
Abstract
We here demonstrate for the first time that 5-carboxytetramethylrhodamine (TAMRA) covalently linked to nuclear localization signal (NLS)-conjugated peptide nucleic acids (PNAs) are photosensitizers (PSs) with the capacity to initiate photochemical damage to endocytic membranes, resulting in release of endocytosed material into cytosol. Our results show that TAMRA/PNA/NLS conjugates work as multifunctional molecules by offering cellular uptake, PNA-directed gene silencing, and the possibility for targeting in a light-controlled manner. In addition to PNA-directed gene silencing, we demonstrate that TAMRA/PNA/NLS molecules may function as a PS for light-controlled release of small interfering RNA molecules from the endocytic pathway when combined with an appropriate carrier. Using these strategies, we could silence the S100A4 gene at both protein and mRNA levels in a light-controlled manner, without any detectable reduction in cell viability. Our data demonstrate the possibility for light-controlled delivery of macromolecules entrapped within endocytic vesicles using multifunctional TAMRA/PNA/NLS molecules as PSs.
Introduction
There is a long history of research dealing with the spectroscopic properties of xanthene dyes, starting with the synthesis by von Baeyer of fluorescein in 1871 (Rompp and Raaf, 1982). As PSs, xanthenes such as rose Bengal, eosin, and rhodamine 6G have been found effective in the production of ROS. Interestingly, the commonly used red fluorescent dye tetramethylrhodamine methyl ester (TMRM) has been found to produce ROS in mitochondria, leading to phototoxicity after 546 nm excitation (Zorov et al., 2000). In vivo, the spectral properties of rhodamines restrict their use to thin lesions because of the limited penetration of light. Alkylation or halogenation of amino groups introduces, however, some red shift to the light absorption spectra (Diwu et al., 1993). Of note is that a dibrominated rhodamine derivative has been granted orphan drug designation by the U.S. Food and Drug Administration (FDA) to treat patients with chronic myeloid leukemia (CML) ex vivo (VILLENEUVE, 1999).
Here, we investigate the photosensitizing properties of 5-carboxytetramethylrhodamine (TAMRA) molecules. Our results show evidence for both peptide nucleic acid (PNA) and small interfering RNA (siRNA) delivery into cytosol after 546 nm illumination of TAMRA/PNA/nuclear localization signal (NLS) molecules followed by effective gene silencing of S100A4 mRNA and protein levels. Our results demonstrate for the first time the possibility for light-controlled delivery of macromolecules entrapped within endocytic vesicles using TAMRA/PNA/NLS molecules as PSs.
Materials and Methods
Cell line and culture conditions
The SSX cell line was established from a metastatic lesion of malignant melanoma in a patient treated at the Norwegian Radium Hospital (Aamdal et al., 1984). The osteosarcoma cell line, OHS, was established from a bone tumor biopsy (Fodstad et al., 1986), and SK-MEL-28 (human melanoma) cells were obtained from the American Type Culture Collection (ATCC). All cells were cultured and maintained in RPMI 1640 medium (BioWhittaker), supplemented with 10% fetal calf serum (PAA) and GlutaMAX (Gibco, Invitrogen). The cells were maintained at 37°C in a humidified atmosphere containing 5% CO2 and routinely tested for Mycoplasma infections (VenorGeM; Minerva Biolabs), and the cell line identities were also verified by short tandem repeat analysis. Importantly, cells were light-protected by aluminum foil under all experiments to avoid unwanted PS activation.
Molecules
Unconjugated TAMRA molecules and TAMRA conjugated to either NLS or PNA/NLS molecules were purchased from PANAGENE and are summarized in Table 1. A PNA molecule (S100A4ATG TAMRA/PNA/NLS) containing a complementary sequence against the ATG start codon of the S100A4 gene (sequence: 5-GAGGGCACGCCATGACAGCA-3) in addition to a scrambled (scr.) PNA molecule (scr. TAMRA/PNA/NLS) with sequence 5-GCCACATGCGACGCAGAGAG-3 were all selected based on prior work (Boe and Hovig, 2006). For the S100A4ATG PNA binding site, see Supplementary Fig. S1 (Supplementary Data are available online at www.liebertonline.com/nat). All molecules were HPLC-purified and characterized by mass spectroscopy. Stock solutions (1 mM) were made in sterile water and further diluted into working solutions of 100 μM. Absorption spectra of S100A4ATG TAMRA/PNA/NLS molecules were measured using a NanoDrop (ND-1000) fluorospectrometer (Supplementary Fig. S2A). Chloroquine diphosphate salt was obtained from Sigma. For the S100A4 siRNA transfection experiments, siRNA molecules (S100A45′UTR siRNA) directed against the 5′UTR of the S100A4 mRNA sequence (GenBank accession No. NM_002961) were ordered from Eurogentec. The S100A45′UTR siRNA molecules (481–499; 5-UGAGCAAGUUCAAUAAAGA-3, 3-ACUCGUUCAAGUUAUUUCU-5) were chosen based on effective gene silencing in previous studies (Boe et al., 2007, 2008, 2010). For the S100A45′UTR siRNA binding site, see Supplementary Fig. S1. In addition to S100A4 gene silencing, we also used MEK1 siRNA molecules directed against the MEK1 mRNA sequence (GenBank accession No. NM_002755.3) to demonstrate that the scr. TAMRA/PNA/NLS molecules have the capacity to work as a PS for siRNA delivery. The MEK siRNA molecules (1534–1552; 5-AGCAUGAACCAUGAGUUGC-3, 3-UCGUA CUUGGUACUCAACG-5) were chosen based on effective gene silencing in a previous study (Zhang et al., 2010). S100A45′UTR and MEK1 siRNA molecules were transfected with 25-kDa branched polyethylenimine (B-PEI), obtained from Sigma (St. Louis, MO). B-PEI was diluted in sterile water before use.
