
Abstract
Small interfering RNAs (siRNA) are able to activate the mammalian innate immune system depending on their structure, sequence, and method of delivery. The immunostimulatory activity of double-stranded RNA can be applied to antiviral and antitumor therapy. Here we identified a set of 19-bp RNA duplexes with 3-nucleotid overhangs in the 3′ ends that display immunostimulating activity (here and after immunostimulating RNA, or isRNA) and studied their sequence/activity relationships. It was found that the introduction of substitutions in the middle part of the isRNA sequence (10–16 positions counting from the 5′ end of strand 1) does not alter the antiproliferative activity, while substitutions in the 3′ end region of isRNA substantially reduce it. isRNAs efficiently inhibit the proliferation of human oral epidermoid carcinoma cells [half-maximal inhibitory concentration (IC50) values varied from 10 to 100 nM]. Our research demonstrated that antiproliferative effects of isRNAs are related to cell growth arrest, rather than the induction of apoptosis. These isRNAs strongly stimulate the synthesis of interferon-α (IFN-α), and to a lesser extent the synthesis of tumor necrosis factor (TNF-α) and interleukin-6 (IL-6), in adherent peripheral blood mononuclear cells. An intravenous injection of isRNA/Lipofectamine complexes into C57BL mice increases IFN-α and IL-6 levels in the blood serum up to 15-fold and 3-fold, respectively, compared to the control mice. The results obtained clearly demonstrate the pronounced immunostimulatory and antiproliferative properties of the isRNAs under study. Hence, these short double-stranded RNAs can be considered as potential agents for the therapy of oncological and viral diseases.
Introduction
In the case of specific gene silencing, the activation of innate immunity by siRNAs is considered an undesirable side effect. However, immunostimulatory siRNAs, which can induce 1 or more undesirable effects, such as proliferation blockage, differentiation, or apoptosis of cancer cells, can be used as potent immunomodulatory agents. The immune system can serve as an extrinsic tumor suppressor (Shankaran et al., 2001; Bui and Schreiber, 2007; Koebel et al., 2007), and immunostimulating agents can be used in antitumor therapy. The antitumor efficiency of siRNAs was demonstrated in 2 recent studies, with 2 simultaneous modes of action: activation of antitumor immunity and silencing of protooncogenes (Poeck et al., 2008; Kortylewski et al., 2009). These findings implied the potential prophylactic and therapeutic use of immunostimulatory siRNAs as adjuvants.
Several dsRNA-based interferon inducers are currently used as immunomodulating, antiviral, and antiproliferative therapeutics. They include poly inosine:cytosine [poly(I:C); “Ampligen”] (Ablashi et al., 1994; Essey et al., 2001; Currie et al., 2008), and dsRNA from virus-like particles from the killer strain of yeasts (“Ridostin”) (Scheglovitova and Ershov, 1995; Masycheva et al., 1997; Bulychev et al., 2003). Interferon-inducing activity of long dsRNAs with specific sequences was investigated earlier by our group (Akimov et al., 2009). We demonstrated that long dsRNAs effectively down-regulate the expression of interferon-sensitive genes and induce antiproliferative and proapoptotic mechanisms in cancer cells.
In this study we identified a set of 19-bp dsRNAs with 3-nucleotide (nt) overhangs in the 3′ ends, which have the ability to efficiently activate innate immunity [here and after immunostimulating RNAs (isRNAs)], and investigated the sequence dependence of their interferon-inducing and antiproliferative activities. We showed that the studied isRNAs efficiently induce interferon-α (IFN-α) and cytokines interleukin-6 (IL-6) and tumor necrosis factor (TNF-α) secretion in both human and mouse immune cells, and that they also inhibit the proliferation of human cancer cells.
Materials and Methods
isRNA
Oligoribonucleotides (Table 1) were synthesized on automatic ASM-800 DNA/RNA synthesizer (Biosset) using ribo-β-cyanoethyl phosphoramidites (Glen Research). After standard deprotection, oligoribonucleotides were purified using denaturing polyacrylamide gel electrophoresis and were then isolated as sodium salts. Oligoribonucleotides were characterized by matrix assisted laser desorption/ionization time-of-flight mass spectra on REFLEX III (Bruker Daltonics). siRNAs were annealed at a concentration of 50 μM in a buffer containing 30 mM HEPES-KOH (pH 7.4), 100 mM sodium acetate, and 2 mM magnesium acetate.
Bold letters indicate the nucleotide differences in sequences of isRNAs within the sequence of isRNA-0; N indicates random nucleotide.
IC50=concentration of isRNA that heralds a 50% decrease in the number of living cells 96 hours after transfection is observed (see Materials and Methods).
A mixture of 19-bp duplexes with 2-nt overhangs (isRNA-0) and 19-bp duplexes with 3-nt overhangs where 3′-terminal nucleotides are random (isRNA-N), representing a mixture of 19-bp duplexes with 2-nt or 3-nt overhangs (isRNA-0&N), was prepared via in vitro transcription with T7 RNA polymerase and linear dsDNA-template according to Ambion technology (available at www.ambion.com/techlib/tn/103/2.html) with modifications. To summarize, in separate reactions, 2 DNA oligonucleotides (5′-AAGTGTCAGGCTTTCAGATTT
Cell culture and transfection of isRNA in vitro
Human carcinoma KB-3-1 cells were purchased from the Institute of Cytology RAS, Russia. The cells were grown in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum, 100 u/m penicillin, 100 μg/m streptomycin, and 0.25 μg/m amphotericin at 37° C in a humidified atmosphere containing 5% CO2/95% air.
Human peripheral blood mononuclear cells (PBMCs) were isolated from the blood of healthy donors by gradient density centrifugation on a Limphocyte Separation Medium (MP Biomedicals), washed, and then resuspended in serum-free DMEM. Enriched monocyte population was isolated via plastic adherence. After 3-hour incubation at a temperature of 37° C, non-adherent cells were removed by repeated gentle washing with the medium.
