
Abstract
Pseudomonas aeruginosa is an opportunistic pathogen causing severe infections in hospital settings, especially with immune compromised patients, and the increasing prevalence of multidrug resistant strains urges search for new drugs with novel mechanisms of action. In this study we introduce antisense peptide–peptide nucleic acid (PNA) conjugates as antibacterial agents against P. aeruginosa. We have designed and optimized antisense peptide–PNA conjugates targeting the translation initiation region of the ftsZ gene (an essential bacterial gene involved in cell division) or the acpP gene (an essential bacterial gene involved in fatty acid synthesis) of P. aeruginosa (PA01) and characterized these compounds according to their antimicrobial activity and mode of action. Four antisense PNA oligomers conjugated to the H-(R-Ahx-R)4-Ahx-βala or the H-(R-Ahx)6-βala peptide exhibited complete growth inhibition of P. aeruginosa strains PA01, PA14, and LESB58 at 1–2 μM concentrations without any indication of bacterial membrane disruption (even at 20 μM), and resulted in specific reduction of the targeted mRNA levels. One of the four compounds showed clear bactericidal activity while the other significantly reduced bacterial survival. These results open the possibility of development of antisense antibacterials for treatment of Pseudomonas infections.
Introduction
P. aeruginosa possesses a 6.3 Mb genome encoding 5,570 predicted open reading frames and differing dramatically from the human genome. Therefore, it is not difficult to identify antisense sequence targets that are unique for essential genes in P. aeruginosa, and it should be possible to develop novel antibacterial drugs against P. aeruginosa based on RNA interference, drawing on the experience with other bacteria, particularly Escherichia coli. Indeed, it was demonstrated 10 years ago that a peptide nucleic acid (PNA) oligomer targeting the start codon on the acpP gene, an essential bacterial gene involved in fatty acid synthesis, is able to inhibit the growth of the E. coli strain AS19 (a cell wall mutant that is more permeable to drugs), but not of wild type E. coli (Good and Nielsen, 1998). However, when using the same PNA conjugated to the peptide KFFKFFKFFK, which was known to facilitate bacterial uptake of antibiotics, and thus enhance antibiotic activity, antibacterial activity (minimal inhibitory concentration [MIC]≅1 μM) in wild type E. coli was obtained (Good et al., 2001) even in the presence of eukaryotic cells (HeLa cells) which were not affected by the PNA conjugate. Many subsequent studies have confirmed these findings (Dryselius et al., 2003a; Dryselius et al., 2003b; GELLER, 2005; Gruegelsiepe et al., 2006; Nikravesh et al., 2007), and extended them to other gram negative bacteria such as (multiresistant) Klebsiella pneumonia (Kurupati et al., 2007) and also gram positive bacteria, such as Staphylococcus aureus (Nekhotiaeva et al., 2004). Most recently, simple mouse peritonitis model experiments were used to demonstrate that such antibacterial effects may be extended to an in vivo situation (Tan et al., 2005; Bai et al., 2012). Analogously, several studies have demonstrated similar antibacterial activity of PMO (morpholinophosphordiamidate)–peptide conjugates, and new peptide carriers have been identified (Mellbye et al., 2009; Greenberg et al., 2010; Mellbye et al., 2010).
In the present study, we aimed at identifying antisense peptide–PNA conjugates exhibiting potent antibacterial, bactericidal activity against P. aeruginosa as part of the quest for developing novel antibiotics for resistant infections. Unexpectedly, we find that the (KFF)3F peptide, which has shown great promise as antisense agent carrier in both gram-positive and gram-negative bacteria (Good et al., 2001; Nekhotiaeva et al., 2004), is not very efficient in P. aeruginosa. However, anti-acpP or anti-ftsZ PNA oligomers conjugated to the (R-Ahx)6-βala or (R-Ahx-R)4-Ahx-βala peptides resulted in validated antisense conjugates with MIC around 1 μM against 3 P. aeruginosa strains: PA01 (the standard strain), PA14 (a highly virulent clinical isolate), and LESB58 (a highly virulent epidemic strain) (Ibrulj et al., 2008).
Materials and Methods
Bacterial strains, medium, PNAs, and peptide–PNA conjugate synthesis
Pseudomonas aeruginosa strains, PA01, PA14, and LESB58 were obtained from Prof. Miguel Camara (University of Nottingham). Tryptic soy broth (TSB) and Mueller Hinton broth (MHB) were used in all experiments. Peptide–PNA conjugates were synthesized by continuous solid phase synthesis using Boc-chemistry as previously described (Christensen et al., 1995; Good et al., 2001). The conjugates were purified by reversed phase high-performance liquid chromatography (HPLC) on an RP18 column using a 0%–50% acetonitrile gradient in 0.1% trifluoroacetic acid. Characterization in terms of purity and identity was done by HPLC and Matrix-assisted laser desorption/ionization-Time Of Flight (MALDI-TOF) mass spectrometric analyses. All conjugates exhibited masses within experimental error corresponding to the calculated mass and were 90+% pure by HPLC analysis (see Supplementary Tables S1, S2; Supplementary Data are available online at www.liebertpub.com/nat). Gentamicin, ciprofloxacin, ofloxacin, and ceftazidime were purchased from Sigma and tested in similar conditions.
