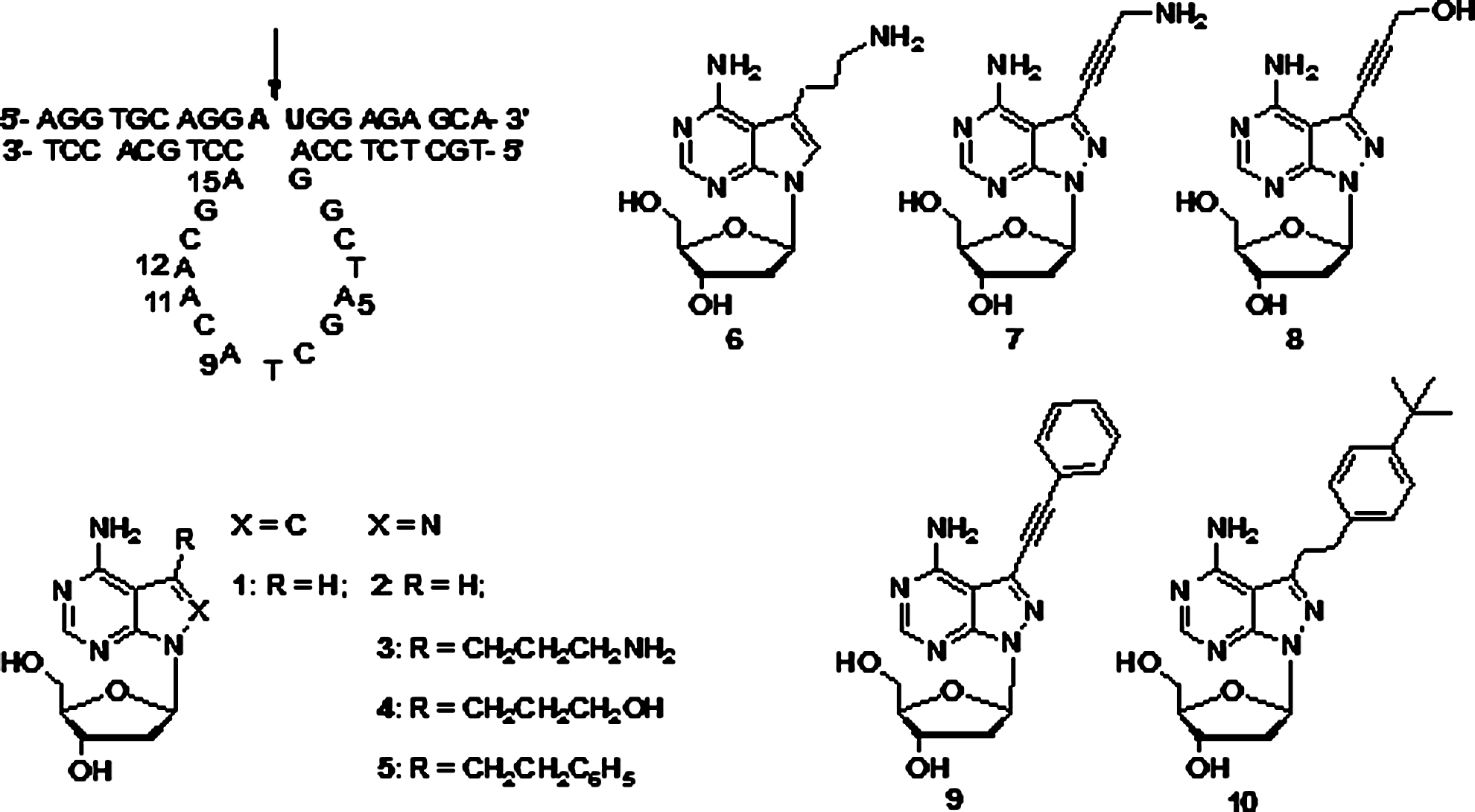
Modification on the catalytic core of 10-23 DNAzyme with protein-like functional groups is a potential approach to obtain its more efficient analogs. In our efforts for this purpose, a lead structure (DZ-2-9) with 8-aza-7-deaza-2′-deoxyadenosine at the A9 position in its catalytic core was obtained. Here we report our structure–activity relationship studies on this lead structure. Various functional groups of different chemical properties were introduced through the 7-substituents of 8-aza-7-deaza-2′-deoxyadenosine to DZ-2-9. The functional groups capable of forming hydrogen bonds, like amino and hydroxyl groups, are more favorable for catalytic rate enhancement than the large groups with spacial occupation, like phenyl and tert-butylphenyl groups, and the flexible alkyl linkage was the more preferred choice for optimizing their positive effect. Furthermore, they exerted positive effect cooperatively with the N8 atom. These results give us a clear hint in the design of compounds for A9 substitution of 10-23 DNAzyme for more efficient DNAzymes.