The PNA sequences are written in a 5′–3′ direction, and the amino acids are written in uppercase as they are in L-form and the NLS peptide have a C-terminal amide. O is an AEEA (aminoethoxyethoxy acetyl) linker.
NLS, nuclear localization signal; PNA, peptide nucleic acid; TAMRA, 5-carboxytetramethylrhodamine.
546 nm excitation with LumiSource lamp
When illuminated, cells were exposed to light with a LumiSource prototype (PCI Biotech AS), containing a bank of 4 fluorescent tubes for 546 nm excitation (Philips TL-D 18W/Green SLV) (Supplementary Fig. 2B). Cells were then exposed to green light at 11.5 mW/cm2 for various durations.
Live-cell fluorescence microscopy
Three different cell lines, OHS, SSX, and SK-MEL-28, were cultivated for 48 hours in 12-well plates prior to use. Unconjugated and conjugated TAMRA molecules listed in Table 1 were added to each well at different concentrations and reincubated for 24 hours prior to analysis. TAMRA was then excited at 546 nm by the live-cell fluorescence microscope at different time intervals to monitor relocalization of molecules after damage to the endocytic vesicles. A Zeiss inverted microscope, an Axiovert 200 (equipped with Rhodamine 546/12 nm BP excitation filter, a 580 nm FT beamsplitter, and a 590 nm LP emission filter), was used for excitation and detection of TAMRA fluorescence. Pictures were composed by the use of Carl Zeiss AxioCam HR, Version 5.05.10, and AxioVision 3.1.2.1 software.
Transfection
Cells were cultivated for 48 hours in 12-well plates prior to use. Transfections were performed by adding TAMRA/PNA/NLS or TAMRA/NLS molecules (listed in Table 1) at various concentrations to cells in serum-containing medium. Some of the experiments were additionally transfected with S100A45′UTR or MEK1 siRNA complexed with B-PEI at a nitrogen/phosphate (N/P) ratio of 3, making a final concentration of 50 nM. Of note, siRNA molecules were complexed with B-PEI in serum-free medium for 30 minutes before combining with the transfection solution containing either S100A4ATG TAMRA/PNA/NLS molecules or scr. TAMRA/PNA/NLS molecules. After transfection for 18 hours, cells were washed 3 times with fresh medium and reincubated for 4 hours before illumination at 546 nm for different durations, depending on the experiment. After illumination, cells were incubated for various time points prior to analysis by MTS analysis and/or harvesting for RNA or protein. For all experiments, samples were calculated as percentage of untreated control cells without 546 nm illumination. All experiments were performed 3 times (3 individual experimental parallels).
Cell viability after 546 nm excitation
Cells were cultivated for 48 hours in 12-well plates prior to use. Cells were transfected as described under the Transfection section and incubated for 96 hours after 546 nm illumination. After incubation, cells were washed once with RPMI 1640 medium before MTS (3-(4.5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium) (Promega) analysis. Absorbance was measured at 490 nm in wells with serum-containing medium diluted with MTS to a final volume of 1 mL/well (1:5 dilution).
