
Abstract
Telomeres are important multifunctional nucleoprotein structures located at the ends of eukaryotic chromosomes. Telomerase regulates telomere elongation, and its activity is associated with tumorigenesis. Because the activity of telomerase can be inhibited by G-quadruplex (G4) formation (a four-stranded DNA with stacks of G-quartets formed by four guanines in a planar structure), the role of G4 in cancer therapy has attracted many research interests. We studied the effects of three natural alkaloids—tetrandrine, fangchinoline, and berbamine—on the stability and formation of telomere DNA G4 with circular dichroism melting spectroscopy (melting-CD), variable temperature ultraviolet (melting-UV), proton nuclear magnetic resonance spectroscopy (1H NMR), and molecular docking, and examined the relationships among the alkaloid structure and their activities. We further investigated their cytotoxicity with the 3-(4,5-dimethylthiazole-2-yl)-2,5-diphenyltetrazolium bromide assay (MTT) and flow cytometry (FCM). The results demonstrated that alkaloids increased G4 stability and induced its formation, which added structure diversity of G4-ligands. The results showed that –OH at R1, –OCH3 at R2, and  at R3 had higher stability than other substituent groups for these alkaloids. We also found a transition of antiparallel to parallel G4 as the temperature increased. The result indicated the possible advantage of parallel G4 in adversity. In addition, the alkaloids demonstrated a moderate cytotoxicity and proved to be cell cycle blocker in the G1 phase. These alkaloids have revealed promising potentials to be the agents for antitumor therapy.
at R3 had higher stability than other substituent groups for these alkaloids. We also found a transition of antiparallel to parallel G4 as the temperature increased. The result indicated the possible advantage of parallel G4 in adversity. In addition, the alkaloids demonstrated a moderate cytotoxicity and proved to be cell cycle blocker in the G1 phase. These alkaloids have revealed promising potentials to be the agents for antitumor therapy.
Introduction
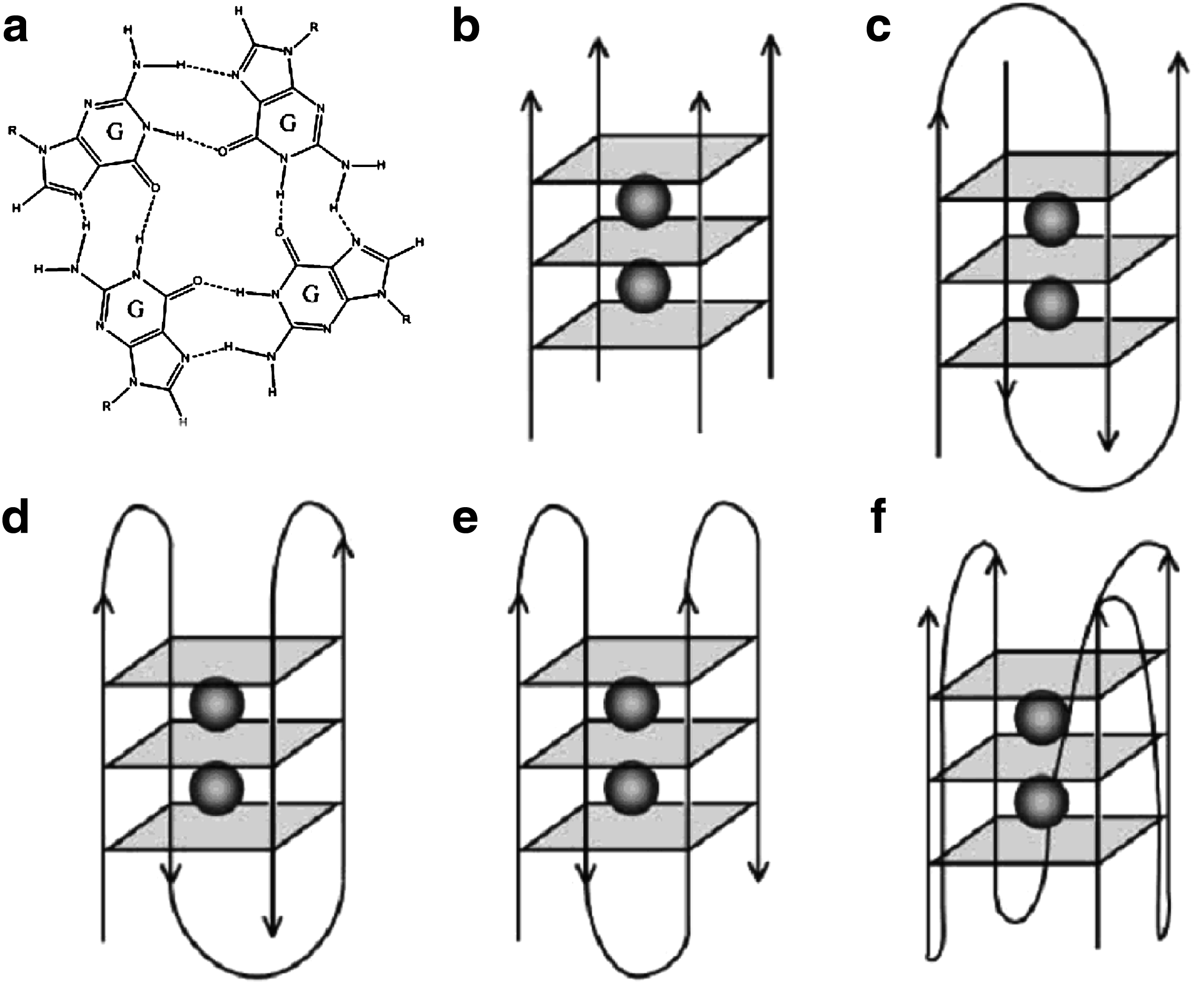
Samples schematic different types of G-quadruplex folds.