Cells were plated in 96-well plates (KB-3-1 cells at density 103 cells/well) or 24-well plates (KB-3-1 cells at density 3×104 cells/well and PBMC cells at density 6×105 cells/well) a single day before transfection. On the day of the experiment, the culture medium was changed to serum- and antibiotic-free medium and the cells were transfected with different amounts of isRNA complexed with 0.25 μL (for 96-well plate) or 1 μL (for 24-well plate) of Lipofectamine 2000 (Invitrogen) in OptiMEM according to the manufacturer's recommendations. One-half volume of DMEM with 30% fetal bovine serum was added to the cells 4 hours later.
MTT Assay
The relative amount of living cells was determined using a colorimetric assay based on the reduction of the dye 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) by living cells (Carmichael et al., 1987). Exponentially growing KB-3-1 cells plated as described above in 96-well plates were transfected with 25–200 nM isRNA (see above). After incubation at 37°C for 96 hours, MTT was added to the cells at a final concentration of 0.5 mg/mL. After 3 hours, the culture medium was removed, formosan crystals in each well were solubilized in 100 μL DMSO, and the differences in absorbance at 570 and 620 nm were measured spectrophotometrically.
Flow cytometry
The percentage of apoptotic and dead cells in the population was analyzed using the ApopNexin™ FITC Apoptosis Detection Kit (Millipore) according to the manufacturer's instructions. In brief, the cells were harvested, washed twice with phosphate buffered saline (PBS), and stained with ApopNexin™ FITC and Propidium Iodide (0.2 μg/mL) in 200 μL of PBS for 15 minutes on ice in the dark.
For the analysis of cell cycle, KB-3-1 cells after treatment with one of isRNA preparations were fixed with 70% ethanol, treated with 0.1% Triton X-100 solution containing RNAse A (100 μg/m), and then stained with Propidium Iodide (20 μg/m) for 30 minutes at 37°C.
Stained cells or nuclei preparations were analyzed utilizing flow cytometer Cytomics FC500 (Beckman Coulter) and CXP Software. A total of 30,000 cells were analyzed from each sample. The described above apoptosis and cell cycle analyses were carried out three times in independent experiments and the mean values±standard deviation were used for data presentation.
Analysis of cytokine levels in mouse blood serum
All animal procedures were carried out in accordance with the protocols approved by the Bio-ethics committee of the Siberian Branch of the Russian Academy of Science and recommendations for proper use and care of laboratory animals (European Communities Council Directive 86/609/CEE). Mice were housed in groups of 8–10 individual specimens in plastic cages. The mice had free access to food and water; daylight conditions were normal. Twelve- to thirteen-week-old female C57BL mice were injected intravenously (i.v.) with 200 μL of sterile OptiMEM medium containing 10 μg isRNA or poly(I:C) (about 0.4 mg/kg) and 35 μL of Lipofectamine 2000. Blood was collected 1, 3, 6, 16, or 24 hours after the injection via head clipping. The serum was prepared from the whole blood by coagulation for 30 minutes at 37°C and subsequent centrifugation. The levels of IFN-α and IL-6 were measured using sandwich enzyme-linked immunosorbent assay (ELISA) kits (BD Biosciences), in accordance with the manufacturer's instructions. Two independent experiments were performed and samples were measured in a duplicate.
Analysis of cytokine levels in PBMC cell culture
Freshly isolated PBMCs were transfected with isRNA as described above. Cell-free supernatants were removed 16 hours later and analyzed for human IFN-α, IL-6, and TNF-α levels using a sandwich ELISA (Vector-Best). Analysis was performed in triplicate, and independent experiments were carried out 2 and in some cases 3 times.
Results
Determination of immunostimulating sequence consensus
We previously showed (Kabilova et al., 2006a) that enzymatically prepared siRNA derived from the sequence of the first intron of MDR1 gene and chosen as a negative control efficiently inhibits the proliferation of human oral epidermoid carcinoma KB-3-1 cells and the expression of interferon sensitive genes in these cells. At the same time, its chemically synthesized analogs display no such activities. As a result of the studied siRNA having no mRNA target in the cells and due to fact that it does not participate in the RNA-interference process, we designated it as an isRNA. This isRNA was prepared by in vitro transcription using T7 RNA polymerase as described in the materials and methods section. Our analysis of the enzymatic products revealed the presence of the supplementary product along with the target 19-bp with 2-nt overhangs (19+2) product (Kabilova et al., 2006b), which was formed due to non-template addition of an arbitrary nucleotide to the 3′ end of growing RNA chain by T7 RNA polymerase (Milligan et al., 1987). We believe that this 19-bp with 3-nt overhangs (19+3) product is responsible for the activation of innate immunity and antiproliferative activity of enzymatically prepared siRNA. We proved this assumption by comparing the antiproliferative activity with respect to KB-3-1 cells of chemically synthesized 19+2 isRNA-0, 19+3 isRNA-N with random extra nucleotide on its 3′ ends and enzymatically prepared isRNA-0&N that contained a mixture of 19+2 and 19+3 products. Poly(I:C) (5–500 ng/mL) was used as a positive control of activation of innate immunity. Both enzymatically and chemically prepared 19+3 isRNA (isRNA-0&N and isRNA-N) almost halved the number of living cells when used at concentrations of 100–200 nM. In contrast, 19+2 isRNA-0 had no effect on the number of living cells (Fig. 1). These results confirmed our assumption that the 19-bp isRNA with 3-nt overhangs on 3′ ends exerts antiproliferative activities in KB-3-1 cells. Control interferon inducer poly(I:C) reduced the number of living cells to 50% relative to mock-transfected cells at concentration 50 ng/mL. The increase of poly(I:C) concentration up to 500 ng/mL resulted in a massive rate of cell death. This was the reason why we began to use poly(I:C) at a concentration of 50 or 250 ng/mL.