Western immunoblotting
Preparation of whole-cell lysates was performed by addition of lysis buffer [150 mM NaCl and 50 mM Tris-HCl (pH 7.5)], 0.1% NP-40, and 10 μg/mL of each leupeptin hemisulfate, aprotinin, pepstatin A, and PMSF to dry cell pellets, left on ice for 1 hour, sonicated, and centrifuged to remove cell debris. The supernatant (protein lysate) was removed from the cell debris and the amount of total protein was quantified by standard Bradford protein assay (Bio-Rad Laboratories, Inc.). Protein lysates (22 μg) were separated on 4%–12% NuPAGE® Novex Bis-Tris Gels (Invitrogen) and blotted using an iBlot Dry Blotting system (Invitrogen) according to the manufacturer's instruction. The membranes were subsequently incubated in 20 mM Tris-HCl (pH 7.5), containing 0.5 M NaCl and 0.25% Tween 20 (TBST) with 10% dry milk (blocking solution) before incubation with rabbit polyclonal anti-S100A4 (diluted 1:300; DAKO) and mouse monoclonal anti-α-tubulin (diluted 1:250; Amersham Life Science) in TBST containing 5% dry milk. After washing, the immunoreactive proteins were visualized using horseradish peroxidase-conjugated secondary antibodies (diluted 1:5000; DAKO) and the SuperSignal chemiluminescence detection system (Pierce), by means of a G:Box iChemi Chemiluminescence Image Capture (Syngene).
Real-time reverse transcriptase–PCR of S100A4
Total cellular RNA was isolated with the GenElute Mammalian Total RNA Miniprep Kit (Sigma-Aldrich) and the iScript cDNA synthesis kit (Bio-Rad Laboratories, Inc.) was used for reverse transcription. Both kits were used according to the manufacturers' manuals. Real-time detection was obtained by use of SYBR Green I. For each PCR, 10 μL cDNA, 30 μL iQ SYBRGreen Supermix (Bio-Rad Laboratories, Inc.), 300 nM of each primer, and nuclease-free water were added to a final volume of 60 μL. Samples of 25 μL each were then applied to the PCR plate. This method ensures that the parallels are true parallels and that there is sufficient PCR mix for parallels. Primers were designed with the Primer Express software from Applied Biosystems (Applied Biosystems). The primer set used for S100A4 (forward primer 5′-AAGTTCAA GCTCAACAAGTCAGAAC-3′ and reverse primer 5′-CATCT GTCCTTTTCCCCAAGA-3′) and MEK1 (forward primer 5′-TTCTACAGCGATGGCGAGAT-3′ and reverse 5′-TCCAG CTTTCTTCAGGACTTG-3′) amplifies the 79- and 81-bp segments that span the intron between exon 2 and 3, and 3 and 4, respectively. Real-time reactions were run on an iCycler (Bio-Rad Laboratories, Inc.) with the following amplification protocol: 3 minutes initial denaturation at 95°C, 50 cycles of 10 seconds denaturation at 95°C and 35 seconds annealing/extension at 60°C, one hold at 95°C for 20 seconds followed by a hold for 1 minutes at 55°C, and finally, a melt curve analysis of 80 steps each for 10 seconds, with 0.5°C increase until a final temperature of 95°C. The quality of the RNA samples was verified by amplification of 2 housekeeping genes, the TATA-binding protein with forward primer 5′-GCCCGAAACG CCGAATAT-3′ and reverse primer 5′-CGTGGCTCTCTTA TCCTCATGA-3′ and the human acidic ribosomal phosphoprotein PO with forward primer 5′-CGCTGCTGAACATG CTCAAC-3′ and reverse primer 5′-TCGAACACCTGCTGG ATGAC-3′. These housekeeping genes were chosen as they were unaffected by the different treatment modalities in this study (data not shown). The Gene Expression Macro, version 1.1 (Bio-Rad Laboratories, Inc.), was used for the quantitative calculations. The program performs calculations based on the ΔΔCT method (Vandesompele et al., 2002), which allows comparison of cycle threshold values obtained using different sets of primers on the same set of samples.
Results
Uptake and intracellular distribution of TAMRA molecules
We investigated the intracellular distribution of unconjugated TAMRA, TAMRA/NLS, and TAMRA/PNA/NLS molecules (listed in Table 1) by live-cell fluorescence microscopy. The NLS peptide was used to improve cellular uptake, as a +5 net charge have previously shown to be superior for cellular uptake of PNA conjugates compared with PNA molecules containing a peptide with less cationic charge (Boe and Hovig, 2006). Our results implied an endocytic uptake pattern of TAMRA/NLS and S100A4ATG TAMRA/PNA/NLS molecules, whereas unconjugated TAMRA molecules were distributed differently (Supplementary Fig. S3). Of note, a comparable intracellular distribution to S100A4ATG TAMRA/PNA/NLS molecules was observed when using scr. TAMRA/PNA/NLS molecules (data not shown). Surprisingly, we observed an illumination time-dependent relocation of TAMRA fluorescence in cells treated with 1 μM S100A4ATG TAMRA/PNA/NLS molecules after 546 nm illumination from the microscopy (Fig. 1). We when tested 3 different cell lines treated with 1 μM S100A4ATG TAMRA/PNA/NLS molecules alone, all showed TAMRA fluorescence distributed in cytosol and nucleus following 546 nm illumination (Fig. 2). Importantly, all molecules containing TAMRA in this study were added to cells for spontaneous cellular uptake without any additional transfection agent.