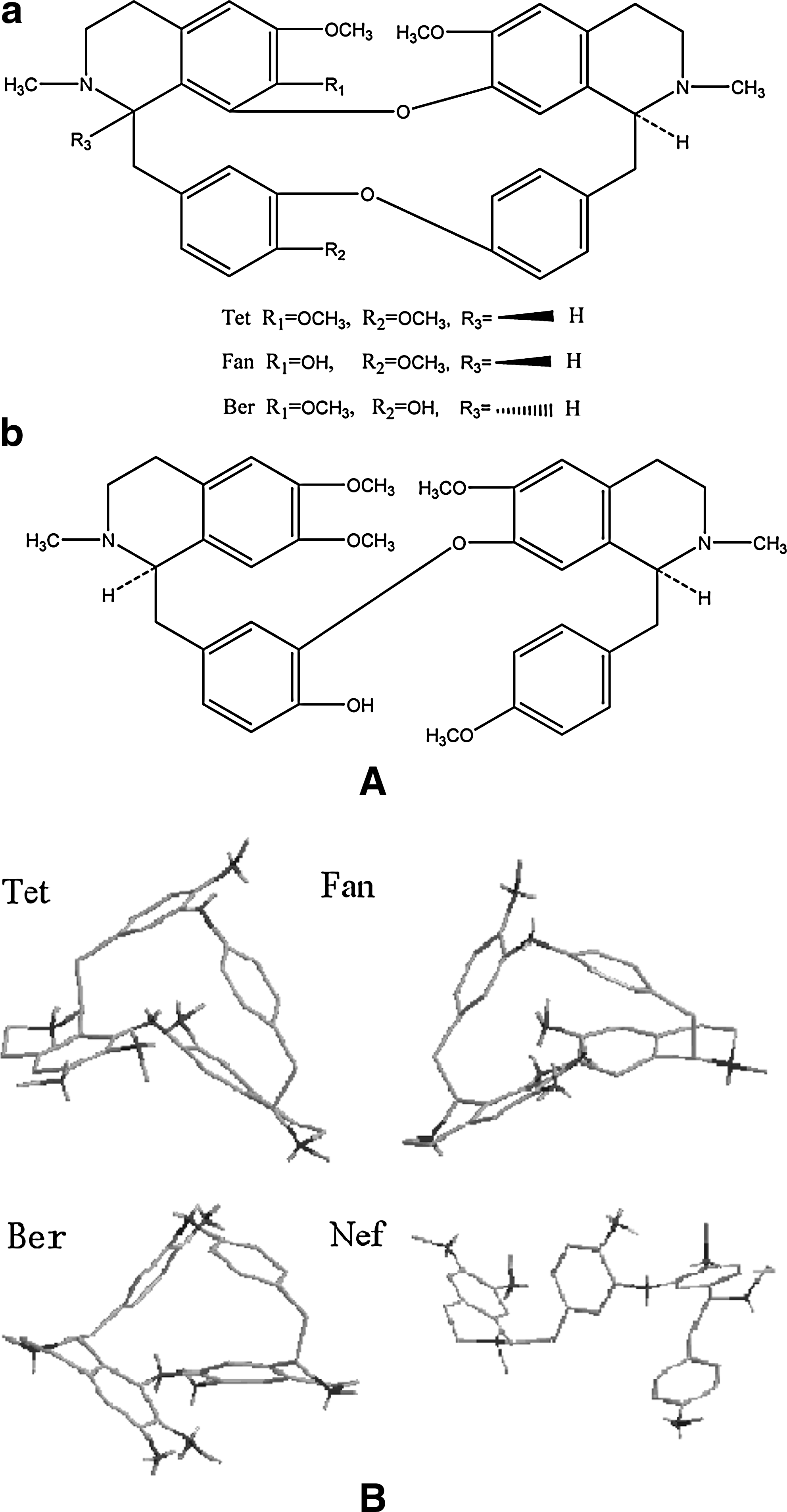
Telomeres are important multifunctional nucleoprotein structures at the ends of eukaryotic chromosomes that ensure the complete replication of chromosomal DNA and protect the chromosome ends from fusion and degradation (Wright et al., 1997). Telomeric DNA ends as the single strand (TTAGGG) n in vertebrates (Haider et al., 2003). Telomerase is the enzyme responsible for the elongation of telomeres in tumor cells. Telomerase shows high activity in 85% of immortal cancer cells, but remains silent in normal cells, suggesting a relationship between telomerase activity and cancer cells (Lingner et al., 1997). Increasing evidence shows that G4 ligands can effectively inhibit both the catalytic and capping functions of telomerase through stabilization of G4 (Phatak et al., 2007). Notably, although rich in the genome, G4 DNA formation potential is mainly found in human telomeric DNA and proto-oncogenes, but is low in tumor-suppressor genes (Eddy and MAIZELS, 2006), suggesting the possibility of treating cancer cells distinctively with effective G4-ligands. Therefore, a small-molecule approach has been devised to target telomeric G4 in cancer theropy. Efforts have been made to find effective G4-ligands from chemosynthesis and natural products. Some G4 ligands have been reported, but more therapeutically useful ligands are believed to exist. The natural products were proposed to be rich in G4 ligands for their high diversity and potential cytotoxicity. Here we investigated the interaction of telomeric DNA G4 with three natural alkaloids purified from traditionally used Chinese medicinal herb: Tetrandrine (Tet), fangchinoline (Fan), and berbamine (Ber) (Fig 1A).