Introduction
10-23 DNAzyme is a small artificial catalytic DNA molecule consisting of a conserved 15-mer catalytic core and two recognition arms of 7- to 9-mer length (Santoro and Joyce, 1997; Santoro and Joyce, 1998. Its ability to specifically cleave virtually any complementary mRNA in purine-pyrimidine junctions has been demonstrated to be of great potential applications, ranging from therapeutic agents to diagnostic tools (Zhang et al., 2002; SUN, 2005; Bhindi et al., 2007; Benson et al., 2008; Dass et al., 2008; Hengsbach et al., 2008; Chan et al., 2009). Extensive modifications were thus conducted on this structure for further improvement of the properties to meet its practical usages. Its biostability against nucleolytic degradation was significantly enhanced by modification on the two recognition arms with 3′-3′-linked thymidylate, phosphorothioate linkage, 2′-O-methyl RNA, and locked nucleic acid (LNA) (Dass et al., 2002; Schubert et al., 2003). Particularly, several positions in the catalytic core were substituted with corresponding 2′-O-methyl RNA monomers to obtain its stability enhancement against endonucleases. In addition, the incorporation of LNA (Vester et al., 2002) and guanidinium-modified residues into DNAzymes (LAM, 2010) could improve target affinity for rate enhancement and even accessibility for more target choices (Schubert et al., 2004).
However, one of the more challenging objectives of chemical modifications on 10-23 DNAzyme is to establish the role of each functional group of the 15 residues in the catalytic core in order to find a rational approach to improve its catalytic efficiency and understand the catalytic mechanism. Currently, in the absence of information about the catalytic conformation and the active site, the folding of DNAzyme has been investigated with fluorescence resonance energy transfer (FRET), circular dichroism, and surface plasmon resonance (Cieslak et al., 2003). Conformational interruption studies have been conducted through modifications on the sugar-phosphate with replacement of acyclic spacer (Wang et al., 2010), LNA (Wengel et al., 2003), (2′S) or (2′R)-2′-C-methyl-2′-deoxyuridine (Robaldo et al., 2010), or intercalation of large hydrophobic groups at the boundary site between the catalytic core and the binding arms (Asanuma et al., 2006). Generally, the freedom of the sugar moiety should not be discounted. Stereodefined phosphorothioate modification on scissile site of the substrate and its systematic replacement in the catalytic core were used to study the roles of the two non-bridging oxygen atoms and divalent metal ion (Nawrot et al., 2007; Nawrot et al., 2008). The conservation of the fifteen nucleobases and even their functional groups has been evaluated by residue replacement or deletion (Zaborowska et al., 2002; Okumoto et al., 2003; Zaborowska et al., 2005); however, their specific roles have not yet been demonstrated clearly. All of these functional groups are supposed to form multiple interactions, contributing to the catalytic conformation and/or possibly constituting the catalytic site. On the other hand, the lgKa (pKa) values of natural nucleobases (<4.5 or >9) appear to be not sufficient as acid-base catalysts at neutral pH. In the case of 10-23 DNAzyme, it has not been reported that these pKa values might shift close to neutral pH as suggested for ribozymes (Oyelere et al., 2002; Liu et al., 2009). Thus, protein-like functional groups like imidazolyl, amino, amido groups, which are known to be capable of proton transfer in the catalytic reactions of nucleases, have been introduced to the catalytic core to expect the same functions as they display in the catalytic reactions of protein enzymes (Hollenstein et al., 2009; Thomas et al., 2009; Smuga et al., 2010). However, the positive roles of these functional groups were seldom fully explored, because the most appropriate position for their expected roles remains to be determined.