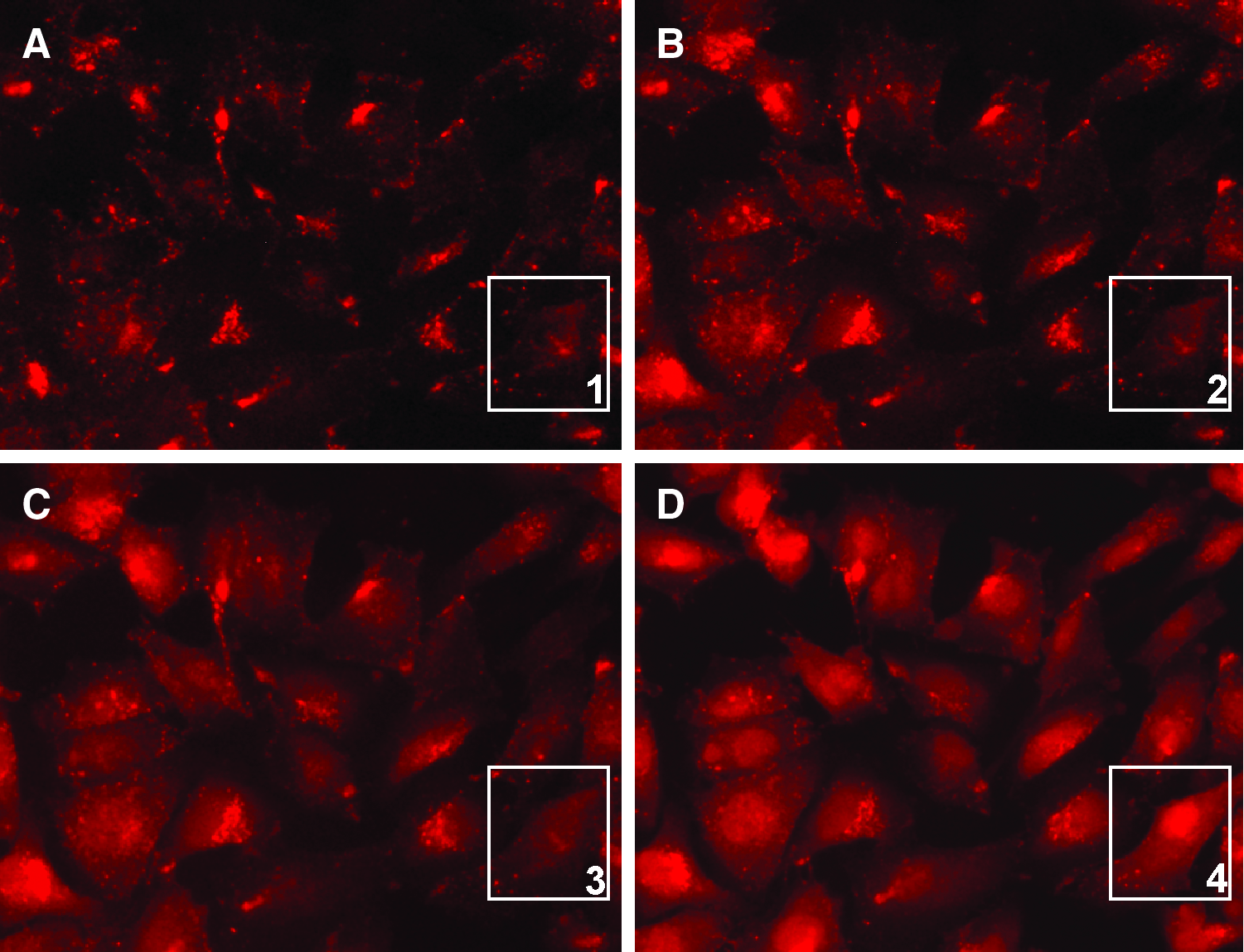
Live-cell fluorescence microscopy pictures (32×) show a time-course study of SSX cells treated with 1 μM S100A4ATG TAMRA/PNA/NLS molecules and illuminated at 546 nm by using the microscopy as an excitation source.
