
Abstract
Streptococcus mutans is one of the major cariogenic bacteria in the oral environment. Small non-coding RNAs (sRNAs) play important roles in the regulation of bacterial growth, stress tolerance, and virulence. In this study, we experimentally verified the existence of sRNA, L10-Leader, in S. mutans for the first time. Our results show that the expression level of L10-Leader was growth-phase dependent in S. mutans and varied among different clinical strains of S. mutans. The level of L10-Leader in S. mutans UA159 was closely related to the pH value, but not to the concentrations of glucose and sucrose in culture medium. We predicted target mRNAs of L10-Leader bioinformatically and found that some of these mRNAs were related to growth and stress response. Five predicted mRNA targets were selected and detected by quantitative reverse transcriptase-polymerase chain reaction (qRT-PCR), and we found that the expression levels of these mRNAs were closely related to the level of L10-Leader at different growth phases of the bacteria. Our results indicate that L10-Leader may play an important role in the regulation of responses in S. mutans, especially during its growth phase and acid adaption response.
Introduction
Major sRNAs interact by pairing with their target mRNAs, acting as antisense RNAs. Some sRNAs form parts of RNA–protein complexes with the involvement of chaperone protein, Hfq. A few sRNAs regulate by binding with a target protein (Vogel and Wagner, 2007). Many sRNAs, such as the MicF, OxyS, DsrA, Spot42, and RyhB RNAs, act by base pairing to activate or repress translation, or to destabilize mRNAs. Major groups are based on their mode of action; these are cis- and trans-acting regulatory sRNAs. Riboswitches and RNA thermometers (Narberhaus et al., 2006) belong to the class of cis-acting regulatory RNAs, which are located at the 5′-untranslated region (UTR) of their genes. Riboswitches modulate their regulatory structure in response to metabolite binding, which are present in the shared environment. RNA thermometers are involved in sensing global signals. Both cis- and trans- regulatory RNA structures are essential for fine regulation systems, to ensure an immediate physiological response within the bacterial cell to a variable environment. As the function of sRNAs becomes increasingly clear, there is also the prospect of broad applications of sRNAs, such as in the design of novel potential therapeutics based on sRNA-complementary peptide nucleic acids (Karkare and Bhatnagar, 2006) and developing a novel ultrasensitive diagnostic system, which has been conceived for the detection of small target samples (Mraheil et al., 2010).
Streptococcus mutans is a Gram-positive facultative anaerobic bacterium. It is the main etiological agent in the development of dental caries (Gibbons and van Houte, 1975). S. mutans possesses a variety of virulence factors that enable it to accumulate in large numbers, colonizing the tooth surface. It uses a wide array of carbohydrate sources, and is able to produce and tolerate acid, being able to survive at low pH. It adheres to the surfaces of teeth, which results in the demineralization of tooth surfaces (BURNE, 1998). The regulatory mechanism of acid adaption and the virulence factors required are not clear, despite much research having been performed. To date, there has not been confirmation of the sRNAs present in oral bacteria. With a greater knowledge of sRNAs in oral bacteria, especially in S. mutans, we will better understand oral infectious diseases, such as dental caries.
This study was carried out using bioinformatic approaches to predict sRNAs in S. mutans, followed by confirmation using experimental techniques. The sRNA sequence was found through the analysis of the Rfam website (http://rfam.sanger.ac.uk/), and annotated as L10-Leader (RF00557) family. We detected all 40 sRNAs predicted by at least 2 software application methods, including the software and the web page by quantitative reverse transcriptase-polymerase chain reaction (qRT-PCR). The transcript level of L10-Leader was relatively high compared with the other sRNAs and the changes were more significant. So L10-Leader was chosen for the further study. We detected the levels of L10-Leader in different growth phases, different clinical strains, and different growth media. Additionally, we predicted 5 target mRNAs for L10-Leader computationally. This study was the first to attempt to identify sRNA in oral bacteria, and the first to find sRNA-L10-Leader in S. mutans.
Materials and Methods
Strains and growth media
Streptococcus mutans UA159 and 2 clinical strains of S. mutans were used in this study. All S. mutans strains were routinely grown in brain heart infusion (BHI) medium (Becton Dickinson) or Todd-Hewitt broth (Becton Dickinson) supplemented with 1% yeast extract at 37°C in a 5% CO2 atmosphere. For growth studies, cultures were grown overnight in BHI broth and then diluted 1:50 into fresh BHI broth and grown to the necessary growth phase [optical density at 600 nm (OD600)=0.2, 0.4, 0.5, 0.6, 0.8, 1.0, and 1.5). Tryptone yeast extract medium (TYE) (10% tryptone, 5% yeast extract, 17.2 mMK2HPO4) was utilized for acid shock. For acid shock assays, HCl was added to TYE medium to adjust final the pH to 5.5 or 7.5. For sugar concentration assays, 0.1% or 1% glucose and 0.5% or 1% sucrose were added to TYE.
