
Abstract
Human DNA-polymerase iota (Pol ι) is an extremely error-prone enzyme and the fidelity depends on the sequence context of the template. Using the in vitro systematic evolution of ligands by exponential enrichment (SELEX) procedure, we obtained an oligoribonucleotide with a high affinity to human Pol ι, named aptamer IKL5. We determined its dissociation constant with homogenous preparation of Pol ι and predicted its putative secondary structure. The aptamer IKL5 specifically inhibits DNA-polymerase activity of the purified enzyme Pol ι, but did not inhibit the DNA-polymerase activities of human DNA polymerases beta and kappa. IKL5 suppressed the error-prone DNA-polymerase activity of Pol ι also in cellular extracts of the tumor cell line SKOV-3. The aptamer IKL5 is useful for studies of the biological role of Pol ι and as a potential drug to suppress the increase of the activity of this enzyme in malignant cells.
Introduction
The biological role of Pol ι in the cells is not precisely defined (Ohkumo et al., 2006; Choi et al., 2006). The enzyme was implicated in both the prevention and the promotion of carcinogenesis (Stallons and McGregor, 2010). For example, the experiments with cultured breast cancer cells have shown a correlation between a high level of Pol ι expression and enhanced mutagenesis in these cells (Yang et al., 2004). We have previously found that cell extracts of human basal-cell carcinoma and melanoma possess an elevated ability for DNA synthesis after wrong nucleotide misincorporation, which can be attributed to the high activity of Pol ι (Gening et al., 2006a; Kazakov et al., 2010). It was also suggested that Pol ι participates in the protection of cells from oxidative stress and UV irradiation (Dumstorf et al., 2006; Ohkumo et al., 2006; Petta et al., 2008). One of the approaches to further study the role of Pol ι is the use of aptamers, which would specifically inhibit the DNA-polymerase activity of this enzyme. The aptamers possess high specificity and affinity, but contrary to the monoclonal antibodies, are not recognized by the immune system and produce no immune response. For this reason, the aptamers are promising tools for studies of protein mechanisms and roles as well as for new-generation diagnostics and therapy (Nimjee et al., 2005; Ulrich et al., 2006; Yan et al., 2005).
The aim of this work was to develop and characterize the aptamers for the human Pol ι capable of providing a specific inhibitory effect on the DNA polymerase activity of this enzyme. Using the systematic evolution of ligands by exponential enrichment (SELEX) procedure (Ellington and Szostak, 1990; Marshall and Ellington, 2000), we obtained the aptamer IKL5, capable of inhibiting DNA Pol ι polymerase activity in both homogenous preparations and in extracts of the human ovarian cancer SKOV-3 cells. Under our experimental conditions, the aptamer did not inhibit the other DNA polymerases present in the extracts.
Materials and methods
Production of recombinant proteins
We used the catalytic core of the human Pol ι, 420 amino acids long, fused with glutathione-S-transferase (GST) at the N end. The enzyme was produced in yeast using the expression yeast vector obtained and purified as described earlier (Makarova et al., 2011). The purified human Pol β and Pol κ preparations used in this work were produced and isolated from Escherichia coli (Gening et al., 2006b). All enzymes were >95% pure.
Oligodeoxynucleotide library and synthesis of the initial oligoribonucleotide pool
At the first step we synthesized a library of single-chain oligodeoxyribonucleotides as described by Gening et al. (2006b). An aliquot used in the first round of selection contained about 8×1012 variants of a 30-nucleotide sequence flanked with constant regions. One constant region contained a promoter sequence for phage T7 RNA polymerase. The initial pool of oligoribonucleotides was produced by in vitro transcription with T7 RNA polymerase overnight on a template of the PCR product obtained by amplification of the single-chain DNA library. The oligonucleotides in the library had the following structure: 5′-GGGAATGGATCCACATCTACGAATTC—30N—TTCACTGCAGACTTGACGAAGCTT-3′, where 30N is a variable 30-nucleotide site. Primer sequences for PCR are 5′-GA
SELEX procedure
To select the aptamers for Pol ι from the oligoribonucleotide pool, we ran 10 rounds of SELEX (Conrad et al., 1996). The SELEX procedure includes 2 repeating processes: separation of nucleotides with higher affinity toward the target molecule from other nucleotides by affinity binding and amplification of the selected oligoribonucleotides pool. In the first round of SELEX, Pol ι in a concentration of 20 nM was incubated with the 3 μM oligoribonucleotides in 500 μL of the binding buffer. The concentrations of Pol ι and of the oligoribonucleotide pool was then changed (Table 1). For the first 5 rounds, the pool was enriched by filtration through the nitrocellulose membrane. After each filtration, the membrane containing the RNA-protein complex was incubated in the rinsing buffer No. 1 (7 M urea, 20 mM Tris-HCl, pH 7.7, 120 mM NaCl, 1 mM MgCl2, 5 mM KCl) for 10 minutes at 50°C. Then, the oligoribonucleotides dissociated from the RNA–protein complex were precipitated with cold ethanol and used for reverse transcription, amplification of synthesized cDNA, and RNA transcription in the next round of SELEX (Conrad et al., 1996).
Pol ι, Human DNA-polymerase iota.