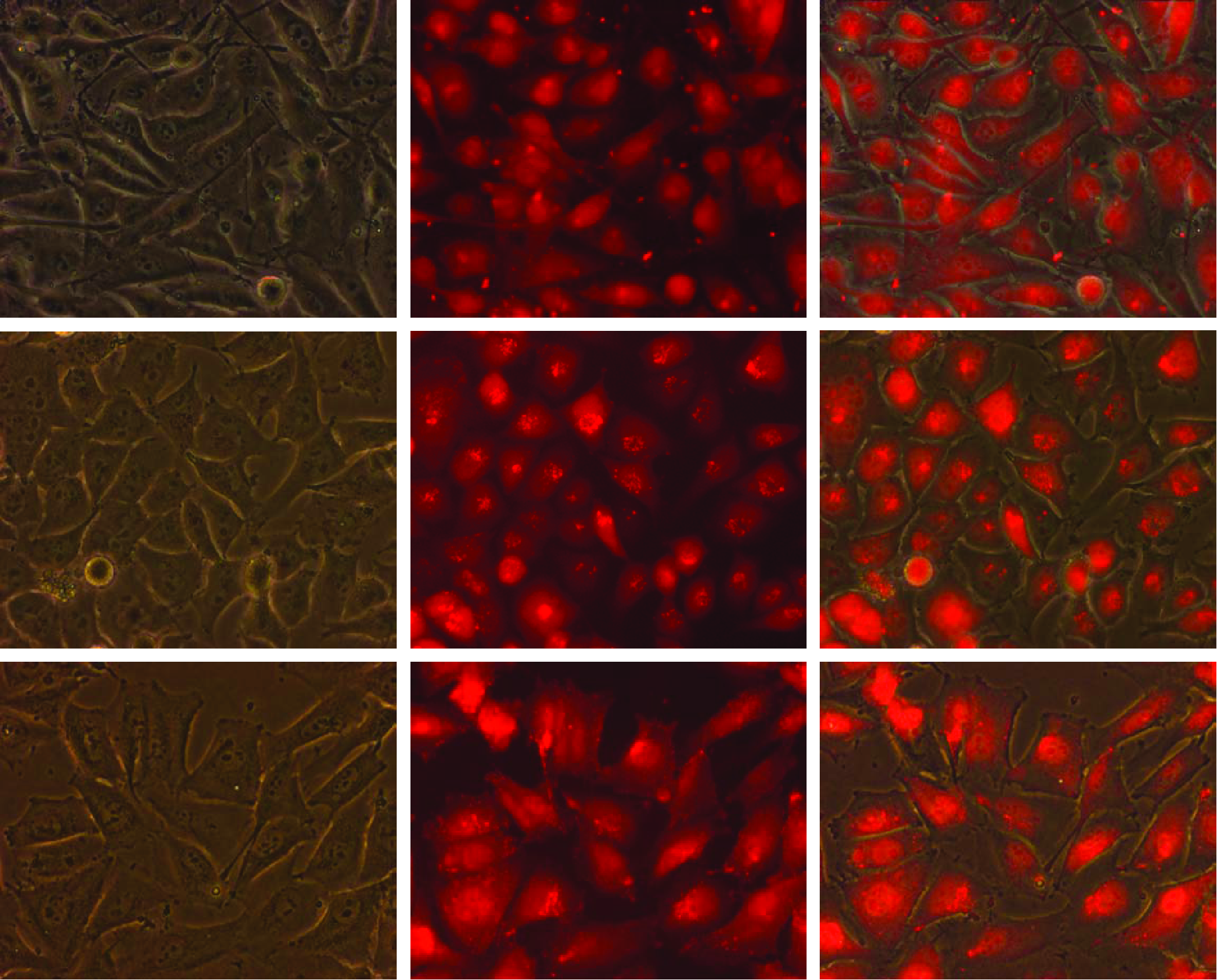
Live-cell fluorescence microscopy pictures (32×) show cystolic and nuclear localization of TAMRA fluorescence after 546 nm illumination in the SK-MEL28, OHS, and SSX cell lines (from upper to lower panel) using 1 μM S100A4ATG TAMRA/PNA/NLS molecules. Pictures from left to right show phase-contrast image, fluorescence image, and merge between phase contrast and fluorescence. Color images available online at www.liebertonline.com/nat
Light-controlled gene silencing at the protein level using S100A4ATG TAMRA/PNA/NLS
To confirm that PNA molecules are liberated from endosomal and lysosomal compartments and not only the TAMRA fluorescent part of the conjugated molecule, S100A4 gene silencing was measured at 96 hours after 546 nm illumination of OHS cells incubated with various concentrations of S100A4ATG TAMRA/PNA/NLS molecules (Fig. 3A–C). Using 0.5 or 1 μM S100A4ATG, TAMRA/PNA/NLS molecules did not result in a very effective S100A4 gene silencing after 12 minutes of illumination (Fig. 3A, B). However, S100A4 protein levels were clearly decreased when using 1 μM compared with 0.5 μM. When increasing the S100A4ATG TAMRA/PNA/NLS concentration to 2 μM, we measured a decrease in S100A4 protein levels to remaining 10% (compared with untreated control levels without illumination) after 12 minutes of illumination. Decreasing the illumination time to 9 minutes did not reduce gene silencing efficiency, but lowering of the illumination time to 6 minutes reduced the gene silencing efficiency from around 90% down to 80% (compared with untreated control without illumination). No major decrease (<5%) in S100A4 protein levels was detected in untreated cells or in cells treated with 2 μM scr. TAMRA/PNA/NLS molecules, either with or without illumination. Further, no major decrease (<10%) in S100A4 protein levels was detected in cells treated with 2 μM S100A4ATG TAMRA/PNA/NLS molecules without illumination. Cell viability in the experiment using the highest S100A4ATG TAMRA/PNA/NLS concentration (2 μM) was measured by the MTS assay. Results showed no major (<10%) reduction in cell viability after 96 hours, regardless of treatment modality (Fig. 3D).