Our recent research for more efficient 10-23 DNAzyme analogs with 2′-deoxyadenosine analogs 1-5 (Fig. 1) revealed that extra protein-like functional groups at A9 position in the catalytic core always resulted in a rate enhancement for the cleavage reaction and DZ-2-9 was hypothesized to be the lead structure (He et al., 2011). Based on these results, we continued our investigation on the identification of the lead structure and the possible structure-activity relationship about it. Compounds 6-10 (Fig. 1) were designed to scrutinize the importance of N8 atom and the properties of the 7-substituents of compound 1 or 2 needed in the design for more efficient 10-23 DNAzyme analogs.
The complex of 10-23 DNAzyme and its DNA–RNA–DNA substrate, in which non-bold letters represent DNA residues and bold letters represent the RNA residues and the arrow denotes the cleavage site on the substrate. Compounds 1–10 were 2′-deoxyadenosine (dA) analogs used for replacement of dA residues in the catalytic core of 10-23 DNAzyme. The DNAzyme with compound 2 at A9 (DZ-2-9) is recognized as the lead structure for further modifications.
Materials and Methods
Oligodeoxyribonucleotides
The synthesis of single-site modified 10-23 DNAzyme analogs were conducted with phosphoramidite chemistry and performed on an Applied Biosystems 392 synthesizer with a 1-μmol scale. The phosphoramidites of modified nucleoside analogs 6 and 10 (see Supplementary Data, available online at www.liebertpub.com/nat), 7 (He et al., 2011), 8 (Zhang et al., 2009), and 9 (Seela et al., 1999) were used in 0.15–0.2 M in acetonitrile, and the coupling time of 3 minutes was applied. The 4,4′-dimethoxytriphenylmethyl (DMT)-off oligodeoxyribonucleotides were cleaved from controlled pore glass (CPG) and deprotected by concentrated aqueous ammonia overnight at 55°C and purified by denaturing polyacrylamide gel electrophoresis (20% polyacrylamide/7 M urea). Desaltation was realized using SEP-PAK cartridges (Oasis MAX, C18, Waters) with repeated washing with sterilized doubly distilled water. The product was lyophilized and stored at −30°C. Characterization of the synthesized DNAzymes was conducted with Matrix-Assisted Laser Desorption Ionization Time of Flight Mass Spectrum (MALDI-TOF MS) performed on an autoflexTOF/TOF (Bruker Daltonics) with 2′, 4′, 6′-trihydroxyacetophenone as the matrix (Supplementary Table S1).
Cleavage reaction under single-turnover conditions
The cleavage reactions were conducted under single-turnover conditions, with deoxyribozyme (2 μM) in large excess over the 32P-5′-end labeled DNA–RNA–DNA substrate (20 nM) in a buffer of 50 mM Tris-HCl (pH 7.4) containing Mg2+(Santoro and Joyce, 1997; He et al., 2011). According to the initial screening results, 2 mM Mg2+ was used in order to conduct an accurate manual measurement. Different time intervals were adopted for monitoring the reaction process of different 10-23 DNAzyme analogs. The deoxyribozyme and the substrate were mixed in a buffer of 50 mM Tris-HCl; after incubation at 90°C for 3 minutes, the reaction was initiated by addition of Mg2+ in Tris-HCl to the final concentration of 2 mM in the reaction buffer. The reaction process was monitored by sampling at appropriate time intervals followed by immediate quenching with stopping solution (100 mM EDTA, 8 M Urea). The cleaved products and substrate were separated by electrophoresis on 20% polyacrylamide/7 M urea denaturing gel and quantitated by a Storm 840 PhosphorImager (Molecular Dynamics). The first-order rate constants (kobs) were calculated according to the equation P%=P∞% – C·exp [–kobst], where P is the percentage of the cleaved product at time t, C is the difference in P% between t=∞ and t=0, and P∞ is the endpoint, assuming an endpoint of 90%. The data was the averaged result of at least 3 independent experiments with less than 20% variation.