Tet and Fan are bis-benzylisoquinoline alkaloids identified as the active ingredient in Radix Stephanae tetrandrae (Choi et al., 2000). Ber is a natural bis-benzylisoquinoline alkaloid purified from Berberis amurensis (Wang et al., 2007). Tet can suppress the production of interleukin-1 (IL-1) and tumor necrosis factor (TNF) function in DNA alkylation and nuclear factor-κB (NF-κB) inhibition (Efferth and Wink, 2010) and has inhibitory effects on many cancer cell types (Wong et al., 1992), including Neuro 2a mouse neuroblastoma cells (Jin et al., 2002) and Hep G2 cells (Wong et al., 1992; Kuo and Lin, 2003). Ber can exert antiproliferative effects on human hepatocellular carcinoma SMMC7721 cells (Wang et al., 2007), K562 cells (Wu et al., 2005), and others and can induce cell death in chronic myeloid leukemia cells (Xiao-Ying et al., 2007). Particularly, Ber can stimulate normal hematopoiesis in cancer patients undergoing chemotherapy or radiotherapy and is used to protect tumor patients from the cytotoxic effects of chemotherapeutic agents on bone marrow (Wang et al., 2007). Its derivatives have been synthesized as antileukemia agents (Xie et al., 2009). Moreover, Tet and Ber can reverse multidrug resistance (MDR) (CHEN, 2002), which increases efflux of anticancer drugs and is regarded as one of the huge obstacles for effective cancer therapy (Efferth and Wink, 2010).
In general, Tet and Ber could be potential effective and safe antitumor drugs. Tet exhibits high affinity in binding with c-myc G4 DNA (Liu et al., 2010). Ber displays antitelomerase activity with only 5% telomerase activity remaining and induced growth arrest and apoptotic cell death in HL-60 cells at the concentration of 10 mM (Ji et al., 2002). We propose that Fan has similar functions as Tet and Ber because they have the same core structure. These alkaloids have yet to be applied to clinical cancer treatments because their antitumor mechanisms are poorly understood.
In this article, we present preliminary results of the antitumor mechanism of Tet, Fan, and Ber. Considering the potential important role G4 plays in oncotherapy, we include a study of alkaloids and the stability and formation-inducing capability of telomere G4, as well as the cytotoxicity and function of cell cycle blocking. Furthermore, we investigated the structure–activity relationship. Our results demonstrated that Tet, Fan, and Ber are G4 ligands that could be promising agents for antitumor therapy.
Materials and Methods
Materials
d(T2AG3) (Hum6) and d((T2AG3)4) (Hum24) were purchased from Invitrogen (Beijing, China) and purified by polyacrylamide gel electrophoresis (PAGE). Tet, Fan, Ber, and nefnerine (Nef) were purchased from Shenzhen Medherb Biotechnology Co., Ltd. Heavy water (D2O), 3-(4,5-dimethylthiazole-2-yl)-2,5-diphenyltetrazolium bromide (MTT), propidium iodide (PI), and ribonuclease (RNase) were purchased from Sigma Co. (USA). Tris (D11, 98%) was purchased from Cambridge Isotope Laboratories, Inc. Analytical-grade inorganic salts were purchased from Sinopharm Chemical Reagent Beijing Co. Trypsinase was purchased from AMRESCD.
Cell line
Adenocarcinomic human alveolar basal epithelial cells (A549) were provided by the College of Life Sciences, Beijing Normal University. Cell lines were cultured in Dulbecco modified Eagle medium (DMEM; Sigma Chemical Co.) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Beijing YuanHeng ShengMa Biology Technology Research Institute, China) in the presence of 5% CO2 at 37°C.
Sample preparation
The preparation of DNA samples (Hum6 and Hum24) was according to Lu et al. (1993) and Phan and Patel (2003). The buffer solution for circular dichroism melting spectroscopy
Circular dichroism melting spectroscopy
CD spectra were collected using melting-CD at 200–350 nm on a Jasco-815 automatic recording spectropolarimeter with a 1-cm pathlength quartz cell at 25°C. Each spectrum was the average of two scans. A buffer blank correction was made for all spectra. The temperature of the cell holder was regulated by a JASCO PTC-423S temperature controller, and the cuvette-holding chamber was flushed with a constant stream of dry nitrogen gas to avoid water condensation on the cuvette exterior. Melting curves of the G4 were obtained by recording the CD intensity at 262 nm. The heating rate was 2.0°C min−1.
Variable temperature ultraviolet
Melting-UV studies were carried out on a Varian Cary 1E UV-visible spectrophotometer equipped with a Peltier temperature controller. The spectral absorbance was measured at 295 nm according to reference (Mergny et al., 1998) with a 1-cm pathlength quartz. The heating rate was 3.0°C min−1, and absorption data recorded were collected every 0.5 minute on melting steps.
1H NMR measurements
1H NMR measurements were performed on a Bruker AVANCE 600 spectrometer equipped with a 5-mm broadband inverse (BBI) probe capable of delivering z-field gradients and TOPSPIN software (Bruker, version 2.0). All experiments were carried out at 298.2 K. The 1D proton spectra were recorded by the standard pulse program p3919gp that applies 3-9-19 pulses with gradients for water suppression.
Molecular modeling simulations
The molecular modeling and simulation work was performed using the Insight II 2005 software package (San Diego, CA) on a Dell Precision T5400 workstation under CHARMM force field. The structures of Tet, Fan, and Ber were first built using the sketching module and then minimized for 200 steps by the steepest descent algorithm. The structures of the d(TTGGG(TTAGGG)3A) (H24) G4 were obtained from the Research Collaboratory for Structural Bioinformatics (RCSB) Protein Data Bank, and proper modification was performed. The binding sites of the receptors were from previous reports (Luu et al., 2006).