PNA-induced S100A4 protein gene silencing and cytotoxicity in the OHS cell line at 96 hours after 546 nm illumination.
Light-controlled gene silencing at the mRNA level using S100A4ATG TAMRA/PNA/NLS
Before we evaluated the possibility for PNA-induced mRNA gene silencing, we investigated whether the S100A4ATG TAMRA/PNA/NLS binding site at the S100A4 mRNA molecules could influence the quantification of S100A4 mRNA levels after real-time RT-PCR analysis. We therefore spiked an untreated sample with different concentrations of S100A4ATG TAMRA/PNA/NLS molecules after RNA harvesting, but before the cDNA synthesis and real-time RT-PCR quantification. Our results showed that none of the concentrations tested (2 μM, 2 nM, and 2 pM) had any influence upon the quantification of the S100A4 mRNA levels (Supplementary Fig. S4). This result implies that S100A4ATG TAMRA/PNA/NLS molecules do not influence the real-time RT-PCR quantification of S100A4 mRNA levels when bound to S100A4 mRNA molecules. Next, we addressed the capacity of 2 μM S100A4ATG TAMRA/PNA/NLS molecules to silence S100A4 mRNA levels in OHS cells at 24 hours after 546 nm illumination (Fig. 4A). S100A4 mRNA levels were down to around 80% and 45% (compared with untreated control cells without illumination) after 6 and 12 minutes of illumination, respectively. No major decrease (<5%) in S100A4 mRNA levels was detected in untreated control cells or cells containing 2 μM scr. TAMRA/PNA/NLS molecules after 6 and 12 minutes of illumination (compared with untreated control cells without illumination). Then, we increased the incubation time to 96 hours instead of 24 hours. Our results showed no further decrease of S100A4 mRNA levels when increasing the incubation time from 24 to 96 hours, rather a minor increase in S100A4 levels was observed (Fig. 4B).

PNA-induced S100A4 mRNA gene silencing in the OHS cell line at 24 and 96 hours after 546 nm illumination.
Light-controlled delivery of siRNA molecules using different TAMRA/PNA/NLS molecules
To address the universality of TAMRA as a PS when conjugated to PNA/NLS molecules, we investigated the possibility of delivering S100A45′UTR siRNA molecules by the PEI carrier in combination with either 2 μM S100A4ATG or scr. TAMRA/PNA/NLS molecules. When delivering siRNA molecules, endocytic entrapment prior to illumination of PS was necessary to achieve a light-controlled delivery effect. This was accomplished by forming siRNA/PEI complexes with a very low N/P ratio, sufficient for siRNA delivery into cells, but inadequate for release into cytosol. Of note, the endosmolytic capacity of B-PEI increases with growing monomer size and amount of PEI used (see the Discussion section). Our results showed a decrease in S100A4 mRNA levels down to around 30% (compared with untreated control without illumination) after 12 minutes of illumination in samples transfected with S100A45′UTR siRNA/PEI complexes and 2 μM scr. TAMRA/PNA/NLS molecules (Fig. 5). An additional decrease in S100A4 mRNA levels down to around 10% (compared with untreated control without illumination) after 12 minutes of illumination was measured in samples when using S100A45′UTR siRNA/PEI complexes and 2 μM S100A4ATG TAMRA/PNA/NLS molecules (Fig. 5). These results demonstrate the capacity of TAMRA/PNA/NLS molecules to work as a PS for siRNA delivery independent of the PNA targeting sequence. Importantly, no major decrease (<5%) in S100A4 mRNA levels was measured in cells treated with S100A45′UTR siRNA/PEI and illumination (compared with untreated control without illumination). Further, complexing TAMRA/NLS molecules in different concentrations (1–10 μM) directly with S100A45′UTR siRNA (50 nM), or transfected separately as TAMRA/NLS (1–10 μM) and S100A45′UTR siRNA/PEI (50 nM), did not result in any measurable siRNA gene silencing under the 3 illumination times (10, 15, and 20 minutes) tested (Supplementary Fig. S5). The latter results demonstrate that only TAMRA conjugated to PNA molecules have the capacity to work as a PS for siRNA delivery under the parameters tested here.