For acid shock assays, overnight cultures of S. mutans UA159 were grown in THYE and diluted 10-fold in TYE supplemented with 0.5% glucose at pH 7.5 and incubated at 37°C until cells reached the mid-logarithmic phase (OD600=0.4). Cell cultures were then divided into 2 aliquots, pelleted by centrifugation, and resuspended in TYE supplemented with 0.5% glucose at pH 5.5 or 7.5. The cells were subsequently incubated for 0.5, 1.5, 2.5, 3.5, or 4.5 hours at 37°C, harvested by centrifugation, and utilized for RNA isolation.
For different sugar concentration assays, overnight cultures of S. mutans UA159 were grown in THYE medium and later diluted 1:20 using sterile TYE supplemented with 0.1% or 1% glucose and 0.5% or 1% sucrose for 24 hours at 37°C, harvested by centrifugation, and utilized for RNA isolation.
Bioinformatic identification of sRNAs in S. mutans UA159 genome
The genome of S. mutans UA159 (NCBI No.: NC_004350) was analyzed with sRNA Predict, sRNASVM, and SIPHT software for bacterial genomic sRNA prediction. SIPHT (sRNA identification protocol using high-throughput technology) and sRNAPredict predict candidate sRNA-encoding loci, identified based on the presence of putative Rho-independent terminators downstream of conserved intergenic sequences, and each locus is annotated for several features, including conservation in other species, association with one of several transcription factor binding sites and homology to any of over 300 previously identified sRNAs and cis-regulatory RNA elements (LIVNY, 2007; Livny et al., 2008). sRNASVM is a model for the prediction of small non-coding RNAs in E. coli using support vector machines (Saha and Raghava, 2006). The web page “Oral Pathogens Non-Coding Small RNA Prediction” (Los Alamos National Laboratory Oral Pathogen Sequences Databases; http://www.oralgen.lanl.gov/oralgen/bacteria/analysis/srna_result/index.html) also predicted the sRNA of S. mutans.
qRT-PCR
Protocols for the isolation of RNA from S. mutans call for the pre-treatment of bacteria with lytic enzymes in TE buffer. Total RNA was then extracted using Trizol Reagent (Invitrogen) according to manufacturer's instructions. Contaminated genomic DNA was removed by DNase I (Promega). RNA was reverse transcribed using M-MLV reverse transcriptase (Promega), according to the manufacturer's protocol. Quantitative PCR was performed according to the protocol of the SYBR Green qPCR Master Mix (2×) (Fermentas) with Mx3000p (Stratagene); 16S mRNA levels were used for normalization. The experiment was repeated at least 3 times.
The primer sequences were: 16S forward: CTTACCAGGTCTTGACATCCCG; 16S reverse: ACCCAACATCTCACGACACGAG; L10-Leader forward: CTAAGACAGCAGGGGAGCGT; L10-Leader reverse: GTCTGGGTTTGGGTGCGATT; T7 promoter sequence: TAATACGACTCACTATAGGG; M1 forward: AAAAGCCCCTCTTGTCAATG; M1 reverse: TCTGTGACATCTGGTTGAAAAGT; M2 forward: GTGGCGATCCAACTGGTACT; M2 reverse: TAGGCTGCTGCAATTTCCTT; M3 forward: CGTCTTCTCAACAACCAGCA; M3 reverse: TTTGAACGCCCATAAAGTCC; M4 forward: TGACCGTGGTGACAGGTCTA; M4 reverse: ATCCGCCTTTCCGATAAGTT; M5 forward: CCGCAAATTTCTAGCGACTC; M5 reverse: TCGTCCACACGCAAATACAT.
Northern blotting
Overall, 50 μg total RNA samples were analyzed by northern hybridization. The in vitro transcription was carried out with T7-RNA polymerase (Fermentas) on PCR fragments predicted by bioinformatics, which were constructed to contain the T7 promoter as a 100-nt marker. To prepare the hybridization probe, the L10-Leader gene was labeled with [α-32P] using Random Primer DNA Labeling Kit Ver. 2 (TaKaRa). sRNA was run on a 2% agarose gel containing 2.2M formaldehyde and transferred to nylon membrane (Hybond). RNA was linked to the membrane under the ultraviolet light for 15 minutes. Hybridizations with strand-specific DNA probe in Perfect HybTM Hybridization Solution (TOYOBO) were carried out according to the hybridization solution manufacturer's instructions; 16S ribosomal RNA (rRNA) was used as a loading control. The experiment was repeated at least 3 times.
Bioinformatic prediction of target mRNAs of L10-Leader target in S. mutans UA159
The genome of S. mutans UA159 (NCBI No.: NC_004350) and L10-Leader sequence were analyzed with web services sTarPicker and RNAPredator. sTarPicker (http://ccb.bmi.ac.cn/starpicker/index.php) is based on complementary base-pairing and energy calculations (Ying et al., 2011). RNAPredator (http://rna.tbi.univie.ac.at/RNApredator2/target_search.cgi) uses a dynamic programming approach to compute putative targets (Eggenhofer et al., 2011).