To exclude the enrichment of the pool with oligoribonucleotides that specifically interact with nitrocellulose, after round 5 the selection was continued using Pol ι immobilized on glutathione sepharose. Immobilization of Pol ι on the sorbent was done in the binding buffer at room temperature for 40 minutes. After subsequent 30 minutes incubation with the oligoribonucleotide pool at room temperature, the oligoribonucleotides not bound with Pol ι were removed by rinsing buffer No. 2 (20 mM Tris-HCl, pH 7.7, 120 mM NaCl, 1 mM MgCl2, 5 mM KCl, 0.2% Tween 20). The rinsing buffer No. 1 was added to glutathione sepharose containing the RNA–protein complex, and the mixture was incubated for 10 minutes at 50°C. Then, the eluted oligoribonucleotides were precipitated with cold ethanol and used for the next round of SELEX as described previously.
The RNA obtained after round 9 of the SELEX was amplified by real-time PCR (RT-PCR). The synthesized cDNAs were ligated with vector pGEM-T (Promega). The lysates of E.coli transformants with the obtained vector were used for PCR. These PCR products were used as templates for aptamer synthesis (Conrad et al., 1996; Marshall and Ellington, 2000).
Evaluation of the affinity of selected oligoribonucleotides
In the course of the SELEX procedure, in order to control the increase in affinity of the oligoribonucleotide pools for Pol ι, we evaluated the affinity after rounds 2, 4, 6, 8, 9, and 10. A radiolabeled oligoribonucleotide pool was synthesized in the reaction mixture containing 40 mM Tris-HCl, pH 7.9, 25 mM MgCl2, 30 mM DTT, 0.2 mM each rNTP, 50 pM [α-32P]UTP, 20 units T7 RNA polymerase, and 0.2 picomoles of the PCR product in a final volume of 20 μL. After 1 hour of incubation at 37°C, the synthesized labeled oligoribonucleotide pool was added to the binding buffer containing a series of dilutions of Pol ι. After 30 minutes of incubation at room temperature, the mixture was filtered through a nitrocellulose filter. Affinity of an oligoribonucleotide pool toward Pol ι was determined by the dependence of filter radioactivity on the enzyme concentration.
Analysis of SSCP for individual RNA aptamers
In order to study a higher number of aptamers and to exclude identical aptamers, we performed a single-strand conformation polymorphism analysis (RNA-SSCP) of the nonradioactive oligoribonucleotides selected by the SELEX procedure. For this purpose, individual RNA aptamers were mixed with one-half volume of 50% sucrose containing 0.001% bromphenol blue. Then, the samples were analyzed by electrophoresis in 8% nondenaturing PAAG at 8 mA current at 4°C. Next, the gel was either stained with ethidium bromide and imaged with UV transilluminator or stained with SybrGreen II and analyzed using Phosphoimager Storm 840 (Amersham Biosciences Corp.).
Preparation of aptamers for in vitro analysis
Individual RNA aptamers were synthesized in vitro by transcription of the PCR products obtained on the lysates of individual E. coli clones (see above). The in vitro transcription reaction was run in the mixture containing 40 mM Tris-HCl, pH 7.9, 25 mM MgCl2, 30 mM DTT, 2 mM each of the four rNTPs, 200 units T7 RNA polymerase, and 5 picomoles of the PCR product in the final volume of 200 μL. The mixture was incubated overnight at 37°C. Next, the synthesized RNA aptamers were purified by denaturing electrophoresis in PAAG (Conrad et al., 1996). After elution from the gel and precipitation with ethanol, the aptamers were dissolved in 100 mM NaCl. For aptamer refolding, the mixture was incubated for 3 minutes at 73°C and gradually cooled down to room temperature. The concentration of RNA aptamers was measured using spectrophotometer NanoDrop 1000 (Thermo Fisher Scientific).
Determination of Kd for the RNA aptamers
Individual radiolabeled RNA aptamers were synthesized as described previously for oligoribonucleotide pools produced for affinity evaluation. Labeled aptamers were incubated in the binding buffer with a series of dilutions of Pol ι for 30 minutes at 25°C. Then, the mixtures passed through 2 filters: the upper nitrocellulose filter retained RNA within the complex with Pol ι, while the lower diethylaminoethyl-cellulose filter (DE-filter) retained the aptamers unbound with the protein. Radioactivity of the filters after filtration of the samples with different concentrations of the protein and the same concentration of the aptamer was quantified on Phosphoimager Storm 840 with ImageQuant program. The data were used to calculate the dissociation constants for each aptamer with GraFit (Erithacus Software), using the equation B=A C/(Kd + C), where B is an amount of the bound aptamer at a given concentration of Pol ι; A, a maximal amount of the bound aptamer; C, a concentration of the aptamer; and Kd, a dissociation constant.
The substrate to test DNA-polymerase activity of Pol ι
The substrates to test the Pol ι activity were complementary single-stranded oligoribonucleotides containing 17 (primer) and 30 (template) nucleotides [5′-GGAAGAAGAAGTATGTT-3′ and 5′-CCTTCTTCATTCTAACATACTTCTTCTTCC-3′]; they hybridize to form a duplex with a protruding 5′ end (Zhang et al., 2000). The primer was 5′-labeled using T4 polynucleotide kinase (10 units) and 2 MBq of [γ-32P]ATP in 70 mM Tris-HCl, pH 7.6, 5 mM DTT and 10 mM MgCl2. Radiolabeled DNA substrates were prepared in 200 μL by annealing (at 73°C for 3 minutes with gradual cooling down to room temperature) 10 pmol of labeled primer with 15 pmol of the template in the polynucleotide kinase buffer with NaCl concentration adjusted to 100 mM.