Results and Discussion
The cooperation of N8 atom and 7-substituents in the lead structure DZ-2-9 for rate enhancement
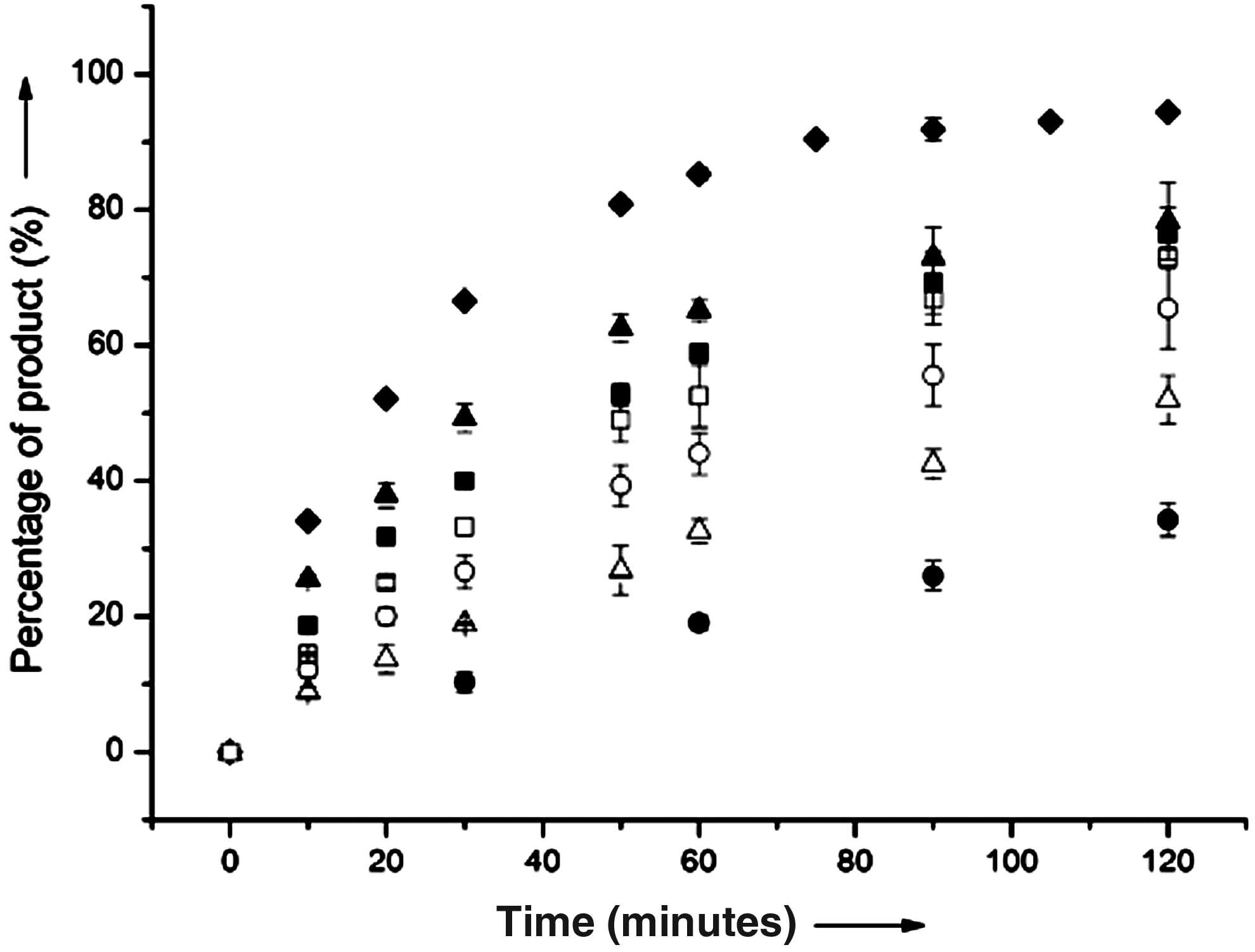
Compounds 2–5 at A9 of 10-23 DNAzyme (He et al., 2011) conducted positive effect on the cleavage reaction through their N8 atom and the 7-substituents. We wondered if N8 atom and the 7-substituents in compounds 3–5 contribute to the reaction additively; compound 6 was thus designed and incorporated at the A9 position in DZ-6-9 (Table 1; Fig. 2), in which only 7-aminopropyl remained. Compared with 10-23 DNAzyzme, a 7-aminopropyl group of compound 6 in DZ-6-9 contributed about 4.5-fold rate increase (kobs=0.0167±0.0012 min–1), but its combination with N8 atom in compound 3 obtained a 12-fold increase for DZ-3-9 (kobs=0.045±0.004 min−1), which was also greater than the addition of both DZ-2-9 and DZ-6-9. Such a difference of rate increase indicated that the N8 atom and 7-aminopropyl group of compound 3 contributed to the reaction cooperatively. This result also demonstrated that compound 2 rather than compound 1 should be recognized as the lead compound for the A9 position in future chemical modifications.