Results
Bioinformatic identification of sRNAs in the S. mutans UA159 genome
The genome of S. mutans UA159 (NCBI No.: NC_004350) was analyzed with sRNA Predict, sRNASVM, and SIPHT prediction software, and the web page “Oral Pathogens Non-Coding Small RNA Prediction” was also used for the prediction of the sRNAs. From the 4 methods of prediction, the total number of sRNAs was 334, of which 40 sRNAs were predicted by at least 2 of prediction methods, including the software and the web page (Table 1; Fig. 1).
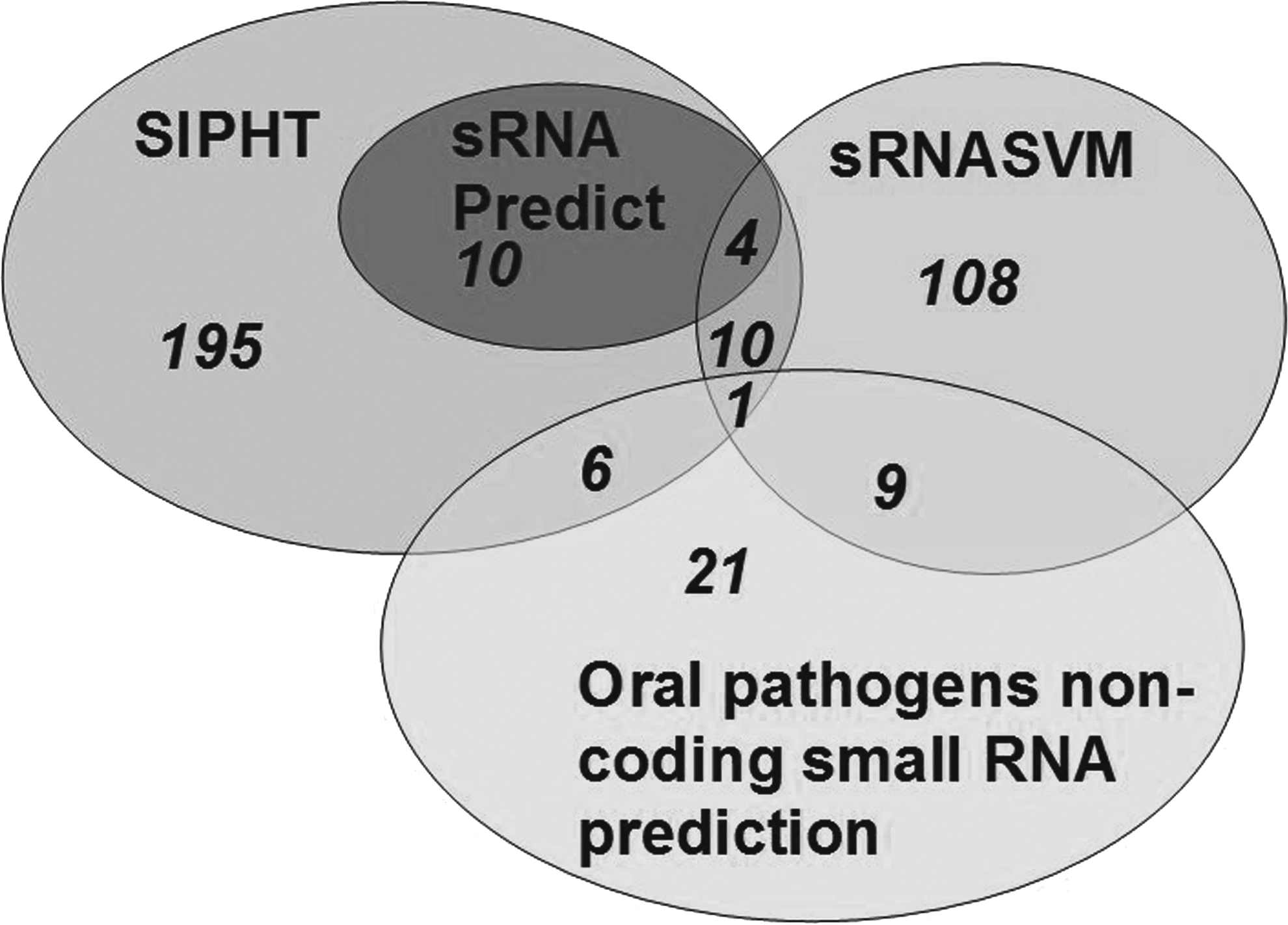
Venn diagram shows the overlap between sRNA Predict, sRNASVM, SIPHT (small non-coding RNA identification protocol using high-throughput technology), and the web page (Oral Pathogens Non-Coding Small RNA Prediction) for the prediction of the small non-coding RNAs (sRNAs). The number of sRNAs is shown in the ellipses and overlapping sections.
Sequence with bold and underline is L10-Leader.
“Oral Pathogens Non-Coding Small RNA Prediction” [Los Alamos National Laboratory Oral Pathogen Sequences Databases (ORALGEN); www.oralgen.lanl.gov/_index.html].