Testing of aptamer effect on DNA-polymerase activity in homogenous preparations of Pol ι, Pol β, and Pol κ
Aiming to analyze the inhibitory effect of the obtained aptamers on DNA polymerases, we measured the DNA-polymerase activity of these enzymes in the presence of the aptamers. RNA aptamers were preincubated with DNA polymerases for 10 minutes at 25°C in the mixture containing 50 mM Tris-HCl, pH 8.0, 100 μM of each dNTP, and either 1 mM MgCl2, or 200 μM MnCl2 (with Pol ι), or 5 mM MgCl2 (for Pol β and Pol κ). Then the radiolabeled substrate was added to a concentration of 5 nM, and the mixture was incubated for 20 minutes at 37°C for the reaction with Mg-containing buffer, or 10 minutes at 37°C for the reaction with Mn-containing buffer. Next, the samples were placed on ice, and an equal volume of the loading buffer (95% formamide, 0.1% bromphenol blue, 0.1% xylene cyanol, 10 mM EDTA) was added to each tube. The reaction products were separated in 18% denaturing PAAG and visualized by autoradiography with Phosphoimager Storm 840. The obtained data were evaluated using the programs ImageQuant and GraFit.
Estimation of the aptamer concentration at which the DNA-polymerase activity of Pol ι is 50% of the activity in the absence of the aptamer
Aiming to study the inhibitory activity of the aptamer IKL5 toward Pol ι we ran a DNA-polymerase reaction catalyzed with homogenous preparation of Pol ι at various concentrations of the aptamer. The half maximal inhibitory concentration (IC50; the aptamer concentration at which the DNA-polymerase activity of Pol ι is 50% of the activity in the absence of the aptamer) was estimated using the program GraFit, from the equation B=
Preparation of extracts from tumor cells SKOV-3
Human ovarian cancer cell line SKOV-3 was kindly provided by Dr. Galina A. Posypanova (Moscow Research Institute of Medical Ecology, Russia). The cell line was cultured in RPMI 1640 containing 10% fetal bovine serum (PAA Laboratories) and 50 μg/mL gentamicin at 37°C in a humidified incubator, with 95% air and 5% CO2.
For extract preparation, the cells were homogenized on ice with a Teflon homogenizer in 0.14 M PBS, pH 7.4 (Helicon). The buffer volume corresponded to 1 μL of buffer per 1 mg of homogenized culture. The obtained homogenate was centrifuged at 4°C for 10 minutes at 14,000 g. The supernatant was used for enzymatic assays. Protein concentration was measured using the Protein Assay Reagent Kit (BioRad) and adjusted to 5 mg protein/mL.
Testing of the inhibitory effect of the aptamer IKL5 on the DNA-polymerase activity of Pol ι in SKOV-3 cell extracts
The activity of Pol ι in SKOV-3 cell extracts was measured using the method of misincorporation of “G, versus“ A, method of gening (misGvA) developed by us earlier (Kazakov et al., 2010; Makarova et al., 2011). This method allows for testing of misincorporation of this enzyme in the presence of other DNA polymerases (e.g., in cell extracts). The method is based on differences in the electrophoretic mobility of primer elongation products generated by Pol ι and other DNA polymerases (containing incorrect nucleotides inserted vs. correct nucleotides, respectively).
To test the inhibitory effect of aptamers on DNA-polymerase activity of the cell extracts, the RNA-aptamer was preincubated with 3 μL cell extract for 10 minutes at 25°C in 15 μL of the mixture containing 50 mM Tris-HCl, pH 8.0, 1 unit RNase inhibitor (Fermentas), 200 μM MnCl2, and 0.5 mM each of dATP and dGTP. Then radiolabeled substrate was added to 5-nM concentration and the mixture was incubated for 20 minutes at 37°C. Next, the samples were placed on ice, and an equal volume of the loading buffer was added to each tube. The reaction products were resolved and analyzed as described previously for pure enzymes.
Relative misincorporation of G (MoG) by Pol ι was calculated with the following formula: MoG=[(d + c + b + a)×100%)/ (4(D + d) + 3(C + c) + 2(B + b) + (A + a)], where A, B, C, and D are intensities of electrophoretic bands +1 (18A), +2 (19G), +3 (20A), and +4 (21A) representing the correct extension products generated by the accurate DNA polymerases; while a, b, c, and d are intensities of electrophoretic bands +1*(18G) and +2*, +3*, and +4* representing oligonucleotides with altered mobility resulted from the formation or extension of +1* product attributed to Pol ι.
Results
In vitro selection of RNA aptamers for Pol ι
RNA aptamers binding to human Pol ι and inhibiting DNA-polymerase activity of this enzyme were selected using the initial pool of oligoribonucleotides obtained as described in Materials and Methods. The final enriched oligoribonucleotide pool was obtained after 5 rounds of the SELEX procedure on nitrocellulose filters with 5 subsequent rounds on glutathione sepharose. Minimal Pol ι concentration allowing for the detection of binding to radiolabeled oligoribonucleotides decreased from round to round, down to round 9. Since the affinity of the oligoribonucleotide pool after round 10 did not differ from the affinity after round 9, we cloned the cDNA pool from the products of round 9.
Diversity of selected individual aptamers, as estimated by RNA-SSCP analysis
To avoid repeated detection of the same aptamers, we did the SSCP analysis for the oligoribonucleotides from 30 different clones of E. coli. We chose to do the RNA-SSCP instead of DNA-SSCP, because RNA-SSCP provides higher accuracy and is more informative. It gives higher chances of detecting oligoribonucleotides differing by even 1 nucleotide. In the course of RNA-SSCP analysis, various conformers of oligonucleotides were registered by electrophoresis in non-denaturing PAAG (Sarkar et al., 1992). As shown by RNA-SSCP, the conformers of all of the studied aptamers were identical, but distinct from the initial oligoribonucleotide pool and from aptamer 32 toward Pol β (data not shown). This suggests that there is no diversity of the oligoribonucleotide pool selected at round 9 of the SELEX.