The cleavage pattern of DNAzymes modified with compounds 3 or 6–10 at A9 within 120 minutes: 3 in DZ-3-9 (solid diamond), 10-23 DNAzyme (solid circle), 6 in DZ-6-9 (solid square), 7 in DZ-7-9 (solid triangle), 8 in DZ-8-9 (open circle), 9 in DZ-9-9 (open triangle), and 10 in DZ-10-9 (open square).
kobsof 10-23 DNAzyme Analogs with a Single Substitution at A9 by 2′-Deoxyadenosine Analogs 6, 7, 8, 9, or 10 in the Catalytic Core Measured Under Single-Turnover Conditions
Name
10–23 DNAzyme analogs
kobs (min–1)
10–23 DNAzyme
5′-d(tgc tct cca GGC TAG CTA CAA CGA cct gca cct)-3′
In the sequences of the DNAzymes, small letters constitute the 2 recognition arms, and the capital letters represent the catalytic core.
The effect of the linkage in 7-subsituents of compound 2 on rate enhancement
In the cleavage reactions conducted by modified DNAzymes DZ-3-9 and DZ-4-9, the incorporation of compounds 3 and 4 at their A9 position led to the most positive effect on the cleavage rate, respectively (He et al., 2011). It was speculated that the extra amino and hydroxyl groups in the specific position could drive a more favorable change of the catalytic conformation through hydrogen binding with other functional groups within the catalytic core. It is reasonable that the spacial location of these functional groups is very crucial for their positive role in the cleavage reaction. When their flexible linker was replaced by a rigid propynyl linker, as shown with compounds 7 and 8, respectively, the new deoxyribozyme DZ-7-9 (kobs=0.024±0.002 min–1) was found to be slower than DZ-3-9, and DZ-8-9 (kobs=0.0111±0.0015 min–1) was slower than DZ-4-9 (kobs=0.025±0.006 min–1), too (Table 1). Such a rate decrease reflected the importance of the linkage, because it decides the location of the amino and hydroxyl groups in the context of the catalytic core. The flexible linkage probably permits more freedom for amino and hydroxyl groups to adapt to the formation of a more favorable catalytic conformation. Similarly, when the phenyl group was restricted by a rigid linker in compound 9, its incorporation at A9 (DZ-9-9) (Table 1) also resulted in a less positive effect than compound 5 (DZ-5-9, kobs=0.0128±0.0018 min–1). These facts verified the importance of the spacial location of the extra functional groups in the catalytic core of 10-23 DNAzyme for an expected positive effect. In the present modification approach, the location of extra functional groups was thus defined by 7-substituents of 8-aza-7-deaza-2′-deoxyadenosine located at A9 for their positive roles. It is notable that the contribution of the 7-substituted functional groups by rigid linkage is in the order of NH2>OH>phenyl, just like their behavior when linked with the flexible alkyl linker in compounds 3–5.