siRNA-induced S100A4 mRNA gene silencing in the OHS cell line at 24 hours after 546 nm illumination. Bars represent control cells (cells treated with only S100A45′UTR siRNA), cells treated with S100A45′UTR siRNA and 2 μM scrambled (scr.) TAMRA/PNA/NLS molecules, and cells treated with S100A45′UTR siRNA and 2 μM S100A4ATG TAMRA/PNA/NLS molecules with 6 or 12 minutes of 546 nm illumination. Bars represent percentage of untreated control cells (without light and without siRNA). Three individual experiments are presented, with error bars showing standard error of the mean±SEM. siRNA, small interfering RNA.
To further validate the possibility of using TAMRA/PNA/NLS molecules with a random and nonspecific PNA sequence as a PS for siRNA delivery, we investigated scr. TAMRA/PNA/NLS molecules as PS for MEK1 gene silencing. MEK1 siRNA molecules show a comparable gene silencing effect to S100A45′UTR siRNA molecules, with a corresponding decrease in MEK1 mRNA levels down to around 30% (compared with untreated control without illumination) after 12 minutes of illumination (Fig. 6).
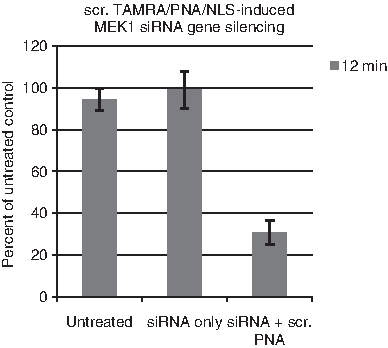
Scrambled (scr.) TAMRA/PNA/NLS induced MEK1 siRNA gene silencing in the OHS cell line at 24 hours after 546 nm illumination. Bars represent untreated cells, cells treated with only MEK1 siRNA, and cells treated with MEK1 siRNA in combination with 2 μM scrambled (scr.) TAMRA/PNA/NLS molecules after 12 minutes of 546 nm illumination. Bars represent percentage of untreated control cells (without light). Three individual experiments are presented, with error bars showing standard error of the mean±SEM.
Discussion
We have here explored the possibility of using various TAMRA molecules as PSs for light-controlled delivery of molecules entrapped in the endocytic pathway. We found that TAMRA/PNA/NLS molecules have the capacity to initiate photochemical damage to endocytic membranes, resulting in release of endocytosed material into cytosol. A prerequisite for light-controlled delivery of molecules is an endocytic uptake route of PS and the given molecule. Our microscopy results implied that both TAMRA/NLS and TAMRA/PNA/NLS molecules were entrapped in endocytic vesicles after cellular uptake in a comparable manner as FITC/PNA/NLS molecules in our previous study (Boe and Hovig, 2006). An endocytic uptake mechanism of PNA/NLS molecules is further supported by Nielsen and colleagues (Shiraishi et al., 2005; Shiraishi and Nielsen, 2006). In contrast to TAMRA/NLS and TAMRA/PNA/NLS molecules, unconjugated TAMRA molecules showed a different distribution within cells. Noteworthy, unconjugated TAMRA molecules have been previously reported to localize to mitochondria in a similar manner to tetramethylrhodamine ethyl ester and TMRM (Scaduto and Grotyohann, 1999). Although TAMRA/NLS molecules localized to endocytic compartments, no gene silencing was measured when added to the cells together with S100A45′UTR siRNA/PEI complexes under the tested parameters. However, our microscopy results did imply endocytic release of TAMRA fluorescence induced by 546 nm illumination in some OHS cells when using very high concentrations of TAMRA/NLS molecules (data not shown). Of importance, a much weaker TAMRA fluorescence signal was detected in OHS cells containing TAMRA/NLS compared with TAMRA/PNA/NLS under the same concentration. Reduced sedimentation due to lower molecular weight combined with NLS degradation before and after endocytosis could explain the observed results.
The ability of TAMRA/PNA/NLS molecules to initiate photochemical damage to endocytic membranes and the possibility to release endocytosed material into cytosol were measured by the effect of S100A4 and MEK1 gene silencing after TAMRA/PNA/NLS induced siRNA delivery. In these studies, the 2 separate production batches of scr. and S100A4ATG TAMRA/PNA/NLS molecules used in this study demonstrated that the photosensitizing effect did not seem to be the result of a flawed production procedure of synthesized molecules. When delivering siRNA molecules, care was taken to obtain high endosomal entrapment prior to illumination of the PS. The endosmolytic capacity of B-PEI in combination with S100A45′UTR siRNA molecules under various N/P ratios have been extensively validated in our previous study (Boe et al., 2008).