Bacteria were grown overnight in TSB. For growth inhibition experiments, a fresh overnight bacterial culture was diluted in MHB at an inoculation of ∼2.5×104 cells per well in a total volume of 250 μL. All experiments were performed in 96-well plates. Cells were grown at 37°C with continuous orbital shaking (220 rpm) and optical density (at 595 nm) was measured continuously at 21-minute intervals.
Bacterial kill kinetics
A fresh overnight culture of P. aeruginosa PA01 strain was diluted in MHB at ∼ 2.5×104 cells in 250 μL in each well of a 96-well plate. The peptide–PNA conjugate dissolved in 30 μL of water was added at 2×MIC value and the cells were grown for 8 hours at 37°C with continuous orbital shaking (375 rpm). Cells were collected at 0-, 1-, 4-, and 8-hour time points and plated on Luria Broth (LB) agar plates. Colony forming units (CFU) were counted after overnight incubation of the plates at 37°C. All experiments were performed in biological duplicates and untreated bacterial culture was used as a control.
Messenger RNA expression
An overnight culture of PA01 was diluted in MHB and ∼2.5×104 cells were added in each well of a 96-well plate with the desired amount of PNA in a total volume of 250 μL. Condition 1: Cells were grown for 8 hours at 37° C with orbital shaking (220 rpm). Antisense PNA treated cells were harvested at optical density (OD)=0.1–0.2, whereas untreated and mismatch PNA treated cells were harvested at OD=0.4–0.6. Condition 2: An inoculate in MHB from an overnight bacterial culture was grown until OD=0.3–0.4. Then the bacterial culture was diluted to an OD of 0.1 and incubated with PNA for 1 hour.
RNA extraction was performed according to the Pure Link RNA Mini Kit (Ambion) instructions, and bacterial RNA degradation was prevented by RNAprotect Bacteria Reagent (Qiagen). In addition, on column genomic DNA (gDNA) digestion was performed by PureLink™. DNase (Invitrogen) and genomic DNA (gDNA) contamination was checked by polymerase chain reaction (PCR).
Real time-PCR
Maxima™ Reverse Transcriptase Kit (Thermo SCIENTIFIC) was used for cDNA conversion by using gene specific primers (for which the PCR product does not include the PNA binding target): acpP (5′-CCATCGAAGAACGCGTTAAG, 5′-CCTGAACGGTGGTGATCTTT), proC (5′-GGCGTATTTCTTCCTGCTGA, 5′-TGGCCTGGAAGGATTTGAT), rpoD (5′-CTGATCCAGGAAGGCAACAT, 5′-TGAGCTTGTTGATCGTCTCG), 16s rRNA (5′-CGTAAGGGCCATGATGACTT, 5′-AAGCAACGCGAAGAACCTTA), and ftsZ (5′-GCGGTATCTCCGACATCATC, 5′-AGGTTGACGTCTTCCAGCAG). The procedures were according to the instructions of the manufacturer.
cDNA in 2 μL of water was amplified in 10μL of PCR mix by using 2 μl LightCycler FastStart DNA Master PLUS SYBER Green (Roche), 0.1μL (0.5 μM) of each primer and 5.8 μL of water. Reactions were carried out in Light Cycler (Roche) by using the following conditions: An initial activation step at 95° C for 10 minutes, 55 PCR cycles composed of: Denaturation at 95° C for 10 seconds, annealing at 59° C for 5 seconds, and extension at 72° C for 30 seconds. The reading was typically achieved around cycle 30. Each reaction was performed in technical duplicate. The Roche real-time software was used for data analysis and normalization. 16S rRNA was used as a reference (internal control).
Membrane disruption assay
A culture of P. aeruginosa PA01 was grown in MHB (195 rpm at 37°C), until OD595=0.4. Then cells were isolated by centrifugation and washed with 0.9% NaCl. The cells were resuspended in MHB reaching an OD595 of 0.1, and Sytox green dye (Invitrogen) was added to the bacterial culture at a concentration of 1μg/mL and incubated for 5 minutes at 37° C. Sytox treated cells (2×107) were added simultaneously to each well of the 96-well plate containing the desired amount of peptide or peptide–PNA conjugate. Fluorescence intensity (λex=480 nm, λem=520nm) was measured continuously for a 90-minute period at 37° C with continuous shaking. Polymyxin–B (Sigma) was used as a control. All compounds were tested at 20 μM (Roth et al., 1997; Srinivas et al., 2010).