Identification of L10-Leader in S. mutans UA159
We detected all 40 sRNAs predicted by at least 2 software applications by qRT-PCR. The transcript level of L10-Leader was relatively high compared with the other sRNAs and the changes were more significant. Therefore, L10-Leader was chosen for the further study.
The lowest-energy secondary-structure for the L10-Leader sRNA was predicted in S. mutans (Fig. 2a). The S. mutans UA159 was grown to 2 typical phases: mid-logarithmic growth phase (OD600=0.5) and stationary growth phase (OD600=1.5). We detected L10-Leader by northern blotting (Fig. 2b). The northern blot result was consistent with the qRT-PCR result (Fig. 2c). The transcript level of L10-Leader was lower in the stationary growth phase (Fig. 2c).
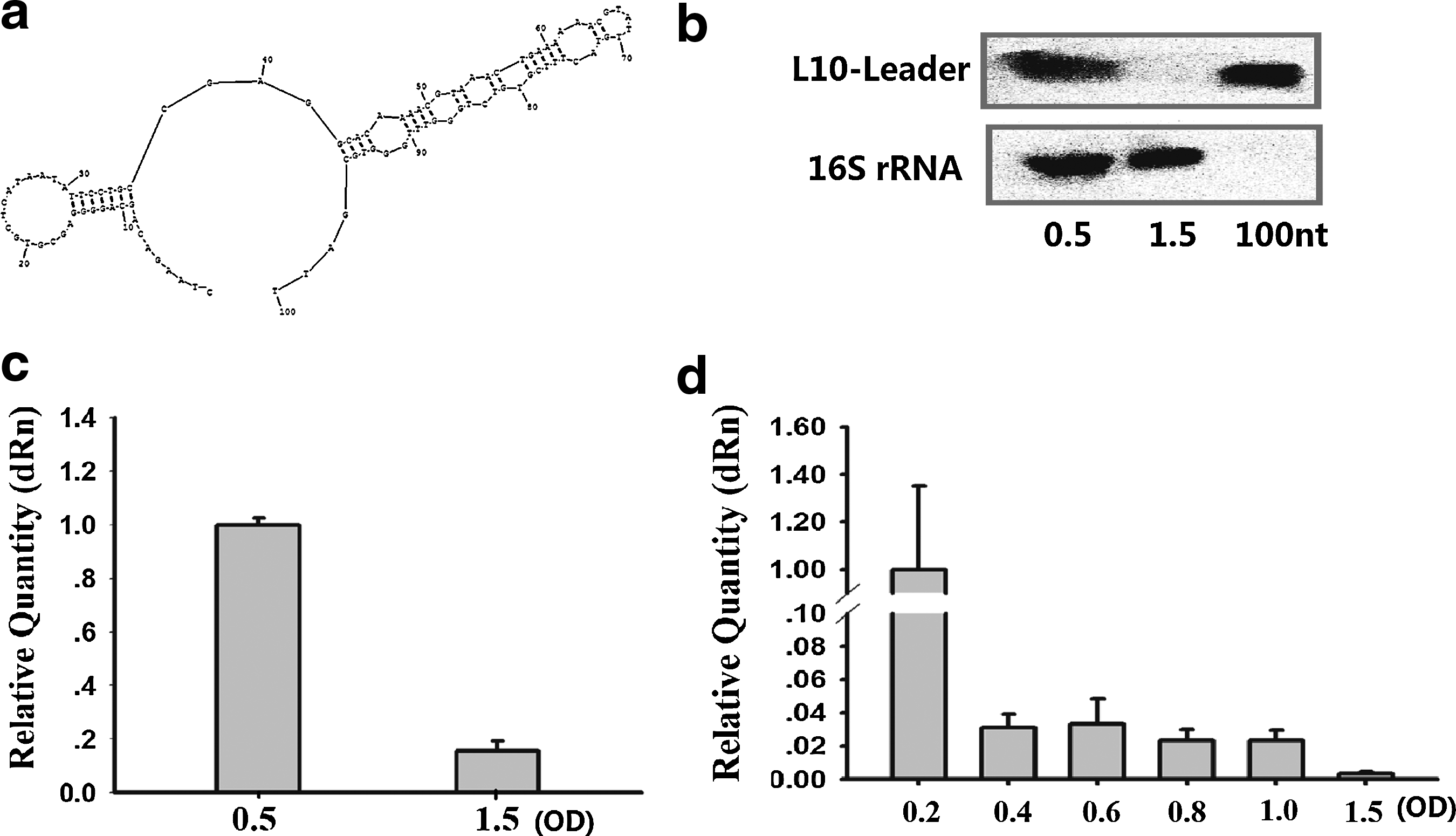
Identification of sRNA L10-Leader in Streptococcus mutans UA159.
S. mutans UA159 was grown in BHI to different growth phases: early-logarithmic growth phase (OD600=0.2), mid-logarithmic growth phase (OD600=0.4, 0.6), late-logarithmic growth phase (OD600=0.8, 1.0), and stationary growth phase (OD600=1.5). The levels of L10-Leader were gradually downregulated from logarithmic growth phase to stationary growth phase (Fig. 2d).