The effect of immunostimulating RNA (isRNA) or poly(I:C) on the proliferation of KB-3-1- cells. Bars show the relative number of living KB-3-1 cells 96 hours after transfection of chemically (isRNA-0, isRNA-N) or enzymatically (isRNA-0&N) synthesized isRNAs, normalized to the number of Mock-transfected cells (cells treated with Lipofectamine only). Experiments were performed in triplicate. Error bars are representative of standard deviations from the mean. Poly(I:C), poly inosine:cytosine.
In order to determine the effective immunostimulating RNA motifs we compared different anti-proliferative activities of chemically synthesized 19+3 isRNA with random nucleotide N or nucleotides A, G, C, or U added to its 3′ ends (isRNA-N, -A, -G, -C, and -U). Subsequently, we analyzed the antiproliferative activity of the heteroduplexes formed by 21 and 22-nt RNAs (isRNA-N/0; isRNA-0/N). In this experiment we transfected KB-3-1 cells with a variety of isRNAs taken at concentrations from 10 to 200 nM, and after 96 hours we performed the MTT test. The isRNA sequences and IC50 values corresponding to 50% reduction of the number of living cells are listed in Table 1. It was found (Table 1) that isRNA-N had IC50 of 145nM, whereas IC50 for isRNA-N/0 and isRNA-0/N increased up to 200 nM. These data affirm that only 19-bp isRNA with 3-nt overhangs exhibited antiproliferative activity. The investigation of isRNAs with defined 3′ end nucleotides (Table 1) showed, that only isRNA-U (U at the 3′ end of both strands; IC50 36 nM) and isRNA-A were able to inhibit cell proliferation, whereas isRNA-G and isRNA-C are not active (IC50 >200 nM). Among isRNAs with “mixed” strands, only isRNA-U/A (IC50 30 nM) but not isRNA-A/U (IC50 100 nM) and corresponding single strands displayed a significant antiproliferative activity.
In order to determine other 3′-overhang sequence determinants that provide for the immunostimulatory activity of isRNA, we synthesized a set of isRNA derived from the sequence of isRNA-U and examined their effect on KB-3-1 cells proliferation (Table 1). To determine consensus immunostimulating motifs in isRNA, we investigated the antiproliferative activity of isRNA-U with substitutions in different parts of its sequence (shown in bold in Table 1). It is seen that substitution of nucleotides in the central part of isRNA-U at the positions from 10 to 12 [isRNA-U(10–12): UUU-motif in strand 1 was substituted by AGA] and positions 13–16 [isRNA-U(13–16): CAGA-motif in strand 1 was substituted by UUUU forming oligoU13 3′ end] lead to isRNA with higher antiproliferative activity as compared with the parent, isRNA-U. The values of IC50 were 15.5, 17.5, and 36 nM, respectively. Substitution of UGU motif in the strand 1 by AGA disrupting GUGU sequence at positions 1–4 at the 5′ end of strand 1 [isRNA-U(2,4)] only slightly reduced its antiproliferative activity in comparison with isRNA-U (IC50 ∼40 nM versus 36 nM for parent isRNA-U). On the contrary, the only substitution U→C in isRNA-U(20) that disrupts the oligoU motif at the 3′ end of strand 1, abolishes the antiproliferative activity of the corresponding isRNA (IC50>200 nM). The extension of 3′ overhangs from 4 to 11 nt (isRNA-U2–9) did not considerably change the antiproliferative activity of isRNA: IC50 was found to be of 32, 44, and 51 nM for isRNA-U2, isRNA-U5, and isRNA-U9, respectively. Thus, the immunostimulating properties of the isRNA under study are tolerated to the substitutions in the middle part of isRNA molecule; however, oligoU motif at the 3′ end of strand 1 is important for this function. Thus, we selected a set of isRNAs exhibiting pronounced antiproliferative activity with IC50 varying from 15.5 nM to 36 nM, isRNA-U(10–12) was revealed to be the most active among them. Data obtained permitted the identification of the consensus sequence of immunostimulating RNA: strand 1, 3′-A/UUUUUUN8N9N10N11N12N13N14CGGACUGUG-5′, and strand 2, 5′-AAAN1N2 N3 N4 N5 N6 N7GCCUGACACUUU/A-3′, where N1–N7 are complementary to N8–N14, respectively.
Cytokine induction by isRNA in human PBMC
Human monocytes/macrophages that were found in a fraction of adherent PBMCs are essential effector cells in inflammatory disorders and infectious diseases. In order to test whether the antiproliferative activity of isRNAs on KB-3-1 cells is related to innate immunity activation we analyzed IFN-α, TNF-α, and IL-6 cytokine production upon stimulation of human PBMCs with the isRNAs. Sixteen hours after transfection, IFNs and proinflammatory cytokines were detected in cell cultural medium by sandwich ELISA. This time point was chosen on the basis of our preliminary data on the cytokine production kinetics (data not shown). The PBMCs were treated with isRNA/Lipofectamine complexes at a concentration of 25–100 nM. It was shown (Fig. 2) that isRNA-U/A, which exhibits a high antiproliferative activity in KB-3-1 cells, also induces IFN-α secretion in a dose-dependent manner in PBMCs (Fig. 2A). Incubation of the cells with isRNA-U/A (200 nM) leads to a ∼4-fold increase of IFN-α secretion, relative to the mock-transfected cells, whereas no changes to the level of IFN-α is observed for isRNA-0 and poly(I:C). The data reveal (Fig. 2B), that some of isRNAs induce also TNF-α and IL-6 secretion in PBMCs. The most effective activators of secretion of proinflammatory cytokines are single-stranded (ssRNA)-U and isRNAs containing this strand, notably isRNA-U and isRNA-U/A: They increase IL-6 and TNF-α levels by 1.6- to 2-fold and 5.5 to 6.5-fold, respectively, relative to the mock-transfected cells. isRNA-0, isRNA-G, isRNA-C, and isRNA-A did not display any activity. isRNA-U(2,4), isRNA-U(10–12), isRNA-U(13–16), and isRNA-U2 induce 3.5- to 4-fold increase in TNF-α level, whereas isRNA-U(20), isRNA-U5, and isRNA-U9 were not active (data not shown). All of them do not activate IL-6 production by PBMC (data not shown). The control innate immunity stimulator poly(I:C) induces the highest level of proinflammatory cytokines TNF-α and IL-6 (15-foldsand 2-fold activation relative to the mock-transfected cells), whereas the level of IFN-α is comparable with that in mock-transfected cells. These results reveal that the isRNA-U/A efficiently activate IFN-α secretion and increase the level of TNF-α, and to a lesser degree IL-6 secretion. Poly(I:C), on the contrary, induces secretion of proinflammatory cytokines but not IFN-α.