MTT assay
Exponentially growing A549 cells were plated in 96-well plates in a 5% CO2 incubator at 37°C. Graded concentrations of the investigational compounds were added to the cell plates after 24 hours. Each concentration had three repeats. Cell proliferation was determined 2 days after continuous exposure to the three alkaloids by addition of MTT. Proliferation was analyzed spectrophotometrically at 570-nm absorbance, and the concentration required to inhibit cell growth by 50% (IC50) and 10% (IC10) was calculated. The cell inhibition rate was calculated as the percentage of MTT as follows: Percentage of inhibition=1−(mean experimental absorbance/mean control absorbance)×100%.
Flow cytometric assay
The effects of Tet, Fan, and Ber on the cell cycle were assessed by flow cytometry (FCM) by measuring the percentage of cells in G1, S, and G2 phases, with and without treatment with the investigational compounds. The FCM evaluation of the cell cycle status and apoptosis was performed according to a method described previously (Wang et al., 2007). In brief, after 24 hours of cultivation, cells were treated with alkaloids (16 μM) for 2 days. The cells were then washed twice with phosphate-buffered saline (PBS), fixed overnight with cold 70% ethanol, and stained with a solution containing 50 mg/mL PI and 10 μg/mL RNase for 30 minutes. Samples were filtered through a 37-μm nylon mesh into 12-×75-mm tubes before measurement. All samples were analyzed with an EPICS Elite flow cytometer (Coulter Cytometry, Miami, FL) (Izbicka et al., 1999).
Results and Discussion
Melting-CD
Melting studies are widely used to determine the stability of nucleic acids and their interaction with ligands. Melting temperature (Tm) indicates the stability of the structure; the increase temperature in Tm (ΔT) is used to compare the heat-stabilizing ability of G4-ligands (Yin et al., 2003). All experimental alkaloids exhibited G4 stabilization ability with obvious and different ΔT values. Fan showed the highest ΔT, whereas Ber showed the lowest (Table 1). This may be the result of the slight difference in their side chains (Fig 2A): (1) Fan has an –OH at R1, whereas for Tet and Ber it is –OCH3, which has a bigger steric hindrance. Moreover, this –OH is on the potential binding plane. It can form a hydrogen bond with the –OH or –NH2 of G4. (2) The substituent group of Ber at R2 and R3 is different from that of Tet and Fan, which may reduce the planarity of Ber. As a result, Fan shows smaller steric hindrance than Tet and Ber (Fig 2B). It is tempting conclude that –OH at R1, –OCH3 at R2, and at R3 is crucial for displaying high stabilization ability.
Tet, tetrandrine; Fan, fangchinoline; Ber, berbamine; CD-melting, circular dichroism melting spectroscopy.
In addition, we observed Hum24-G4 to have a positive band at 286 nm and a smaller shoulder at 273 nm, which is more complex than the common ≈260 nm for parallel G4 and ≈290 nm for antiparallel G4. Intramolecular G4 like Hum24-G4 in K+ solution was identified to contain parallel, antiparallel, and a new hybrid-type structure (Wu and Bonner, 1981). However, the exact structure of intramolecular G4 in K+ solution is extremely controversial to date (Haq et al., 1996). Our experiment provided additional evidence for the intramolecular G4 structure diversity.
Moreover, it is worth noting that there is a dramatic and pronounced change with the addition of the alkaloids, especially with Tet. We observed a shift from the 286-nm band to the 267-nm band, which indicated the destruction of the antiparallel structure and the formation of a parallel structure (Nelson et al., 2002) [Fig 3; Supplementary Fig. 1 (Supplementary materials are available online at www.liebertpub.com)]. The 286-nm band almost disappeared at 63°C. We inferred that parallel G4 was more stable to high temperature, in other words, parallel conformation may be a preponderant structure in adverse situations. However, this conformation exchange was not observed with Ber.