L10-Leader in different clinical strains of S. mutans
Two clinical strains of S. mutans were selected to analyze L10-Leader. Strain 1 has stronger adherence, more synthesis of extracellular polysaccharides, and produces more acid than strain 2. We compared the expression of L10-Leader in these 2 strains at different growth phases: logarithmic growth phase (OD600=0.5) and stationary growth phase (OD600=1.5). The levels of L10-Leader were higher in strain 1 than strain 2, regardless of which stage they were in (Fig. 3a, b). The 2 clinical strains were cultured to early-logarithmic growth phase (OD600=0.2), mid-logarithmic growth phase (OD600=0.5), late-logarithmic growth phase (OD600=1.0), and stationary growth phase (OD600=1.5). Levels of L10-Leader were gradually downregulated from logarithmic growth phase to stationary growth phase (Fig. 3c, d), similar to the effect seen in S. mutans UA159.
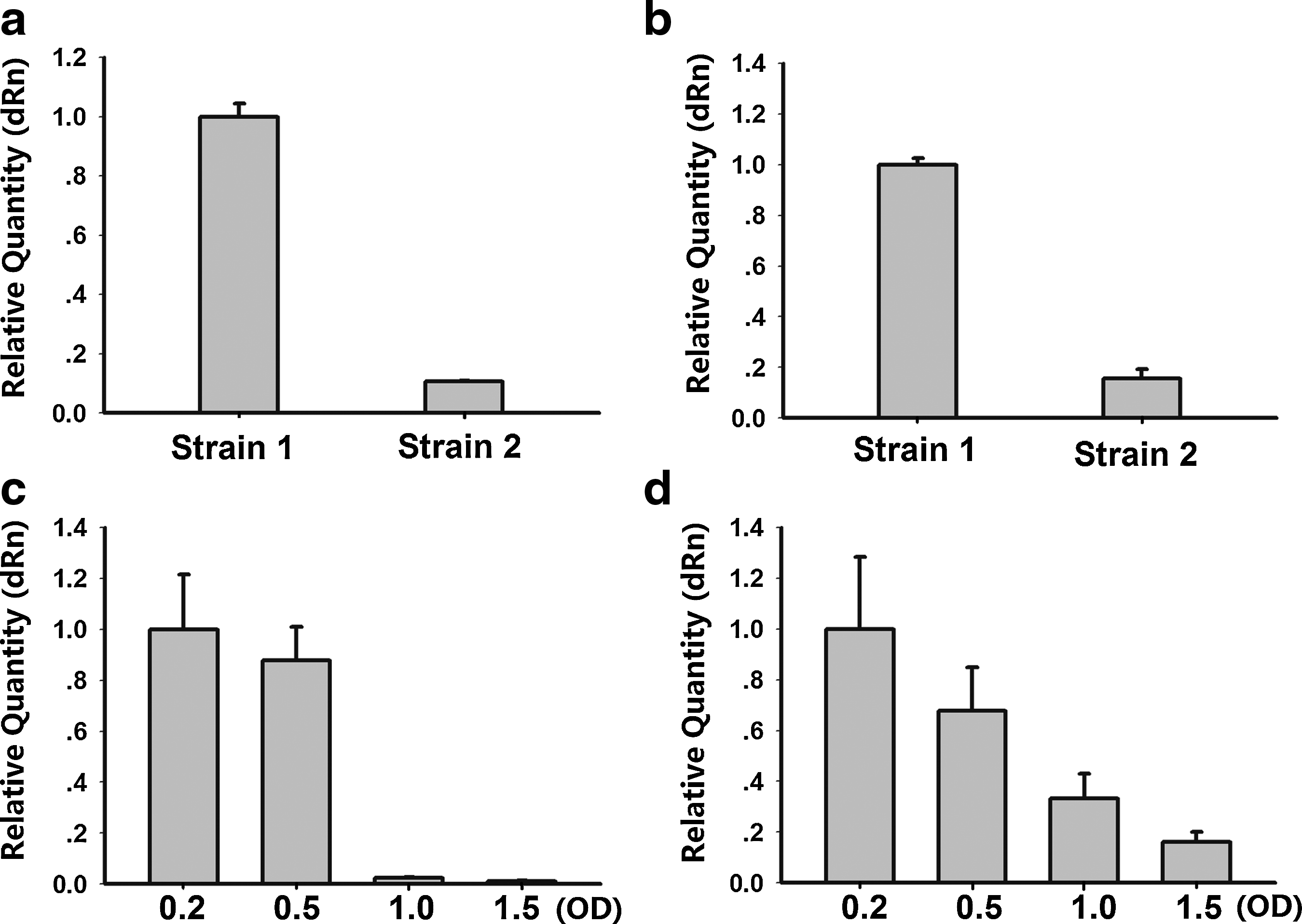
qRT-PCR analysis of L10-Leader expression in clinical strains of S. mutans.