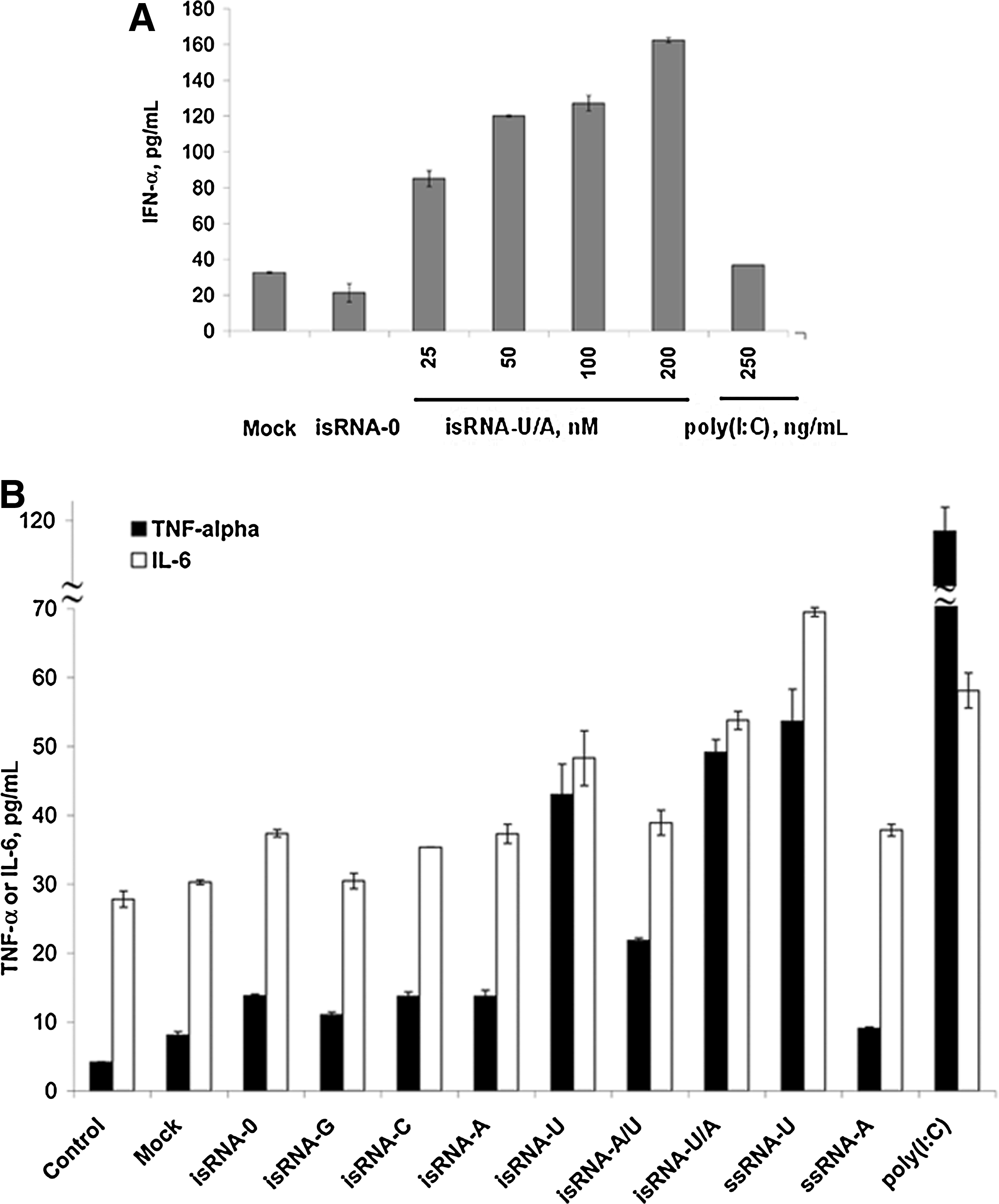
The effects of isRNAs on IFN-α
isRNAs induce cell growth arrest rather than cell death by apoptosis
In order to test whether the antiproliferative activity of isRNAs is related to the induction of apoptosis or to cell growth arrest, we selectively stained living, dead, and apoptotic cells using ApopNexin™ FITC Apoptosis Detection Kit (Millipore). Subsequently, the cell samples were analyzed by flow cytometry 48 hours post-transfection with isRNA-U/A (100 nM) or poly(I:C) (50 ng/mL) complexed with Lipofectamine. All cells grown in the experimental wells were collected, including floating cells. We found that the Lipofectamine cause a moderate toxic effect in KB-3-1 cells resulting in the slow accumulation of apoptotic (4.5%) and dead (6%) cells in the population (Fig. 3A, “Mock”). The positive control poly(I:C) caused a pronounced proapoptotic action: In these samples about 30% of apoptotic and 20% of dead cells were detected. In contrast, isRNA-U/A induces only a slight increase in the number of apoptotic (8%) and dead (6.5%) cells. Since antiproliferative activity of isRNA-U/A and poly(I:C) at the concentrations used was similar, we conclude that isRNAs reduce the number of living cells by arrest of cell growth rather than by induction of apoptosis.