Primary and secondary structure of the aptamers
Consistent with the results of the RNA-SSCP analysis, nucleotide sequences of 5 individual aptamers toward Pol ι selected at round 9 of the SELEX procedure were identical (Fig. 1A). An aptamer from the clone 5, named IKL5, was used for further studies. It should be noted that the fragment of aptamer IKL5 corresponding to the variable region consists not of 30, but of 29 nucleotides. It is possible that the shortening of the aptamer has a structural significance (e.g., stabilization of the structure of IKL5 and/or increase of affinity to the target protein). The secondary structure was evaluated for the full-size 79-base nucleotide sequence of the aptamer IKL5. The optimal secondary structure of the aptamer was predicted using the software RNAstructure; it is most stable and consequently most probable (Fig. 1B).
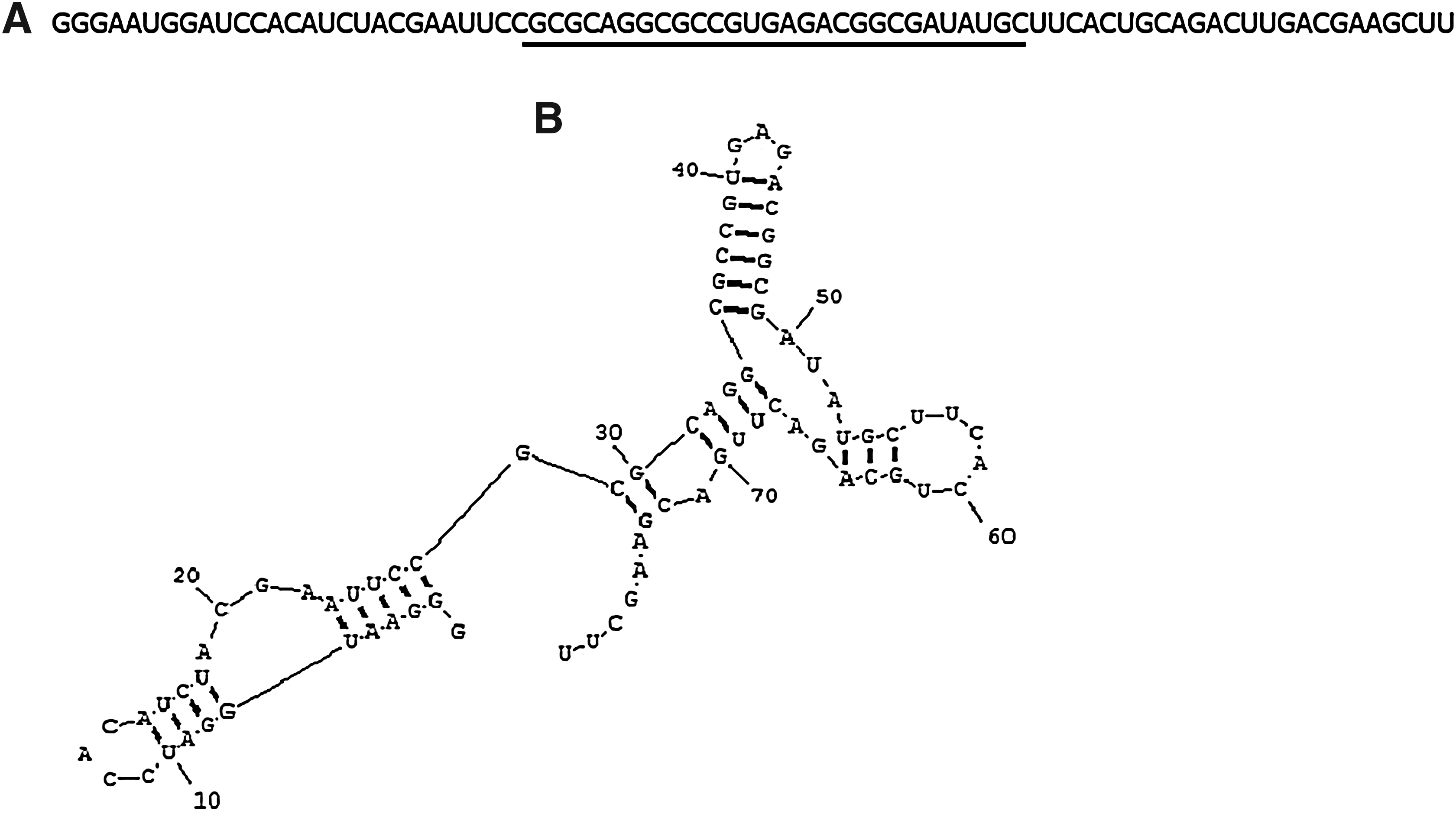
Structure of aptamer IKL5.
Affinity of the aptamer IKL5 for Pol ι and specificity of its binding
To evaluate the affinity of the aptamer IKL5 for Pol ι and to estimate the dissociation constant, we ran experiments to study the binding of radiolabeled IKL5 with different concentrations of Pol ι (see Material and Methods). The dissociation constant was found to be 4.2±1.4 nM.