The positive effect of large hydrophobic 7-substituents of compound 2 at the A9 position
It has been demonstrated that the phenyl group of compounds 5 and 9 at the A9 position in the catalytic core of 10-23 DNAzyme could play a positive role, although its effect was not as evident as that of an amino or hydroxyl group (He et al., 2011). This phenyl group was supposed to exert its role by hydrophobic interactions and/or spacial clash, leading to a favorable conformational change during the DNAzyme–substrate complex formation. For a further confirmation of such speculation, compound 10 with a 7-positioned 4-tert-butylphenyl group was designed and incorporated into the A9 position of 10-23 DNAzyme, the new DNAzyme DZ-10-9 was obtained (Table 1; Fig. 2). As expected, DZ-10-9 (kobs=0.0153±0.0017 min–1) exhibited a tendency toward more efficient cleavage compared with DZ-5-9. Taken together, these observations confirmed that a large hydrophobic group, when introduced by compound 2 at A9 position, as shown by compounds 5, 9, and 10, could induce a favorable change toward the active conformation, and its location could be more appropriately positioned by a flexible linkage.
The unique A9 position for the positive effect of 7-substituents further examined with compounds 6-8
As mentioned above, the effect of the 7-substituents was strongly related to the linkage used. In order to find whether the propynyl-linked functional groups behave with the same position-dependence in the catalytic reaction of 10-23 DNAzyme analogs, further position-dependent effects of compounds 6-8 were investigated. For the amino group with rigid propynyl linker in compound 7, when incorporated in the other 4 positions of 2′-deoxyadenosine (dA) in the catalytic core (A5, A11, A12, and A15), a negative effect was always observed (Table 2; Fig. 3), except for A12 (DZ-7-12). The extent of the negative effect of compound 7 in these four positions is a little different from that observed for compound 3, which might reflect the different influences of the linkage. But their most positive effect was always produced when located at the A9 position. The similar position-dependent effect of 7-aminopropynyl and 7-aminopropyl was observed. A negative effect was also observed for compounds 6 and 8 when located at A5 and A15 (Table 2). Based on the present chemical modification approach on 10-23 DNAzyme, A9 is the unique position for the most positive effect of all the functional groups introduced by 7-substituted 8-aza-7-deaza-2′-deoxyadenosine.
The cleavage pattern of DNAzymes modified with compound 7 at different positions: A5 (open triangle), A9 (solid triangle), A11 (solid circle), A12 (open circle), and A15 (open diamond) within 9 hours.
Random Modifications on 10-23 DNAzyme with 2′-Deoxyadenosine Analogs 6, 7, and 8 Characterized by First-Order Rate Constants Under Single-Turnover Conditions
5′-d(tgc tct cca GGC TAG CTA CAA CG8 cct gca cct)-3′
nd
In the sequences of the DNAzymes, small letters constitute the 2 recognition arms, and the capital letters represent the catalytic core.
kobs, first-order rate constants; nd, not detected under present conditions.
Conclusions
In the catalytic core of 10-23 DNAzyme, all of the residues and their specific positions have been demonstrated to be conservative for the catalytic activity by residue replacement and deletion, except that T8 can tolerate other canonical or modified nucleosides without much influence on the catalytic behavior. Considering that multiple functional groups of each residue and their complex interactions, specific chemical modifications on the 5 dA residues with dA analogs, was demonstrated to be a feasible approach for screening both position and compound for the most positive effect. Of all the deoxyadenosine analogs used for 5 dA positions in the catalytic core of 10-23 DNAzyme, the 7-substituted functional groups, including amino, hydroxyl, phenyl, and p-tert-butyl-phenyl, of 8-aza-7-deaza-2′-deoxyadenosine play the greatest positive role when located at the A9 position. Their contribution is in the order of amino>hydroxyl>p-tert-butyl-phenyl>phenyl, and the amino group exerts the most positive effect, probably because its most powerful hydrogen-bonding ability results in the most favorable conformational change. In each case, the flexible linkage is always more favorable for the positive role than the rigid alkynyl linkage. The present lead structure identification and structure-activity relationship research implied that the functional groups of large volume and strong hydrogen-bonding ability should be the choice for future design of the dA analogs for A9 position. The other 3 kinds of natural residues are also worthwhile to be modified with their respective analogs for more efficient DNAzymes. This research demonstrated that the protein-like functional groups can be used to improve the performance of functional nucleic acids, but the subtle spacial position of the extra functional groups is crucial and must be deliberately modulated for a positive effect.