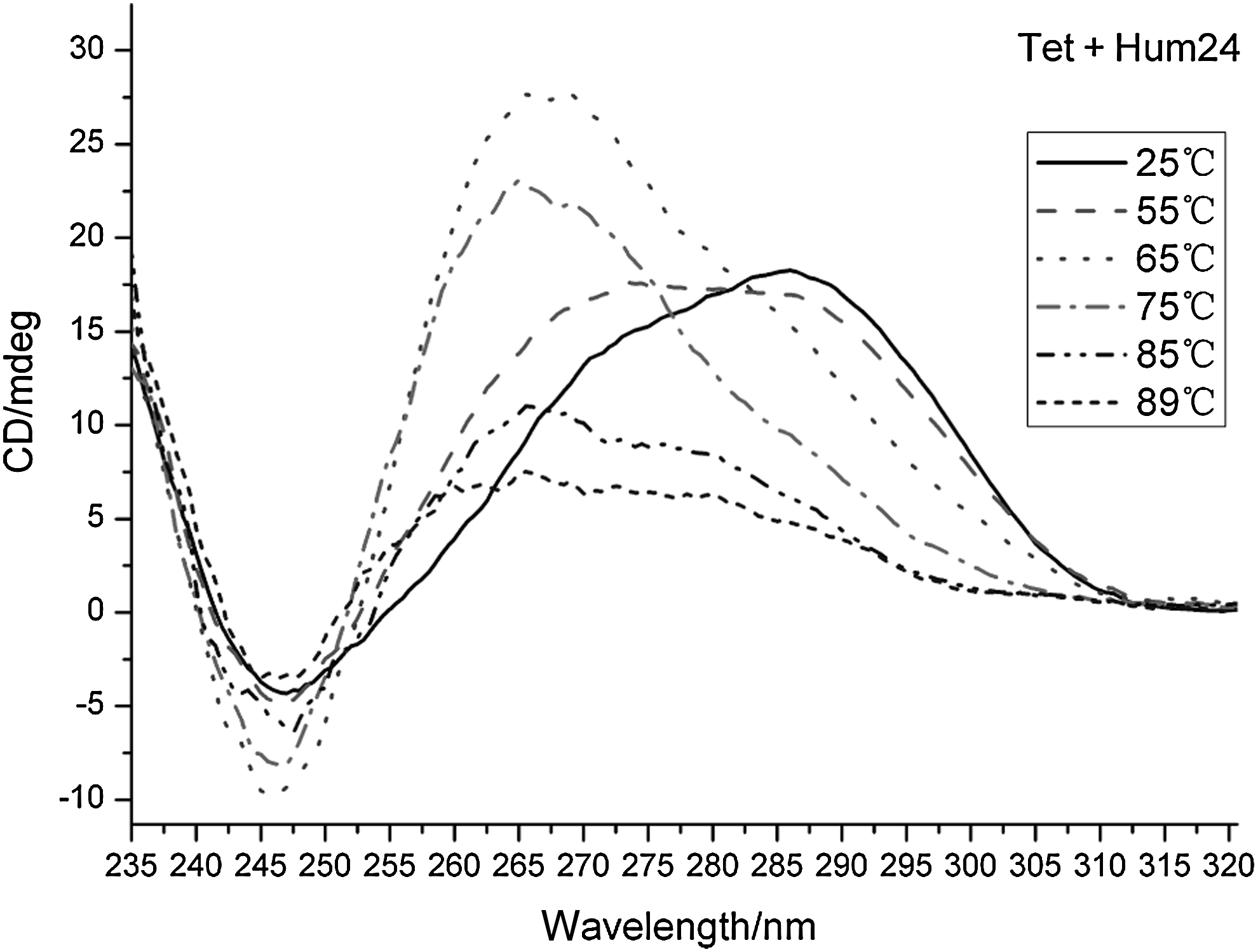
Circular dichroism (CD) spectrum of tetrandrine (Tet) with Hum24 in K+ solution. With the increase of the temperature, there is a shift from the 286-nm band to the 267-nm band.
Structural transition between antiparallel and parallel forms of G4 in telomeric DNA plays an important role in telomere formation (Nelson et al., 2002). Similar conformational conversion from antiparallel to parallel G4 occurred in the in vivo molecular crowded environment imitated by polyethylene glycol (PEG). When the concentration of PEG was large enough (40%), the antiparallel G4 totally disappeared. Parallel-stranded quadruplex was also reported to be unusually stable and greatly decreased telomerase processivity (Wu and Bonner, 1981). However, before the first report of the parallel-stranded crystal conformation by Neidle's group, the parallel-stranded structure had been found in nonhuman telomere G4, mostly in intermolecular form and only in multimeric intermolecular quadruplexes formed by human telomere DNA of less than four repeats (Fox et al., 1995). Our results support the view that parallel-stranded structure can exist in intramolecular G4. Except for the 24-nucleotide Hum24, G4 conformation exchange induced by some small molecular is reported in the 22-nucleotide human telomeric sequence d[AGGG(TTAGGG)3] and c-MYC G4 (Haq et al., 1996). More attention should be paid to parallel G4.
Melting-UV
UV absorbance is the most commonly used technique for determining Tm. It is a qualitative but rapid and convenient initial method to identify promising G4 ligands (Yin et al., 2003). We wondered whether R2 and R3 affected the stability so much, so we examined the Tm of Hum6 and Hum24 with or without Tet, Ber, and Neferine (Nef, Fig. 2). Nef has a structure similar to Tet and Ber, and it has the same substituent groups at R2 and R3 as Ber (Cao et al., 2004). Tet shows the same ΔT to both Hum6 and Hum24, and has a higher ΔT than Ber and Nef (Supplementary Table 1). Nef and Ber have the same structure at R2 and R3, and both of them show weak stability to Hum6 and Hum24. Although Nef is an open-loop molecule and Ber is not, this result supports the conclusion that R2 and R3 are two key sites that determine stabilization.
1H NMR
Except for stabilization ability, we expected the alkaloids to induce the formation of G4. NMR is an essential tool in the study of quadruplex nucleic acids formation. Chemical shifts at 10–12 ppm for guanine imino protons in 1H NMR are characteristic of a G4 structure (Phan et al., 2006). Because of the low proton density of nucleic acids, the bases forming the G4 can be quickly recognized. So we applied 1H NMR to study the G4-inducing ability of the alkaloids. Under salt deficiency conditions, Hum24 is in the form of a random single-strand motif, and there is no guanine imino proton signal in the 10- to 12-ppm region (Fig. 4). When we added alkaloids at the ratio of alkaloid/Hum24=1:8, we observed some obvious imino proton signals, which are unique features of the G4 structure, suggesting the formation of G4. Because the three alkaloids have a similar structure, it is reasonable to think of them inducing a same type of G4. We can see that the signals induced by the three alkaloids were almost the same with some tiny differences (Fig. 4). The proton resonances imply that there may be exchanges from one G4 type to another, consistent with results from CD-melting.
