Shirota et al., 2001; Maurer et al., 2002; Heit et al., 2005; Zhang et al., 2007) or by loading proteins on microparticles (Standley et al., 2007; Bourquin et al., 2008; Schlosser et al., 2008; Zwiorek et al., 2008), suggesting that optimal immune stimulation occurs when the antigen and the adjuvant are presented in proximity. It can therefore be hypothesized that both (a) the ability to efficiently cross the cell membranes and (b) co-absorption of the antigen and the CpG ODN within polycationic particles are required to trigger a strong cellular immune response, which are 2 criteria best achieved by polyR.
One striking new finding of our study is that the CpG ODN-polyR complex compared favorably to Alum, CpG ODN-IFA, and CFA for priming both B-cell and CD8+ T-cell immune responses. Water-in-oil emulsions, as CFA, are among the most potent adjuvants leading immune responses toward Th1, but are associated with a significant local toxicity. CpG ODN-polyR complexes, which are not water-in-oil emulsions, do not harbor such limitations, and their efficiency, when compared to CFA or CpG ODN-IFA, is therefore promising for further clinical developments.
In conclusion, our results demonstrate that the combination of CpG ODN and poly-arginine is the best adjuvant formulation for efficiently enhancing both antigen-specific humoral and cell-mediated responses. This CpG ODN-polyR complex offers a number of advantages over other currently used adjuvants: (i) all components are synthetics and the particles are easy to prepare, (ii) both antigen-specific antibody-mediated and cell-mediated immune responses are induced, and (iii) this adjuvant is at least as efficient as CFA, without the local toxicity associated with water-in-oil emulsions.
Footnotes
Acknowledgments
The authors would like to thank Martine Rancic and Elizabeth Huc for the handling and care of the animals, Federico Sandoval for technical assistance, and professor Eric Tartour for helpful discussions. This work was supported by grants from the Association Oligocyte, the AP-HP (Assistance Publique-Hôpitaux de Paris), and the ADNA (Association pour le Développement des Neurosciences à Avicenne).
Author Disclosure Statement
Potential conflict of interest: the Assistance Publique–Hôpitaux de Paris and AF Carpentier hold a patent position in CpG ODN.