Footnotes
Acknowledgments
We are grateful for the financial support from the National Key Technologies R&D Program for New Drugs 2009ZX09301-002 and the National Natural Science Foundation of China 21072229.
Author Disclosure Statement
No competing financial interests exist.
References
1.
ASANUMAH., HAYASHIH., ZHAOJ., LIANGX., YAMAZAWAA., KURAMOCHIT., MATSUNAGAD., AIBAY., KASHIDAH., KOMIYAMAM.2006. Enhancement of RNA cleavage activity of 10-23 DNAzyme by covalently introduced intercalator. Chem. Commun., 5062–5064.
2.
BENSONV.L., KHACHIGIANL.M., LOWEH.C.2008. DNAzymes and cardiovascular disease. Br. J. Pharm., 154:741–748.
3.
BHINDIR., FAHMYR.G., LOWEH.C., CHESTERMANC.N., DASSC.R., CAIRNSM.J., SARAVOLACE.G., SUNL.Q., KHACHIGIANL.M.2007. Brothers in arms DNA enzymes, short interfering RNA, and the emerging wave of small-molecule nucleic acid-based gene-silencing strategies. Am. J. Pathol., 171:1079–1088.
4.
CHANC.W.S., KHACHIGIANL.M.2009. DNAzymes and their therapeutic possibilities. Inter. Med. J., 39:249–251.
5.
CIESLAKM., SZYMANSKIJ., ADAMIAKR.W., CIERNIEWSKIC.S.2003. Structural rearrangements of the 10-23 DNAzyme to β3 integrin subunit mRNA induced by cations and their relations to the catalytic activity. J. Biol. Chem., 278:47987–47996.
6.
DASSC.R., CHOONGP.F.M., KHACHIGIANL.M.2008. DNAzyme technology and cancer therapy: cleave and let die. Mol. Cancer Ther., 7:243–251.
7.
DASSC.R., SARAVOLACE.G., LIY., SUNL.Q.2002. Cellular uptake, distribution, and stability of 10-23 deoxyribozymes. Antisense Nucleic Acid Drug Dev., 12:289–299.
8.
HEJ.L., ZHANGD., WANGQ., WEIX., CHENGM.S., LIUK.L.2011. A novel strategy of chemical modification for rate enhancement of 10-23 DNAzyme: a combination of A9 position and 8-aza-7-deaza-2’-deoxyadenosine analogs. Org. Biomol. Chem., 9:5728–5736.
9.
HENGSBACHM., MEUSBURGERM., LYKOF., HELMM.2008. Use of DNAzymes for site-specific analysis of ribonucleotide modifications. RNA, 14:180–187.
10.
HOLLENSTEINM., HIPOLITOC.J., LAMC.H., PERRIND.M.2009. A self-cleaving DNA enzyme modified with amines, guanidines and imidazoles operates independently of divalent metal cations (M2+)Nucleic Acids Res., 37:1638–1649.
11.
LAMC.H., PERRIND.M.2010. Introduction of guanidinium-modified deoxyuridine into the substrate binding regions of DNAzyme 10-23 to enhance target affinity: implications for DNAzyme design. Bioorg. Med. Chem. Lett., 20:5119–5122.
12.
LIUL., COTTRELLJ.W., SCOTTL.G., FEDORM.J.2009. Direct measurement of the ionization state of an essential guanine in the hairpin ribozyme. Nat. Chem. Biol., 5:351–357.
13.
NAWROTB., WIDERAK., SOBCZAKM., WOJCIKM., STECW.J.2008. Effect of RP and SP phosphorothioate substitution at the scissile site on the cleavage activity of deoxyribozyme 10-23. Curr. Org. Chem., 12:1004–1009.
14.