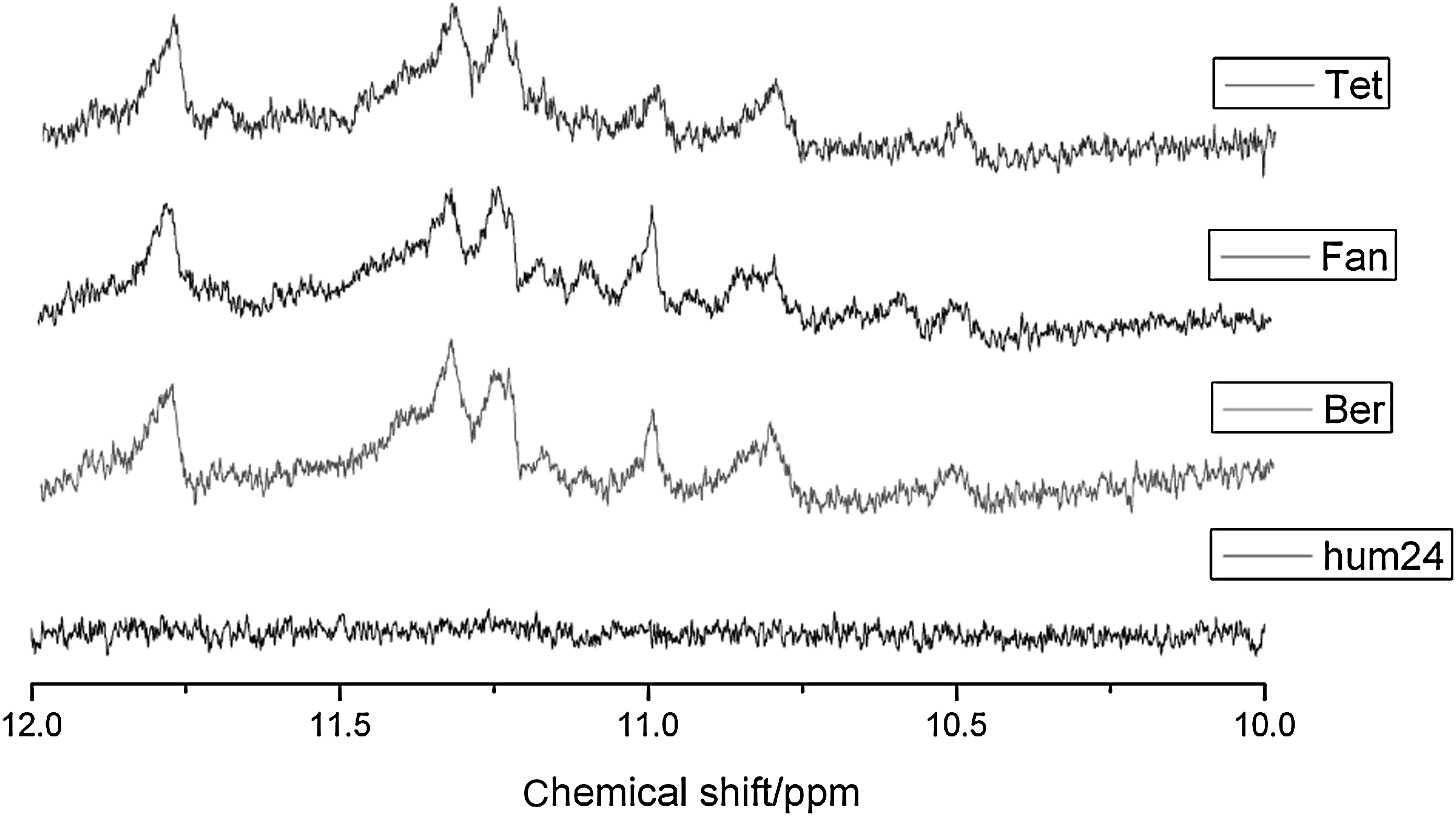
Proton nuclear magnetic resonance (1H NMR) [600MHz, 10% heavy water (D2O)] spectra (10–12ppm) of Hum24 G4 with tetrandrine (Tet), fangchinoline (Fan), and berbamine (Ber).
Molecular docking
To confirm the action patterns of Tet, Fan, and Ber with G4, we applied computer simulation. The receptor model we chose was G4 formed by the d[TTGGG (TTAGGG)3A] (H24) sequence. According to the external character of telomeric G4, it can be divided into three planes, three loops, and a site (Supplementary Fig. 2) (Luu et al., 2006). Among the planes, Plane 1 and Plane 3 interact more frequently with small molecules. The results showed that Tet, Fan, and Ber tended to bond Plane 3, Plane 1, and Plane 3, respectively (Supplementary Table 2). Their highest scores were inconsistent with their stabilization ability, but the scores of Plane 1 fit their stabilization ability well (scores for Tet, Fan and Ber were 6.15, 6.83, and 5.76, respectively). We inferred that in solution conditions, G4-ligands can bind to Plane 1 more easily because it is exposed more. Moreover, conformation exchange in G4 could occur in the solution conditions similar to d[(GGGTTA)3GGG] (Supplementary Fig. 3A) that would not be considered in molecular docking.
From the results of spectrum, chromatograph, and computer simulation, we could determine that Tet, Fan, and Ber can increase the stability and induce the formation of G4. However, the in vivo environment is known to be complex and G4 DNA is a dynamic structure, making it a particular challenge to establish its biological functions. We applied MTT to examine the three alkaloids' cytotoxicity and FCM to determine their effects on the cell cycle.
MTT assay
It has been hypothesized that if the cytotoxicity of the alkaloids accords with their stabilization ability, the potential toxicity and growth inhibition may be the result of the formation of G4. So we assessed the IC50 of Tet, Fan, and Ber on A549 cells by using the MTT assay. All three alkaloids turned out to have moderate cytotoxicity to A549 cells in a dose-dependent manner, with Fan showing the highest cytotoxicity (IC50=21.6 μM). However, Ber showed a higher cytotoxicity than Tet (Supplementary Table 3), discordant with their stability. One attraction is that Tet and Ber can bind to double-stranded DNA (dsDNA) other than G4. The former binds to dsDNA via pure stacking like ethidium bromide, whereas the latter binds to dsDNA similar the traditional minor-groove binders, involves hydrogen binding, and shows a higher affinity (Chen et al., 2011). In addition, Ber is also reported to be a DNA polymerase and reverse transcriptase inhibitor (Efferth and Wink, 2010), which may explain the inconsistency of the growth inhibition ability with the G4 stabilization ability.