The RNA aptamers that we selected earlier toward Pol β have relatively low specificity and inhibit polymerase activity of not only Pol β, but also Pol κ (Gening et al., 2006b). This suggests a possibility that these aptamers will inhibit DNA-polymerase activity of other error-prone DNA polymerases, probably because of some structural similarity of their catalytic centers. The RNA-aptamer 32 had the highest inhibitory activity and we used it to study the effects on Pol ι. To examine if it can displace IKL5 from the complex with Pol ι, we ran an experiment on competitive binding of this aptamer with Pol ι on nitrocellulose filters. Radiolabeled aptamer IKL5 was displaced from the complex with Pol ι only by the same unlabelled IKL5, while neither an excess of aptamer 32 nor an initial oligoribonucleotide pool had no effect on the stability of the complex IKL5-Pol ι (Fig. 2). It should be noted that the addition of the unlabeled aptamer in excess results in the saturation of the DE-filter by the aptamer with lower specific radioactivity. For this reason, sample 5 had lower level radioactivity than sample 1. The obtained results confirm that aptamer IKL5 binds to Pol ι specifically.

Competitive binding of radiolabeled aptamer IKL5 with Pol ι. The labeled aptamer was incubated with 10-nM Pol i in the presence or in the absence of 500-nM unlabeled aptamer 32, unlabeled initial oligoribonucleotide pool, or unlabeled aptamer IKL5. The samples were passed through 2 filters: the upper nitrocellulose filter (NT) retained RNA within a complex with Pol, and the lower diethylaminoathyl-cellulose filter (DE-filter) retained ribonucleotides unbound with the protein.
Inhibition of Pol ι DNA-polymerase activity by aptamer IKL5