Expression of L10-Leader under different environmental stresses
S. mutans is able to adapt to acidic environments, using glucose for acid production and sucrose to synthesize extracellular polysaccharides for adhesion to the tooth surfaces. The expression of sRNA is related to the environmental stress response, so we examined L10-Leader of S. mutans UA159 under stress conditions. For acid shock assays, S. mutans UA159 were grown in TYE medium at pH 7.5 until cells reached the mid-logarithmic phase (OD600=0.4). Cell cultures were then divided into 2 aliquots, pelleted by centrifugation, and resuspended in TYE medium at pH 5.5 or 7.5. The cells were subsequently incubated for 0.5, 1.5, 2.5, 3.5, or 4.5 hours. The levels of L10-Leader were consistently higher in pH 5.5 than in pH 7.5, from 1.5 to 4.5 hours. There was little change in the level of L10-Leader at 0.5 hours (Fig. 4a).
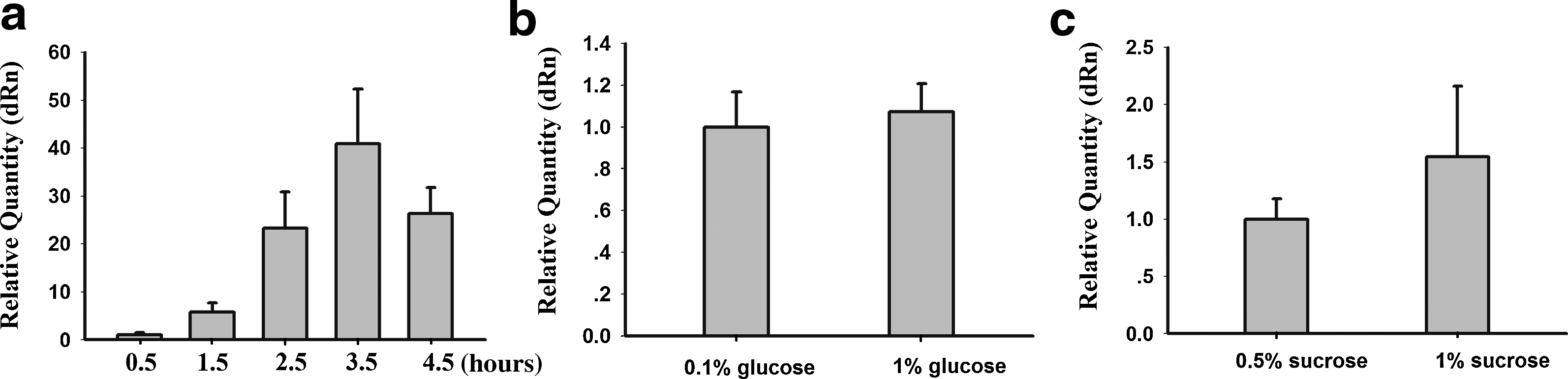
qRT-PCR analysis of L10-Leader expression under different environmental stresses.
When S. mutans UA159 was grown in 0.1% glucose or 1% glucose TYE medium for 24 hours, the levels of L10-Leader showed no significant differences (Fig. 4b). Again, there was no significant change in levels of L10-Leader when the S. mutans strain UA159 was grown in 0.5% sucrose or 1% sucrose TYE medium for 24 hours (Fig. 4c). We propose that L10-Leader has little effect in glucose- and sucrose-starved environments.
Expression of predicted target mRNAs of L10-Leader target in S. mutans UA159
The possible target mRNAs of L10-Leader were predicted according to the bioinformatic prediction web services, sTarPicker and RNAPredator. The numbers of mRNAs predicted by sTarPicker and RNAPredator were 100 and 4946, respectively. The 5 putative targets with highest scores in both prediction web services were chosen by us further investigation. They encode proteins SMU.276c and SMU.688, hydrolase SMU.488, thioesterase SMU.633 and transporter SMU.1149 respectively. L10-Leader and target sites of 5 target mRNAs predicted by sTarPicker (Fig. 5a). The results of qRT-PCR show that the transcription levels of mRNAs were lower in the stationary growth phase than in the logarithmic growth phase (Fig. 5b). These changes are similar to those of L10-Leader. For acid shock assays, S. mutans UA159 were grown in TYE medium at pH 7.5 until cells reached the mid-logarithmic phase (OD600=0.4). Cell cultures were then divided into 2 aliquots, pelleted by centrifugation, and resuspended in TYE medium at pH 5.5 or 7.5. The cells were subsequently incubated for 0.5, 1.5, 2.5, 3.5, or 4.5 hours. The levels of 5 target mRNAs changed differently in pH 5.5 than in pH 7.5, from 0.5 hours to 2.5 hours. But the levels of 5 target mRNAs were consistently higher in pH 5.5 than in pH 7.5, at 3.5 hours and 4.5 hours (Fig. 5c–g), which were similar to L10-Leader. The changes of mRNAs were smaller and slower than L10-Leader.

Target sites and qRT-PCR of L10-Leader 5 target mRNAs.