FCM assay
It has been reported that G4 ligands can not kill tumor cells, but instead induce replicative senescence after a long-term incubation (Cameron and Greulich, 1963). Moreover, telomerase activity changes as cells progress through the cell cycle in tumor cell lines (Fig. 5) (Zhu et al., 1996; Izbicka et al., 1999), with the activity downregulation accompanied by cell cycle arrest (Yin et al., 2004). It is necessary to study the influence of alkaloids on the cell cycle. FCM results indicated that the alkaloids can terminate the cell cycle of A549 cells in G1 (Fig. 6). This is similar to triethylene tetraamine (TETA), which is reported to stabilize both inter- and intramolecular G4 structures, inhibit telomerase in vitro (Yin et al., 2003), suppress cell proliferation, and arrest HeLa cells in G1 (Yin et al., 2004). The alkaloids we studied may function in the same way as TETA. Furthermore, no apoptosis was observed. It is demonstrated that the repression of telomerase activity could also be either a specific component of the differentiation program, or simply a consequence of the same mechanism that downregulates the enzyme in quiescent cells (Bestilny et al., 1996; Yin et al., 2004). We speculated that the three alkaloids induce differentiation or quiescence in these cells by interacting with G4, which in turn results in the repression of telomerase instead of killing cells.
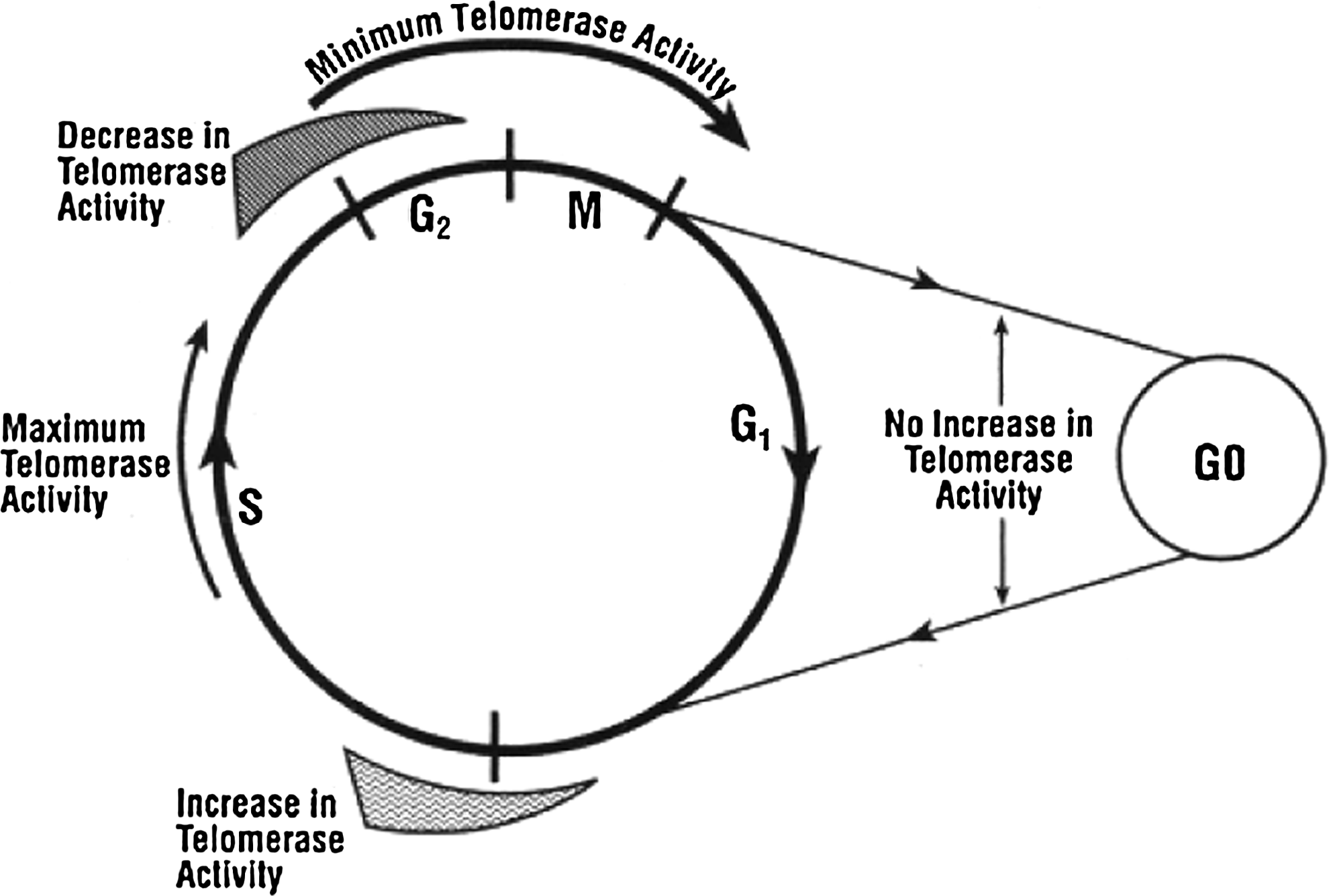
Proposed model for cell-cycle regulation of telomerase (Zu et al., 1996).