Next, we analyzed the inhibition of the DNA-polymerase activity of human Pol ι by aptamer IKL5 in the presence of 1 mM Mg2+. The initial oligoribonucleotide pool, aptamer 32, and NaCl were added to the reaction mixtures instead of the aptamer IKL5 served as negative controls. In the lanes with Pol ι there was a doublet band of the 18-nucleotide product (Fig. 3A). The upper, 18G band corresponds to the product with misincorporated G opposite the template T, and the lower band, 18A, corresponds to the product with correctly incorporated A at the 3′ end (Kazakov et al., 2010; Makarova et al., 2011). There is a considerable, 85% suppression of DNA-polymerase activity of Pol ι in the presence of aptamer IKL5 (Fig. 3A, C). At the same time, the initial oligoribonucleotide pool or aptamer 32 had no effect on the DNA-polymerase activity of Pol ι (Fig. 3A, C). DNA-polymerase activity of Pol ι is likely resistant to aptamer 32 because of unique features of the organization of its active site, different from the other Y-family DNA polymerases (Kirouac and Ling, 2009; Pence et al., 2009).

Inhibition of DNA-polymerase activity of Pol i by aptamer IKL5.
A more detailed time-course analysis of the polymerases reaction confirmed the inhibitory effect of IKL5 (Fig. 3B, D).
Effect of Mg2+ and Mn2+ ions on the inhibition of DNA-polymerase activity of Pol ι by the aptamer IKL5
It is generally assumed that Mg2+ is the physiological cofactor for replicative DNA polymerases in vivo. In the presence of Mg2+ the value of IC50 for inhibition of Pol ι by aptamer IKL5 was 49±16 nM (Fig. 4A, B).
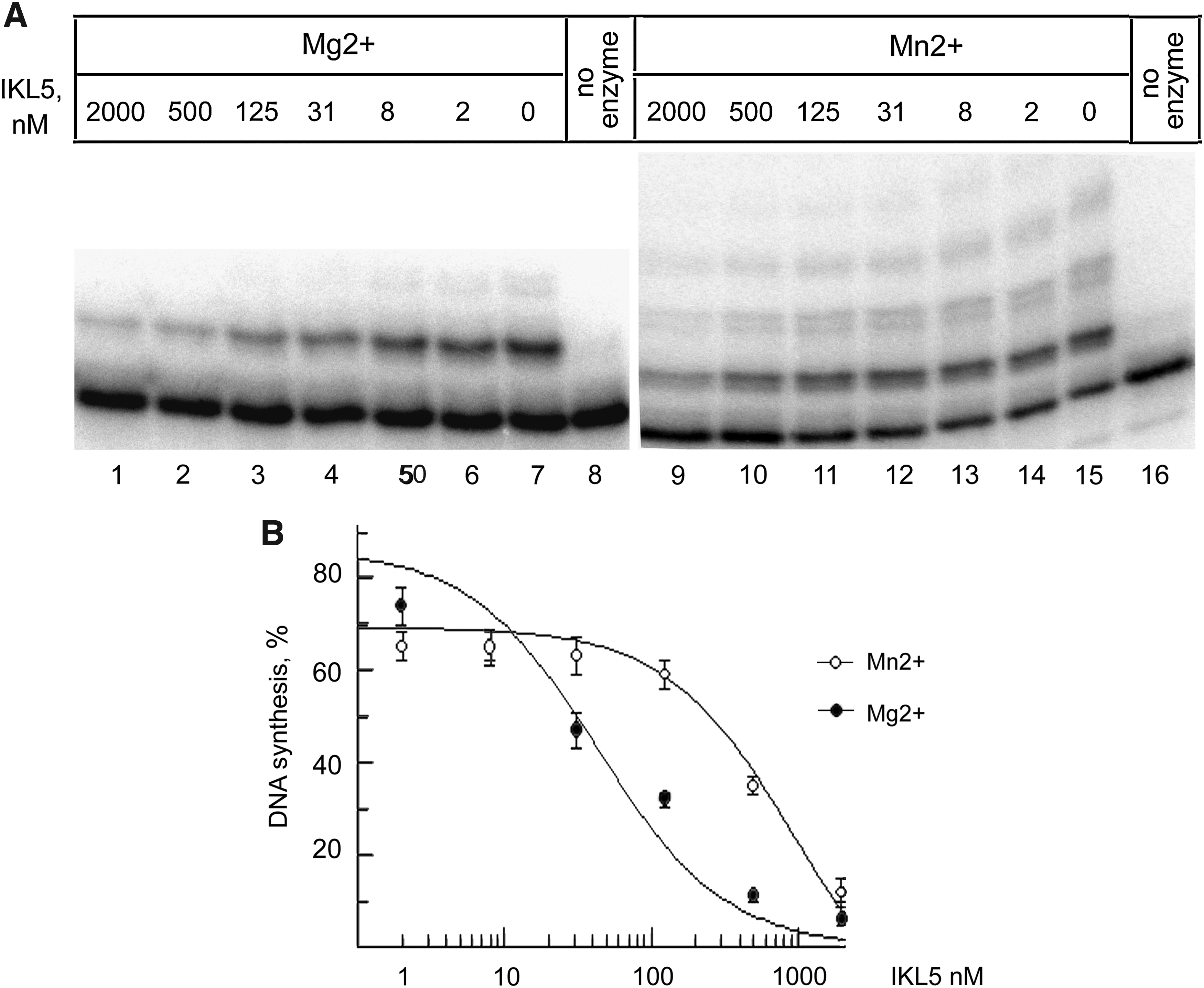
DNA-polymerase activity of Pol i vs. the concentration of aptamer IKL5.
It is known that the presence of Mn2+ in the reaction mixture instead of Mg2+ considerably enhances the DNA-polymerase activity and fidelity of Pol ι (Frank and Woodgate, 2007). Therefore, we studied the effects of IKL5 on the DNA-polymerase activity of Pol ι with Mn2+ as a cofactor. The aptamer IKL5 inhibited Pol ι activity, albeit weakly: IC50 was 930±180 nM (Figs. 3A and 4A, B).
Inhibitory effect of aptamer IKL5 on Pol ι is highly specific