Discussion
In this study, we used 4 kinds of bioinformatic approaches to predict the sRNAs in S. mutans. Bioinformatic approaches have been developed to identify sRNA-encoding genes in intergenic regions of bacterial genomes by searching for the co-localization of genetic features. These include predicted Rho-independent transcription terminators, promoters and transcription factor binding sites, intergenic conservation among closely related species, and conserved secondary structures (Kulkarni and KULKARNI, 2007). While these approaches have proven to be effective in identifying sRNAs in several species, they present significant computational challenges, requiring the positional relationships of thousands of individual genetic features to be found for each genome-wide search. SIPHT and sRNAPredict software predict candidate sRNA-encoding loci, identified based on the presence of putative Rho-independent terminators downstream of conserved intergenic sequences, and each locus is annotated for several features, including conservation in other species, association with one of several transcription factor binding sites and homology to any of over 300 previously identified sRNAs and cis-regulatory RNA elements (LIVNY, 2007; Livny et al., 2008). sRNASVM is a model for the prediction of small non-coding RNAs in E. coli using support vector machines (Saha and Raghava, 2006). These programs are set up using the features of sRNAs that have been found, and so it is hard to detect novel sRNAs that are completely different from known sRNAs. Some research has used genomic tiling microarrays to detect sRNAs in bacteria (Kumar et al., 2010). The advantage of this method is that it will find all transcripted sRNAs, but sRNA is related to the environment, so more specific experiments are required to find stress-related sRNAs. Furthermore, tiling microarrays require copious amounts of computational analysis of the detected sequence of sRNA. We know from research in S. pyogenes that only 7 of 75 candidate sRNAs were identified by both tiling microarrays and bioinformatic methods (Perez et al., 2009). Regardless of whether bioinformatic prediction or tiling microarrays are used to analyze sRNA, they need to be verified experimentally.
In this study, we first verified the existence of sRNA in S. mutans using experimental approaches. The sequence of sRNA we studied was annotated as L10-leader (RF00557) family in Rfam website. L10-Leader family is a putative ribosomal protein leader auto-regulatory structure found in Bacillus subtilis and other low-guanine-cytosine content Gram-positive bacteria. It has also been identified in L. monocytogenes (Toledo-Arana et al., 2009). It is located in the 5′-UTR of mRNAs encoding ribosomal proteins L10 and L12 (rplJ-rplL). A Rho-independent transcription terminator structure that is probably involved in regulation is included at the 3′ end (Zengel and Lindahl, 1994). The lowest-energy secondary-structure prediction for the L10-Leader sRNA was identified for S. mutans, and like most sRNAs, it has a loop structure. L10-Leader was also detected in clinical strains of S. mutans. The sequence was conserved among Streptococci, such as Streptococcus dysgalactiae subsp. Equisimilis, Streptococcus equi subsp. Zooepidemicus, S. pyogenes, and Streptococcus parauberis. However, in clinical strains, the levels of L10-Leader are different. There were also some sRNAs expressed differently in different strains, which is similar to results found in other studies. In S. aureus, strains of RN6390, Newman, and COL, transcriptional analyses showed different transcription profiles of 11 sRNAs between the different strains (Geissmann et al., 2009). In S. pyogenes, sRNAs PEL, FASX, and SR195750 are variably expressed in different strains. In contrast, 5 candidate sRNAs do not show obviously change in S. pyogenes (Perez et al., 2009). sRNAs regulate the virulence of bacteria and different stains have different features, so different levels of some sRNAs may suggest they are related to virulence, while the others may not be.
The levels of L10-Leader were gradually downregulated from logarithmic growth phase to stationary growth phase, regardless of whether it was in S. mutans UA159 or the clinical strains. Numerous sRNAs in Gram-positive pathogens are growth phase dependent. In L. monocytogenes, there are 3 growth phase dependent sRNAs, which vary in response to growth media (Christiansen et al., 2004). Expression of most of the 11 sRNAs in S. aureus was dependent on the growth phage. In S. pyogenes, several sRNAs changed in different growth phases (Perez et al., 2009). It is well known that there are drastic morphological and physiological properties changes during different bacterial growth stages, from the exponential to the stationary phase. In exponentially growing cells, translation is the most energetically expensive process; its efficiency is under important selective pressure. Under some physiological conditions, a small set of genes regulate the large majority of transcription and translation taking place in the cell (Andersson and Kurland, 1990). These sets include genes related to translation, transcription, and energy metabolism, and are under strong selective pressure for translation efficiency. In S. mutans, htrA and vicRKX genes were shown to be related to growth phase (Ahn et al., 2005; Tremblay et al., 2009).
Our predicted target mRNAs of L10-Leader included putative hydrolase (SMU.488), putative thioesterase (SMU.633), and transporter (SMU.1149). Hydrolase, thioesterase, and transporters are related to the metabolic activity of bacteria. L10-Leader, in combination with these mRNAs, may be involved in the regulation of metabolism, cell growth, and division. For example, hydrolase may be involved in the adenosine triphosphate (ATP) hydrolysis reaction, releasing energy. Thioesterase catalyzes fatty chain termination, releasing free fatty acids, which break down to produce a lot of energy (WAKIL, 1989). These reactions are able to supply sufficient energy for physical activity. Transporters are specialized transmembrane proteins, which recognize the substance and allow it to cross the membrane. The proteins involved are usually pumps, and normally use the chemical energy of ATP and transfer particles.