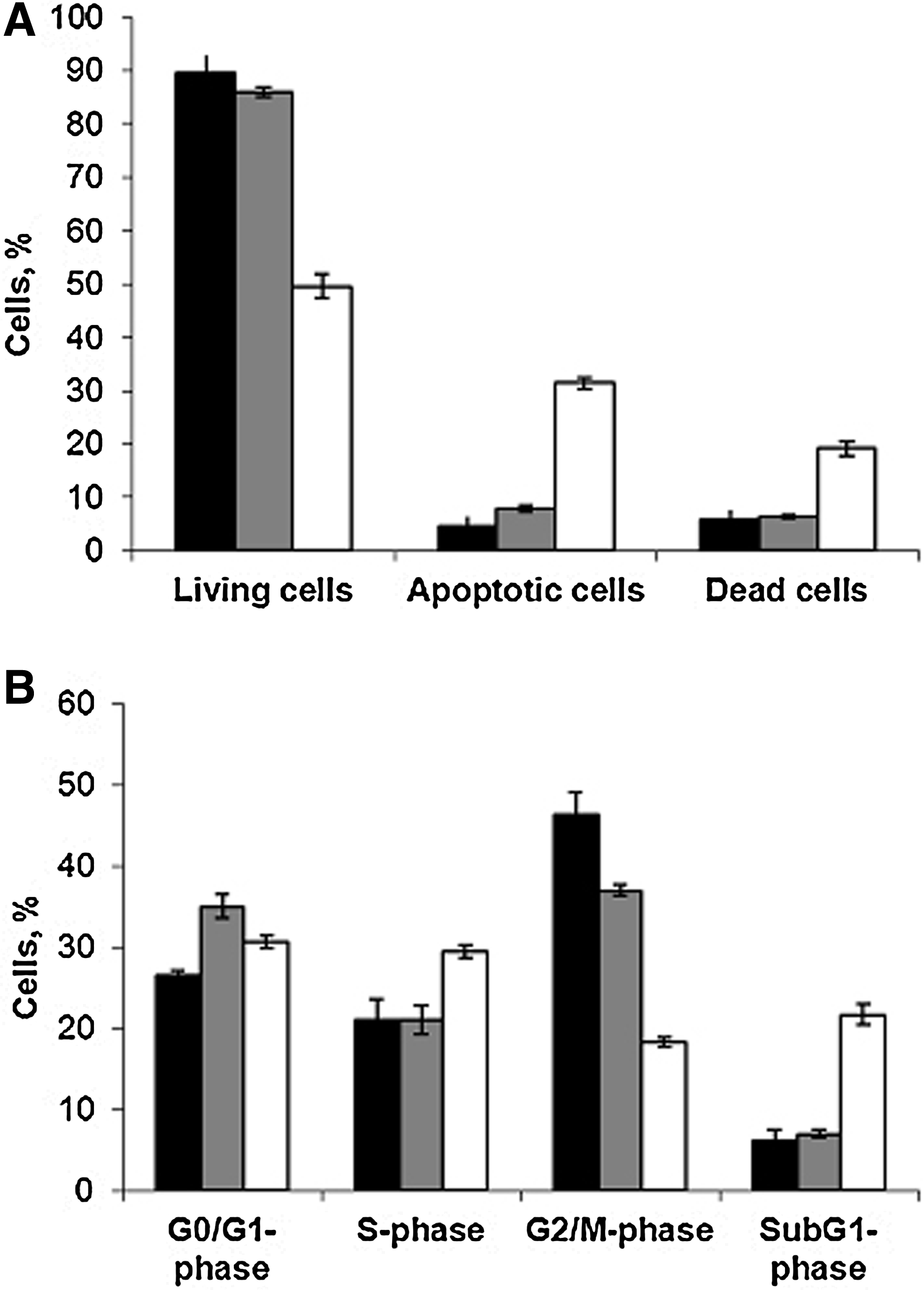
The effects of isRNA and poly(I:C) on cell death
In order to confirm this hypothesis we analyzed the distribution of KB-3-1 cells between cell cycle phases by flow cytometry 48 hours after isRNA-U/A (100 nM) or poly(I:C) (50 ng/mL) transfection. It was found (Fig. 3B) that in the presence of isRNA-U/A the number of cells in G0/G1-phase increases, and the number of cells in G2/M-phase decreases, while the number of cells in S-phase does not alter significantly. The increase of up to 22% of subG1-phase cell population (apoptotic cells) was detected in the samples transfected with poly(I:C). In contrast 6% to 7% of subG1-phase cell population was detected in the samples transfected with either isRNA-U/A/ Lipofectamine complexes or Lipofectamine only. These data are consistent with the data obtained using ApopNexin™ FITC Apoptosis Detection Kit. Despite increasing the number of G0/G1- and S-phase cells, poly(I:C) significantly reduces the number of cells in G2/M phase. Obtained data indicate that poly(I:C) induces apoptosis mainly in dividing cells. These results support the hypothesis, that isRNA-mediated antiproliferative effects are related to the cell growth arrest and reveal the difference in the mechanisms of antiproliferative action of poly(I:C) and isRNA.