Our previous observation that aptamers had cross-reactivity and aptamers selected for Pol β (Gening et al., 2006b) inhibited the DNA-polymerase activity of Pol κ raised a possibility that aptamer IKL5 may inhibit DNA-polymerase activity of other DNA polymerases. We tested the effect of IKL5 on DNA-polymerase activity of Pol κ, which belongs to the Y family of DNA polymerases, and Pol β, which belongs to the DNA polymerase family X.
IKL5, as well as the initial oligoribonucleotide pool, had practically no effect on the DNA-polymerase activities of Pol κ and Pol β (Fig. 5A). At the same time, the activities of Pol β and Pol κ were clearly inhibited by aptamer 32. These results were confirmed by the kinetics of DNA-polymerase activity of Pol β and Pol κ (Fig. 5B) in the presence and in the absence of aptamer IKL5. We have also shown that aptamer IKL5 has no effect on DNA-polymerase activities of the enzymes which belong to A-family: Pol I from E.coli and Taq I from Thermus aquaticus (data not shown). We conclude that the IKL5 is specific for only Pol ι.
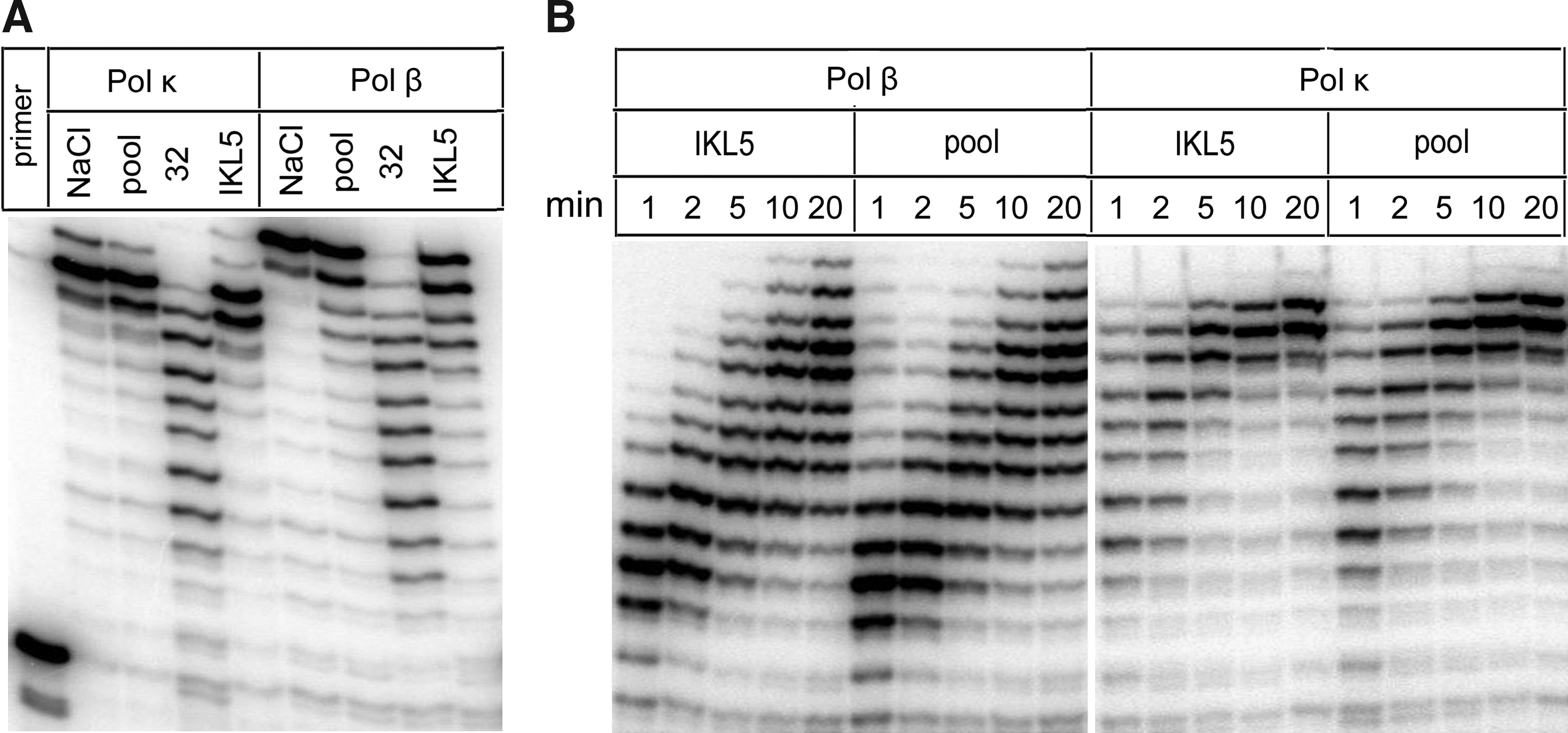
DNA-polymerase activity of Pol κ and Pol β in the presence of aptamer IKL5.
Inhibitory effects of aptamer IKL5 on Pol ι activity in SKOV-3 cell extracts
We tested the effects of various concentrations of IKL5 on the primer extension in the presence of Mn2+ by whole SKOV-3 cell extracts (Fig. 6A). Aptamer IKL5 (500 and 1,000 nM) suppressed the formation of the 18G product attributable to Pol ι activity, and further DNA synthesis after misincorporation of G at the T template is practically completely blocked, a T-stop occurs (Tissier et al., 2000., Zhang et al., 2000). At lower concentrations of IKL5, the intensity of the band corresponding to the Pol ι 18 G product increases and a new band starts to appear, corresponding to the 19-mer product of elongation of the 18G product. The dependence of inhibition of Pol ι activity in SKOV-3 cell extract from IKL5 concentration is shown in Fig. 6A. In this case, the IC50, calculated as described in Materials and Methods, is 207±46 nM (Fig. 6C).
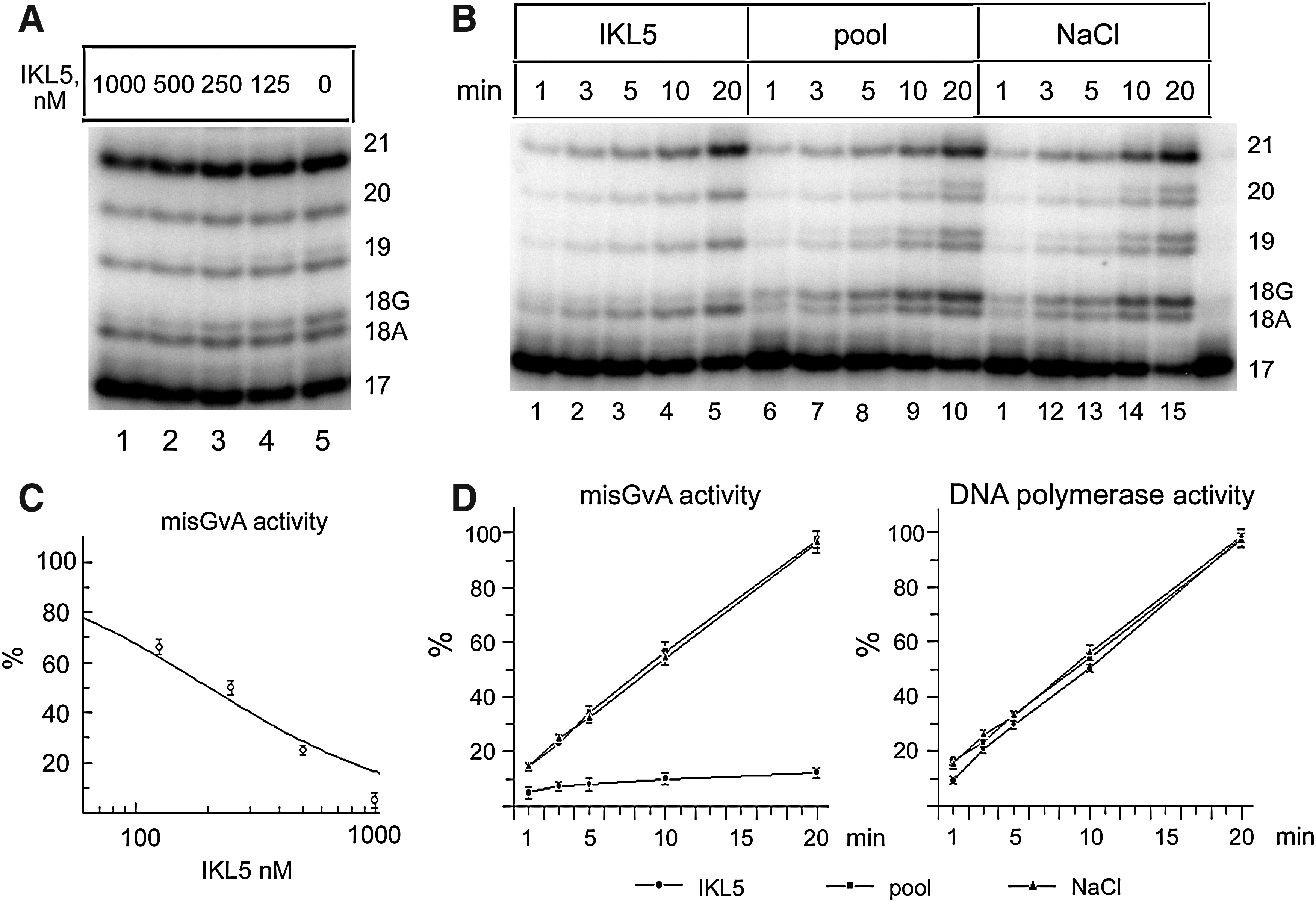
Effect of the aptamer IKL5 on DNA-polymerase activity in SKOV-3 cell extracts.