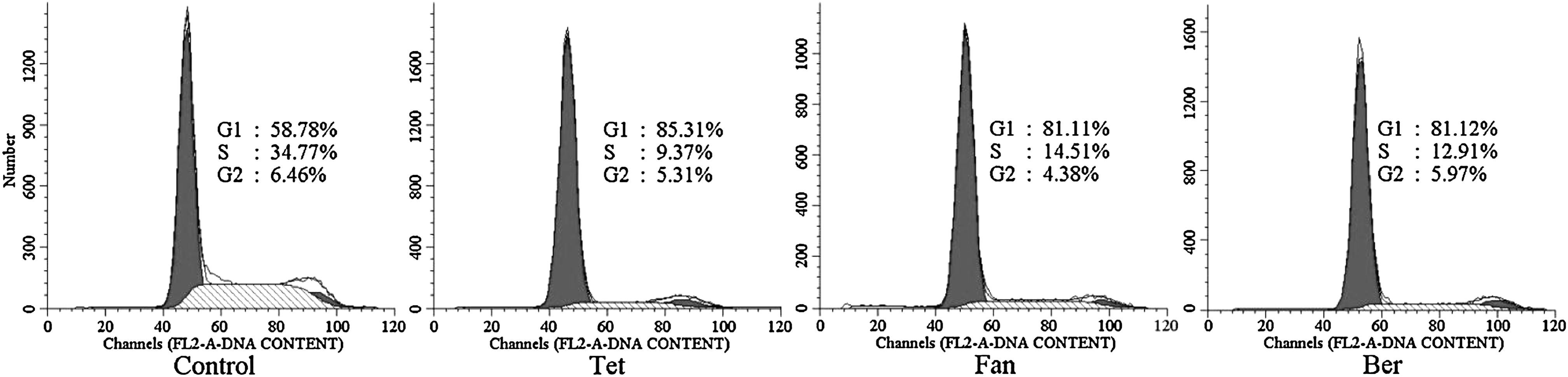
Results of flow cytometry (FCM). Cells in the control group were not treated with drugs. Cells in the experimental group were treated with tetrandrine (Tet), fanchinoline (Fan), and berbamine (Ber), respectively.
Conclusions
In the present study, Tet, Fan, and Ber were shown to increase the stability and induce the formation of G4. Tm increased as much as 12°C with Fan. This added structural diversity to G4-ligands, because Tet, Fan, and Ber possess novel structures with a rigid cyclophane and a rough equilateral triangle shape, which is different from the characteristic G4 ligands (Han and Hurley, 2000; Yin et al., 2003; Liu et al., 2010). –OH at R1, –OCH3 at R2, and at R3 seemed to be important for displaying high stabilization ability. Besides, the three alkaloids showed moderate cytotoxicity and turned out to be cell cycle blockers. Therefore, Tet, Fan, and Ber have been identified as potent telomerase inhibitors. However, further work is needed to perfect this study.
First of all, we found that different structures of G4 co-exist in the K+ solution, and conformation exchange happens in the presence of Tet and Fan with the increase of the temperature. It is noteworthy that different structural forms may be important for participation in different biological functions of telomeres. For example, the unique topology of the mixed parallel/antiparallel structure is reported to facilitate stacking of the G4 units into higher-order packing structures, such as the t-loop (Supplementary Fig. 3) (Xu et al., 2006), which can provide flexibility for responding to environmental conditions as protein binding. These structures might also be important for effectively packing telomeric DNA into active or inactive forms upon chromosome condensation (Haq et al., 1996; Nelson et al., 2002). Moreover, parallel G4 seems to take advantage of antiparallel G4 in an adverse and molecularly crowded environment, which provides some clues regarding cell actions under these conditions. Determining the exact conformation of human telomeric G4 and its specific function is important not only for a better understanding of the biological implications, but also for successful drug design targeting the human telomere quadruplex.
Second, we speculated that Tet, Fan, and Ber can arrest cell cycle by inducing cell differentiation or quiescence. Cell cycle regulation is very complicated, as is the inhibition of telomerase activity, and numerous regulatory proteins are involved in these processes. For example, transforming growth factor-β (TGF-β) (Koff et al., 1993), cyclin-dependent kinases (CDKs) (Redon et al., 2003), and CDK inhibitors such as p21 (Yin et al., 2004) are expressed and/or activated during the G1 phase and can inhibit telomerase activity. In fact, G4 functions through interacting with proteins in many cases (Bates et al., 1999; Dapic et al., 2003). Ligands may interact with G4, which further affects the binding of the G-overhang to telomerase directly, or proteins that will in turn affect telomerase activity. Although the mechanism is unclear, there can be a close relationship between telomeres, telomere-associated proteins, and telomerase itself (Chen et al., 2011).
Third, although a number of compounds that recognize G4 DNAs have been developed, most ligands have broad specificity. Because many genomic regions have the potential to form G4 DNA, off-target effects might be a serious concern (MAIZELS, 2006). Therefore, the specificity should be a concern for finding therapeutically useful ligands.
Fourth, the formation of G4 can not only occur in the telomere G-overhang, but also potentially in G/C-rich dsDNA. It is known that duplex DNA (dsDNA) opens in many biological DNA-processing events, such as replication, transcription, and promoter recognition (Smith and Martin, 1973), exposing the single-stranded DNA in the GC-rich region of a promoter and allowing G4 and i-motif structures to form. Moreover, it has been shown that molecular crowding plays a bifacial role in competition by stabilizing quadruplex and, in the meantime, destabilizing duplex structures. Small molecules that stabilize quadruplex or/and destabilize duplex structures may potentially induce such quadruplex formation, thus offering the possibility of manipulating the structure of double-stranded telomere DNA (Supplementary Fig. 4) (Smith and Martin, 1973; Miyoshi et al., 2004). However, although the work on the promoter of the c-myc gene is consistent with quadruplex formation, the formation of G4 in dsDNA is still unclear. We know the amount of dsDNA is much more than G-overhang in vivo, providing more drug-binding sites. It is of significance to carry out studies on G4 form in dsDNA.
Footnotes
Acknowledgments
The authors are greatly thankful for the help of Professor Mu Pu (College of Life Science, Beijing Normal University) in improving language in this manuscript. This work was support by the Chinese National Natural Science Foundation (No. 30973869) and National Natural Science Foundation of China (Grant No. 81072576).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.