Cytokines induction by isRNAs in mice
In order to investigate immunostimulation activity of isRNAs in vivo, we assayed the levels of IFN-α and IL-6 in mouse blood serum after intravenous administration of isRNAs or poly(I:C). Groups of 3 C57BL mice were injected with isRNAs/ Lipofectamine (10 μg of isRNA per mouse). At different time points (1–24 hours) the blood samples were collected and the levels of cytokines were measured by ELISA. The results reveal (Fig. 4) that both isRNA-U/A and poly(I:C) induce IFN-α and IL-6 secretion in mouse blood serum. isRNA-U/A induces 5-fold increase of IFNα level in serum 3 hours after injection (Fig. 4A), and the maximum level of IFNα synthesis was observed 6 hours after injection, up to a 15-fold increase relative to the level in the mice injected with OptiMEM only (“Control”) or OptiMEM with Lipofectamine (“Mock”). After 16 hours, the IFNα level is decreased, and it reaches the original level by 24 hours. A similar time-dependence was observed for poly(I:C), but the IFNα level was 2-fold higher than that for isRNA-U/A. The most effective activation of IL-6 synthesis (Fig. 4B) was observed already 1 hour after injection of isRNA-U/A or poly(I:C) preparations. Thus, the isRNA-U/A induces 3-fold and poly(I:C) induces 18-fold increases of IL-6 level in serum relative to the level in control mice. Injections of Lipofectamine (“Mock”) without isRNA also induced a 2-fold increase of IL-6 level. After 3–6 hours, the IL-6 level was decreased and reached the same level as in control mice 1 hour after injection.
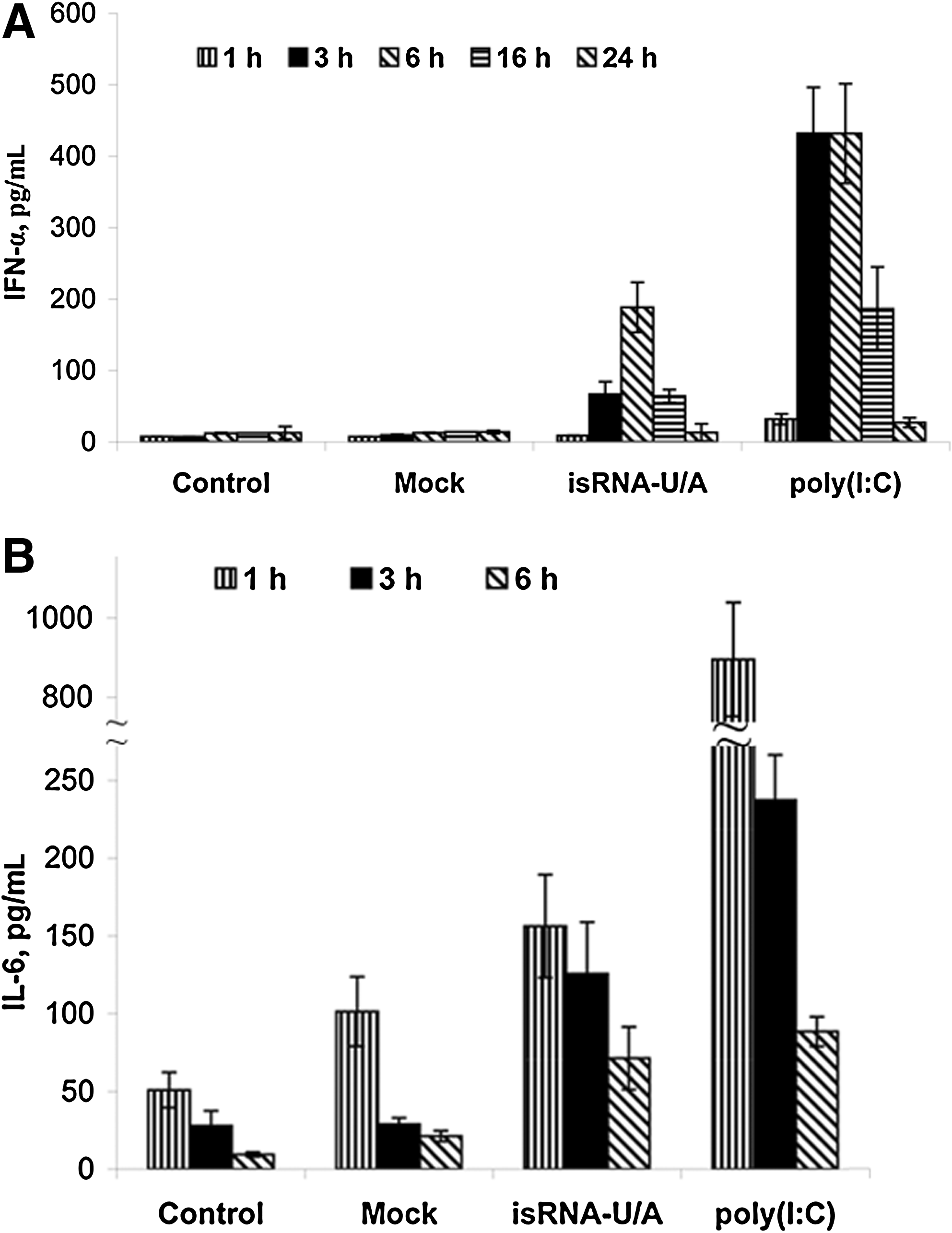
Stimulation of the innate immune response by isRNA and poly(I:C) in vivo. C57BL mice were intravenously injected with isRNA-U/A or poly(I:C) at a dose 0.4 mg/kg in the complex with Lipofectamine™2000. Serum IFN-α
Discussion
In this study we demonstrate that 19-bp double-stranded isRNA with 3-nt overhangs on 3′ ends can efficiently activate innate immunity in vitro and in vivo. Our results on human PBMC and carcinoma KB-3-1 cells demonstrate that the immunostimulatory properties of isRNAs depend on the sequence and that the presence of the last nucleotide at the 3′ ends of molecule is crucial for this function (Fig. 5). Thus, 19+2 isRNA-0 does not affect proliferation of KB-3-1 cells, whereas 19+3 isRNA-N with an additional random nucleotide at the 3′ ends reduces cell proliferation to a significant extent. We found that antiproliferative activities of 21/22-nt heteroduplexes isRNA-N/0 and isRNA-0/N are lower than that of 19+3 isRNA-N, indicating the relevance of 3-nt 3′ overhangs of isRNA. Moreover, we showed that only isRNAs containing 3′ end U, A, or their combination display immunostimulatory properties, whereas isRNAs with 3′ end G or C are not active. The extension of 3′-U overhangs from 4 to 11 nt led to some decrease of the immunostimulatory activity of isRNAs.