We have also studied the dependence of DNA synthesis in the SKOV-3 cell extracts in the presence and in the absence of aptamer IKL5 (Fig. 6B). In these experiments, RNA of the oligoribonucleotide pool was used as a control. The addition of the initial oligoribonucleotide pool had no effect on the Pol ι DNA-polymerase reaction, compared to the samples with no RNA added to the reaction mixture. The addition of IKL5 to the reaction mixture resulted in an almost time-independent inhibition of the formation of the 18G product (Fig. 6B, D). The accumulation of products attributable to the activity of other DNA polymerases was unaffected by aptamer IKL5 or by the addition of the initial oligoribonucleotide pool (Fig. 6B, D).
Discussion
The SELEX procedure was successfully used in this work to produce an aptamer for human Pol ι. It binds to this enzyme with high affinity and acts as a highly efficient inhibitor of its DNA-polymerase activity. The aptamer, named IKL5, is the first for a DNA polymerase of the Y family with high affinity. The dissociation constant (Kd) for IKL5 is about 4 nM, two orders of magnitude less than the Kd for the aptamers obtained earlier to Pol β (Kd∼400
The aptamers selected toward a certain human DNA polymerase earlier were cross-reactive. For example, (Gening et al., 2006b) selected 4 aptamers to Pol β, which showed not only binding and inhibition of the activity of Pol β, but also inhibition of Pol κ, a representative of another polymerase family. Here we demonstrate that the SELEX procedure can generate highly specific aptamers for a DNA polymerase. Aptamer IKL5 binds only to Pol ι and inhibits only Pol ι.
This is confirmed by analysis of the effect of the aptamer IKL5 on the Pol ι DNA-polymerase activity vs. total DNA polymerase activity in the extract of cells SKOV-3. The obtained data provide additional validation for the method to test the Pol ι activity in cell extracts from various animal organs and tissues described by us previously (Kazakov et al., 2010; Makarova et al., 2011).
Aptamer IKL5 was a more efficient inhibitor of Pol ι in the presence of Mn2+ ions in the extracts of the SCOV-3 than it was of pure Pol ι. This is unexpected and requires further investigation.
In conclusion, we have successfully developed the first experimental system to test the inhibition of activity of DNA Pol ι in human cell extracts. The experimental system can be a platform for further studies of the biological role of Pol ι.
The aptamer IKL5 obtained in this work can be used in a wide range of research applications. Since Pol ι is the most error-prone eukaryotic DNA polymerase, the cell should strictly control its function. Generally, this could be achieved by protein-protein interactions and post-translational modifications (Friedberg et al., 2002; Friedberg et al., 2005; Plosky and Woodgate, 2004). However, the mechanisms regulating the activity of Pol ι and the contacts of this enzyme with the replication fork components remain largely unknown. An essential step in the studies of the mechanisms regulating Pol ι function in the cell is the identification of the proteins interacting with Pol ι in vivo. Specific binding of aptamer IKL5 to Pol ι allows for isolation of proteins and protein complexes bound to immobilized aptamer from cell extracts under mild conditions, where the degradation of high-order molecular complexes is minimized (Oktem et al., 2007). Aptamer IKL5 may also be useful in the studies of structure and catalytic mechanisms of Pol ι. It is possible to test if aptamer IKL5 inhibits the activity of Pol ι in certain types of malignant cells overproducing the enzyme. This can suppress the genetic instability (Dumstorf et al., 2006; LEHMANN, 2005; Prakash et al., 2005), thus lowering the probability of acquired resistance of a malignant cell to therapeutic drugs.
Footnotes
Acknowledgments
This work was supported by the Russian Foundation for Basic Research, grants 10-04-01434-a and 10-04-00656-a, and by the Russian Ministry of Defense and Science, contracts 14.740.11.0121 and 14.740.11.0171. The work has also been supported in part by the Russian federal program Innovation scientific personnel, State contract 14.740.11.0916 and the State of Nebraska Cancer and Smoking Disease Research Program DHHS grant 2011-27 to Y.I.P.
The authors are grateful to Arthur Grollman (State University of New York, Stony Brook) for initiation of this work, and also to A.V. Kulbachinsky and I.S. Khromov (both from the Institute of Molecular Genetics at the Russian Academy of Sciences) for their valuable advice in the course of this work.
Author Disclosure Statement
No competing financial interests exist.