Interestingly, the use of S100A4ATG TAMRA/PNA/NLS molecules alone resulted not only in effective S100A4 protein gene silencing, but also in a moderate and specific S100A4 mRNA gene silencing. As S100A4 cDNA amplification from the real-time RT-PCR procedure was not affected by the binding of PNA to mRNA (as shown in Supplementary Fig. S4), the result implies a pretranslational inhibition mechanism. Although a mixed-sequence PNA has been reported to invade duplex DNA, resulting in transcriptional inhibition (Cutrona et al., 2000, 2003; Cogoi et al., 2005), we did not detect any further decrease in S100A4 mRNA levels after 96 hours. Binding of PNA to immature heterogeneous nuclear RNA inside the nucleus, followed by rapid sequence specific degradation before or after nuclear export due to unnatural folding, is a plausible explanation besides transcriptional inhibition. Further investigations are required to disclose the gene silencing mechanism(s) with respect to the observed decrease in S100A4 mRNA levels when using our S100A4ATG TAMRA/PNA/NLS molecule.
Up to 90% S100A4 protein gene silencing in the OHS cell line was also measured in our previous PNA study when using TPPS2a as PS for light-directed delivery (Boe and Hovig, 2006). Of note, this was achieved by using a lower PNA dose (1 μM) and irradiation dose (420 mJ/cm2) than in this work. However, an advantage of using TAMRA compared with TPPS2a is the possibility to achieve up to 90% protein silencing without any measurable reduction in cell viability, in contrast to a 30%–45% reduction as shown in our previous study. TPPS2a localizes to membranes of endocytic vesicles, whereas TAMRA are more likely to be present in the lumen of endocytic vesicles similar to, for example, TPPS4 because of the chemical composition of the PS. This divergence in PS localization is most likely an important factor for the efficacy of the PS to initiate endocytic membrane damage, as previously measured when comparing TPPS2a and TPPS4 (Prasmickaite et al., 2001). We here demonstrate that using PCI in combination with PNA is equally efficient with respect to protein gene silencing as the combination of PCI and siRNA molecules (Boe et al., 2007, 2008, 2010). However, when comparing the ability to deliver siRNA molecules into the cytosol, we only obtained a 70% decrease in mRNA levels when using scr. TAMRA/PNA/NLS as PS, which is 20% less than using S100A4ATG TAMRA/PNA/NLS molecules as PS for siRNA delivery, or a separate TPPS22 as in our previous work (Boe et al., 2007, 2008, 2010). These results imply that the additional 20% decrease in mRNA levels obtained in this work when using S100A4ATG TAMRA/PNA/NLS molecules is due to the gene silencing capacity of the PNA itself. Compared with earlier studies, we imply that TPPS2a is a more effective PS than TAMRA for siRNA delivery when using TAMRA/PNA/NLS molecules with a randomized PNA sequence.
The novel strategy described here opens up for the possibility of delivering a single multifunctional molecule, consisting of a module for cellular uptake (the NLS peptide), a module for endocytic escape in form of the photosensitizing properties of TAMRA, and a PNA module for specific gene silencing. The benefit of using a multifunctional molecule is 100% colocalization of PS and PNA to endocytic vesicles, thereby limiting nonspecific effects due to unwanted PS activation in nonilluminated areas. This is favorable in a clinical situation where effective targeting and specificity is mandatory for therapeutic success. A possible disadvantage of using rhodamine derivates such as TAMRA for in vivo applications is their relatively low excitation wavelengths (∼550 nm) compared with, that is, Amphinex, TPCS2a (∼652 nm). Low excitation wavelengths of PSs restrict their application to thin lesions because of the limited penetration of light through human tissue. Alkylation of amino groups or halogenation can, however, introduce some red shift in the light-absorption spectra (Diwu et al., 1993). An example is the ex vivo application of a dibrominated rhodamine derivative, which has been granted orphan drug designation by the FDA to treat patients with CML (VILLENEUVE, 1999). Further, there is reason to believe that also other types of PSs can be conjugated to, for example, PNA molecules or other types of small molecular therapeutic drugs to obtain higher excitation wavelengths that are more suitable for in vivo use.
In conclusion, we have here shown for the first time that TAMRA/PNA/NLS molecules have the ability to release molecules from the endocytic pathway to cytosol and/or nucleus. We foresee further exploration of TAMRA applications within the field of light-directed therapy.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.