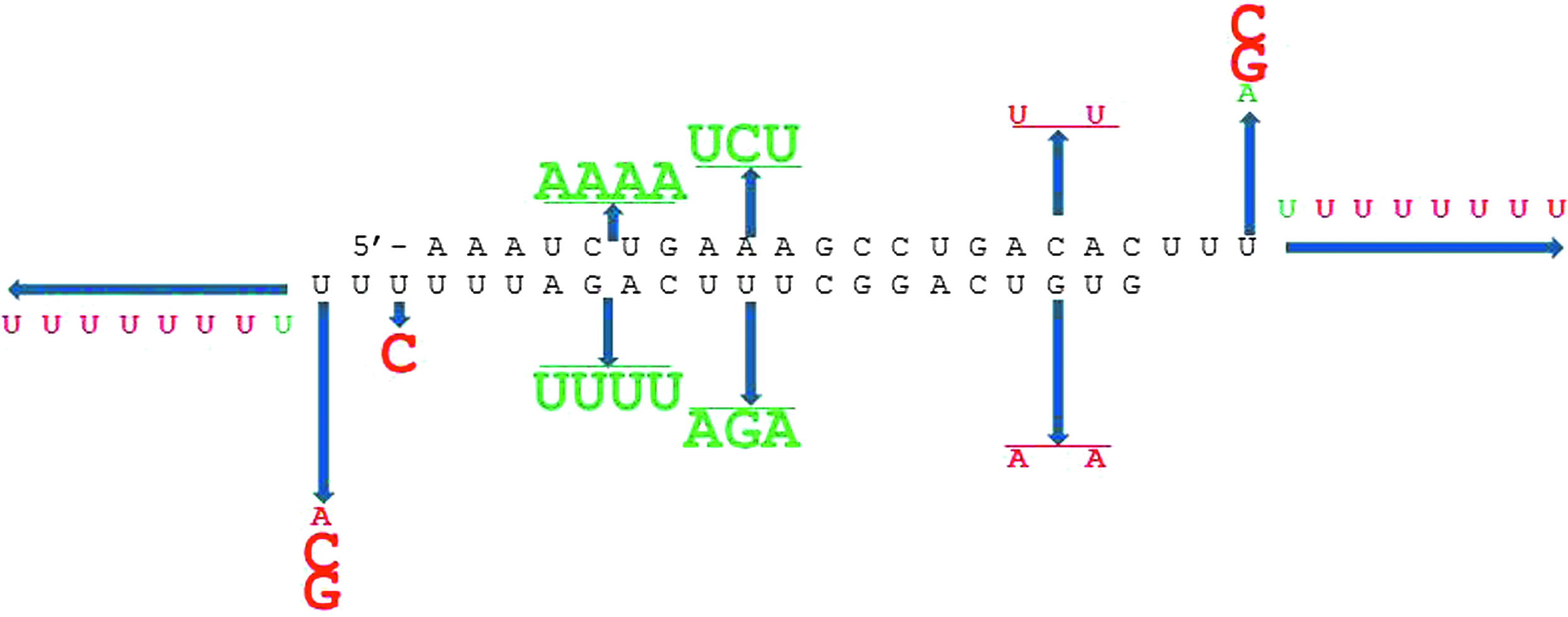
The influence of the substitutions on the antiproliferative activity of is RNA-U. Substitutions that positively affected the activity of isRNA are shown in green; detrimental substitutions are shown in red. The size of the letters correlates with the level of the effect.
Previous publications have reported that certain siRNA motifs are capable of activating the innate immunity via TLRs (Hornung et al., 2005; Judge et al., 2005; Diebold et al., 2006; Forsbach et al., 2008). A number of immunostimulatory motifs have been identified in literature: GU-based 5′-UGUGU-3′ (Judge et al., 2005), 5′-GUCCUUCAA-3′ (Hornung et al., 2005) that are ligands of TLR7/8, repeats of 2 or 3 uridine clusters that allow RNA to act as agonists of TLR7 (Diebold et al., 2006); and AU-rich siRNAs that mediate human TLR8 activation (Forsbach et al., 2008). Herein we introduced substitutions in different parts of the isRNA-U sequence and investigated antiproliferative and interferon-inducing activities of these isRNAs. It was found (Table 1; Fig. 2B) that disruption of the oligoU-motif in the center of the molecule from 10 to 12 positions [isRNA-U (10–12)], as well as substitutions at positions 13–16 [isRNA-U(13-16)] forming oligoU13-motif, increase the antiproliferative activity of respective isRNA in comparison to the activity of parent isRNA-U. Disruption of the GUGU motif, which is similar to the reduced version of the immunostimulatory sequence (Judge et al., 2005) at the 5′ end of strand 1 [isRNA-U(2,4)], caused a minor decrease to the antiproliferative activity compared with the parent isRNA-U. In contrast, introduction of only 1 substitution that disrupts oligoU6 motif at the 3′ end of strand 1 [isRNA-U(20)] entirely abolishes the antiproliferative activity. Thus, immunostimulating properties of the studied isRNAs are tolerant to the substitutions in the middle part of isRNA molecule. However, the sequence of the 3′ ends of isRNA are important for this function. Based on our studies, we concluded that the availability of oligoU motif at the 3′ end of strand 1 and of 3-nt 3′ overhangs are crucial for the immunostimulatory activity of isRNAs.