NAWROTB., WIDERAK., WOJCIKM., REBOWSKAB., NOWAKG., STECW.J.2007. Mapping of the functional phosphate groups in the catalytic core of deoxyribozyme 10-23. FEBS J., 274:1062–1072.
15.
OKUMOTOY., TANABEY., SUGIMOTON.2003. Factors that contribute to efficient catalytic activity of a small Ca2+-dependent deoxyribozyme in relation to its RNA cleavage function. Biochemistry, 42:2158–2165.
16.
OYELEREA.K., KARDONJ.R., STROBELS.A.2002. pKa perturbation in genomic hepatitis Delta Virus ribozyme catalysis evidenced by nucleotide analogue interference mapping. Biochemistry, 41:3667–3675.
17.
ROBALDOL., MONTSERRATJ.M., IRIBARRENA.M.2010. 10–23 DNAzyme modified with (2′R)- and (2′S)-2′-deoxy-2′-C-methyluridine in the catalytic core. Bioorg. Med. Chem. Lett., 20:4367–4370.
18.
SANTOROS.W., JOYCEG.F.1997. A general purpose RNA-cleaving DNA enzyme. Proc. Natl. Acad. Sci. U. S. A., 94:4262–4266.
19.
SANTOROS.W., JOYCEG.F.1998. Mechanism and utility of an RNA-cleaving DNA enzyme. Biochemistry, 37:13330–13342.
20.
SCHUBERTS., FÜRSTEJ.P., WERKD., GRUNERTH.P., ZEICHHARDTH., ERDMANNV.A., KURRECKJ.2004. Gaining target access for deoxyribozymes. J. Mol. Biol., 339:355–363.
21.
SCHUBERTS., GÜLD.C., GRUNERTH.P., ZEICHHARDTH., ERDMANNV.A., KURRECKJ.2003. RNA cleaving ‘10-23’ DNAzymes with enhanced stability and activity. Nucleic Acids Res., 31:5982–5992.
22.
SEELAF., ZULAUFM.1999. Oligonucleotides containing 7-deazaadenines: the influence of the 7-substituent chain length and charge on the duplex stability. Helv. Chim. Acta, 82:1878–1898.
23.
SMUGAD., MAJCHRZAKK., SOCHACKAE., NAWROTB.2010. RNA-cleaving 10-23 deoxyribozyme with a single amino acid-like functionality operates without metal ion cofactors. New J. Chem., 34:934–948.
24.
SUNL.Q.2005. Use of deoxyribozymes for gene knockdown. Med. Chem. Rev., 2:81–87.
25.
THOMASJ.M., YOONJ.K., PERRIND.M.2009. Investigation of the catalytic mechanism of a synthetic DNAzyme with protein-like functionality: an RNaseA mimic? J. Am. Chem. Soc., 131:5648–5658.
26.
VESTERB., LUNDBERGL.B., SØRENSENM.D., BABUB.R., DOUTHWAITES., WENGELJ.2002. LNAzymes: incorporation of LNA-type monomers into DNAzymes markedly increases RNA cleavage. J. Am. Chem. Soc., 124:13682–13683.
27.
WANGB., CAOL., CHIUMANW., LIY., XIZ.2010. Probing the function of nucleotides in the catalytic cores of the 8-17 and 10-23 DNAzymes by abasic nucleotide and C3 spacer substitutions. Biochemistry, 49:7553–7562.
ZABOROWSKAŻ., FÜRSTEJ.P., ERDMANNV.A., KURRECKJ.2002. Sequence requirements in the catalytic core of the “10-23” DNA enzyme. J. Biol. Chem., 277:40617–40622.
30.
ZABOROWSKAŻ., SCHUBERTS., KURRECKJ., ERDMANNV.A.2005. Deletion analysis in the catalytic region of the 10-23 DNA enzyme. FEBS Lett., 579:554–558.
31.
ZHANGL., GASPERW.J., STASSS.A., IOFFEO.B., DAVISM.A., MIXSONA.J.2002. Angiogenic inhibition mediated by a DNAzyme that targets vascular endothelial growth factor receptor 2. Cancer Res., 62:5463–5469.
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.