The levels of L10-Leader in S. mutans UA159 varied in response to low pH growth medium. The ability of S. mutans to tolerate low pH has received considerable attention because it is this property that contributes substantially to the virulence of this cariogenic organism. In one study (Fozo and Quivey, 2004a), it was demonstrated that S. mutans must make major changes to its membrane to survive at low pH, and the action of an enzyme named FabM is required. In fact, changes of cell membrane fluidity by adjustment of the membrane fatty acid composition play an important role in the acid resistance phenotype of S. mutans, which has been shown to influence the virulence of this organism (Fozo and Quivey, 2004b; Fozo et al., 2007). More specifically, studies have shown that at low pH, changes in the membrane fatty acid composition involve a higher proportion of long-chain, monounsaturated fatty acids. This results in a membrane that is less permeable to protons, thereby facilitating survival in highly acidic environments (Fozo et al., 2004). In a study that examined the fatty acid profiles of S. mutans, Streptococcus gordonii, Streptococcus salivarius, and Lactobacillus casei in response to environmental acidification, it was shown that aciduric oral bacteria were able to increase their levels of long-chain, monounsaturated membrane fatty acids at low pH (Fozo and Quivey, 2004b). In S. mutans, the loss of VicK significantly enhanced its survival in an acid tolerance response assay (Senadheera et al., 2009). These observations suggest that membrane fatty acid alteration is a common mechanism utilized by bacteria to withstand environmental stress.
Alterations in the expression of mRNA in S. mutans in acidic environments were analyzed by microarray analysis. Results showed that the expression of most stress response and DNA repair related genes increased (Fozo et al., 2007). Bioinformatic predictions of L10-Leader target mRNAs show related genes, such as the putative manganese-type superoxide dismutase Fe/Mn-SOD (sod SMU.629) and putative DNA mismatch repair protein MutS2 (SMU.1870). An anaerobic bacterium, Propionibacterium shermanii, synthesized either Fe- or Mn-SOD with an identical protein moiety, depending on the metal supplied (Meier et al., 1982). The presence of Fe-containing Mn-SOD in E. coli grown in tryptic soy yeast extract medium suggested the possibility that Fe-substituted Mn-SOD could have a physiological function (Beyer and Fridovich, 1991). Even if a Mn-SOD had been classified as having highly metal-specific enzyme activity, it may also exhibit at least some activity with iron at an acidic pH. The increased expression of L10-Leader in the acidic environment may suggest it combines with the target mRNA to increase its stability and promote the translation of target mRNA into protein. This allows bacteria to adapt to the acidic environment and ensures the correct matching of DNA under stress conditions. However, the role of the L10-Leader of S. mutans under acid conditions remains to be investigated.
We choose 5 possible target mRNAs of L10-Leader according to bioinformatics prediction web services sTarPicker (Ying et al., 2011) (http://ccb.bmi.ac.cn/starpicker/index.php) and RNAPredator (Eggenhofer et al., 2011) (http://rna.tbi.univie.ac.at/RNApredator2/target_search.cgi). The 5 target mRNAs encoded hypothetical proteins (SMU.276c), putative hydrolase (SMU.488), putative thioesterase (SMU.633), transporter (SMU.1149), and hypothetical protein (SMU.688). S. mutans UA159 was grown to logarithmic growth phase and stationary growth phase. The results of qRT-PCR showed the transcript level of mRNAs was lower in stationary growth phase than in logarithmic growth phase. These changes are similar to that seen for L10-Leader. L10-Leader may combine with the target mRNAs to increase their stability, promoting mRNA translation. The proteins could provide particles and energy for the growth of bacteria. For acid shock assays, the levels of 5 target mRNAs changed differently in pH 5.5 than in pH 7.5, from 0.5 hours to 2.5 hours. But interestingly, the levels of 5 target mRNAs were consistently higher in pH 5.5 than in pH 7.5, at 3.5 hours and 4.5 hours. The changes at 3.5 hours and 4.5hours are similar to L10-Leader. The changes of mRNAs are smaller and slower than L10-Leader. As we know, sRNA can response quickly to a variable environment and then regulate mRNAs. An mRNA may be regulated by more than 1 sRNA. So the final changes of mRNAs are very complicated. According to our predictions, many of the potential mRNA targets of L10-Leader are related to bacterial growth and metabolism, such as 30S ribosomal protein S6 s6 – rpsF (SMU.1860), putative citrate lyase alpha subunit cilA (SMU.1021), peptide chain release factor 1 prfA (SMU.1085), and permease protein psaB (SMU.1928). Therefore, we suggest that L10-Leader and its predicted mRNA targets may play important roles in the regulation of growth phase and acid adaption in S. mutans.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 81041028 in 2011 and No. 30772426 in 2008). We are grateful to all the members in our laboratory for their help.
Author Disclosure Statement
No competing financial interests exist.