In order to clarify the mechanism of antiproliferative activity of studied isRNAs on carcinoma KB-3-1 cells, we performed an apoptosis assay and analyzed the distribution of KB-3-1 cells between phases of cell cycle after isRNA-U/A transfection. Our analysis showed (Fig. 4A, B) that isRNA-U/A, in contrast to control apoptosis inductor poly(I:C), does not induce apoptosis, but rather increases the number of cells in G0/G1 phase and reduces the number of cells in G2/M phase 48 hours after transfection. Distribution of the cells between cell cycle phases at 72 hours after isRNA transfection is similar to that at 48 hours after transfection (data not shown). The results suggest that antiproliferative effects of isRNAs are related to cell growth arrest rather than induction of apoptosis.
Analysis of immunostimulatory activity of isRNAs in human PBMC in vitro revealed that isRNA-U/A (100 nM) induces a 4-fold increase in IFN-α level; in contrast, the control innate immunity stimulator poly(I:C) (250 ng/mL) does not activate IFN-α secretion relative to mock-transfected cells. Furthermore, poly(I:C) increased the level of proinflammatory cytokines TNF-α and IL-6 by 15-fold and 2-fold, respectively, when isRNA-U/A induces a 6-fold increase in TNF-α level and 1.6-fold increase of IL-6 level relative to the mock-transfected cells. High levels of TNF-α and/or IL-6 are associated with the development of cytokine-mediated toxicities and inflammation (Judge et al., 2008). Our results (Table 1; Fig. 2B) indicate that there is no direct correlation between antiproliferative and immunostimulatory activities for isRNAs under study; however, all isRNAs demonstrating no or low antiproliferative activity (IC50>200 nM) in KB-3-1 cells were not able to activate IFN-α, IL-6 and TNF-α production in PBMCs. We assume that the antiproliferative effect of isRNAs in KB-3-1 cells is a result of activation of innate immunity, but the levels of the observed effects are different in the 2 cell types because of the different expression pattern of RNA-recognizing receptors expressed by tumor cells and PBMCs. So, TLRs are expressed mainly by immune cell types. Besides the TLR system, mammalian cells have evolved a number of TLR-independent mechanisms, recognizing RNA, including siRNA, in the cellular cytoplasm, and inducing via the dsRNA-binding protein kinase R or RIG-1 proteins (Robbins et al., 2009).
To investigate immunostimulatory activity of isRNAs in vivo, we assayed IFNα and IL-6 levels in mouse blood serum after isRNAs i.v. administration. The control type 1 interferons inducer poly(I:C) (Currie et al., 2008) was used. We showed that injection of naked isRNA (data not shown) or Lipofectamine alone (Fig. 4) did not activate innate immunity significantly, but 1 hour after injection of Lipofectamine alone, a slight and transient increase (2-fold) of IL-6 level was observed. Injections of isRNAs or poly(I:C) complexed with Lipofectamine stimulate the elevation of serum IFN-α and IL-6 levels by 6 hours and 1 hour after injections, respectively (Fig. 4A, B). Similar results were reported by other groups (Hornung et al., 2005; Judge et al., 2005; Shin et al., 2007; Poeck et al., 2008); their research showed that 6–7 hours after injection of siRNA complexed with cationic liposomes in mice, systemic immune responses were induced, accompanied by IFN-α, IL-6 and/or IFN-γ production. Our results indicate (Fig. 4) that the isRNA-U/A induces a strong 15-fold increase of IFNα level and only a 3-fold increase of IL-6 level relative to the level in control mice. Although the poly(I:C) activates the synthesis of IFNα more effectively than isRNA, it induces as high as an 18-fold increase of IL-6 level. These data are in accordance with in vitro results indicating that poly(I:C) induces the highest level of TNF-α, up to 15-fold. In vitro and in vivo data show that isRNAU/A mainly induces the synthesis of IFNα but not proinflammatory cytokines TNF-α and IL-6, whereas the poly(I:C), along with IFNα, activates a high level of proinflammatory cytokines. Thus, isRNAs under study have advantages as type 1 interferon inducers over poly(I:C), which induces the toxic effects related to inflammation.
The difference in the spectrum of the interferon-α and cytokines IL-6 and TNF-α, induced by isRNA or poly(I:C) in experiments in vitro and in vivo, can be accounted for by the difference in the effector cells, represented in the mice blood and in cell culture of human PBMCs. The producers of IFN-α are blood mononuclear cells (Stewart et al., 1979), IL-6 can be produced by a wide variety of cells—monocytes, activated T-cells, macrophages, endothelial cells, and fibroblasts (KISHIMOTO, 1989); whereas the main sources of TNF-
Our results show that 19-bp-long isRNAs with 3-nt overhangs on 3′ ends induce innate immunity in a sequence-dependent manner. This isRNA possesses a high level of antiproliferative and immunostimulating activities and can be used as an adjuvant in antivirus and antitumor therapy. However, the question of effective in vivo delivery yet needs to be solved.
Footnotes
Acknowledgments
This research was supported by the Russian Academy of Science under the programs Molecular and Cell Biology (grant no. 22-1) and Science to Medicine (grant no. 37), as well as the President's program for support of leading scientific schools (grant no. NSh-7101.2010.4), the Russian Foundation for Basic Research (grant no. 11-04-01017-a) and the Siberian Branch of Russian Academy of Sciences (grant no. 41).
Author Disclosure Statement
No competing financial interests exist.