Cell toxicity
Cell toxicity determination was performed using HeLa cells grown in 10% fetal bovine serum supplemented RPMI 1640 medium (Invitrogen). Ten thousand cells were seeded in each well of a 96-well plate. After 24 hours, the antibacterial PNA was added, and incubation was continued overnight. Total amount of adenosine triphosphate (ATP) was used as a measure of cell viability, and the ATP measurement was performed according to the CellTiter-Glo Luminescent Cell Viability Assay (Promega) kit.
Results and Discussion
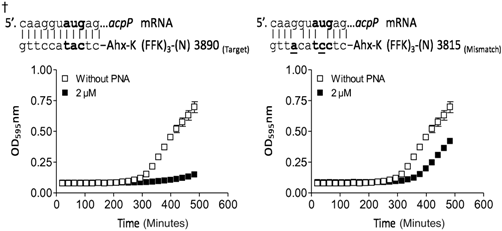
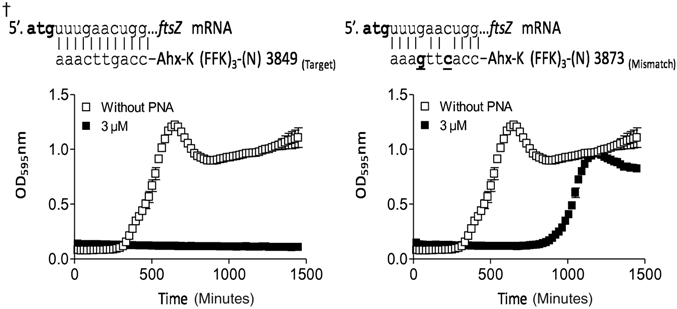
Previous studies have shown that antisense-(KFF)3K–PNA peptide conjugates targeting the acpP and ftsZ gene in E. coli have potent antibacterial activity (Goh et al., 2009; Good et al., 2001). These genes are conserved among gram-positive and gram-negative bacteria and have essential functions in fatty acid synthesis (acpP) and cell division (ftsZ), respectively, and the acpP gene target in particular has been validated for antisense bactericidals (Good and Nielsen, 1998; Good et al, 2001; Mellbye et al., 2009; Mellbye et al., 2010). Accordingly, we initially designed a series of 12 nucleobase anti-acpP and ftsZ PNA oligomers (Tables 1, 2) conjugated to the (KFF)3K peptide, targeting positions in the mRNA around and at the Shine-Dalgarno sequence and translation start site, which in numerous studies were found to be the most sensitive part of the mRNA for antisense targeting in bacteria (Good et al., 2001; Dryselius et al., 2003a) (Fig. 1). These PNA conjugates were tested for antibacterial activity against P. aeruginosa PA01 (Tables 1, 2). Based on antibacterial potency, we selected PNA 3834 (anti-acpP) and PNA 3775 (anti-ftsZ) having MIC values of 4 μM and 3 μM. The targets for these PNAs are located −7 bp upstream to +5 bp downstream from translation initiation on the acpP mRNA and +2 bp to +13 bp downstream on the ftsZ mRNA, respectively (Fig. 1). It is well established that the optimal length of bacterial antisense PNAs and PMOs are typically around 10–12 bases (Good et al., 2001; Mellbye et al., 2009; Greenberg et al., 2010; Mellbye et al., 2010; Nekhotiaeva et al., 2004) (this is markedly different from eukaryotic cells, where the optimal length is typically 15–25 bases), and for optimization we therefore synthesized truncated versions (11, 10, and 9 nucleobases) of the selected peptide–PNA conjugates (Fig. 1). The most active of these conjugates (MIC value of 2 μM and 3 μM, respectively), are an 11-base anti-acpP PNA (PNA3890) with target locations −6 bp upstream to +5 bp downstream, and a 10-base anti-ftsZ PNA (PNA3849) with target locations +4 bp to +13 bp downstream. These were selected for further studies. Mismatch control PNAs (3815 and 3873) were made by interchanging the position of 2 bases inside the antisense PNA sequence, which we consider the optimal design for antisense mismatch controls (Tables 1, 2). Surprisingly, both of these mismatch controls exhibited antibacterial activity only marginally lower than that of the corresponding fully matched PNA (see figure inserts in Tables 1, and 2), although no sequence match to other targets were found by sequence search in the genome. We therefore reasoned that the antibacterial activity of PNAs 3890 and 3849 was only partially due to an antisense mechanism, but nonetheless represent sensitive mRNA targets, and also concluded that the KFF carrier peptide thus appeared suboptimal for use in P. aeruginosa.
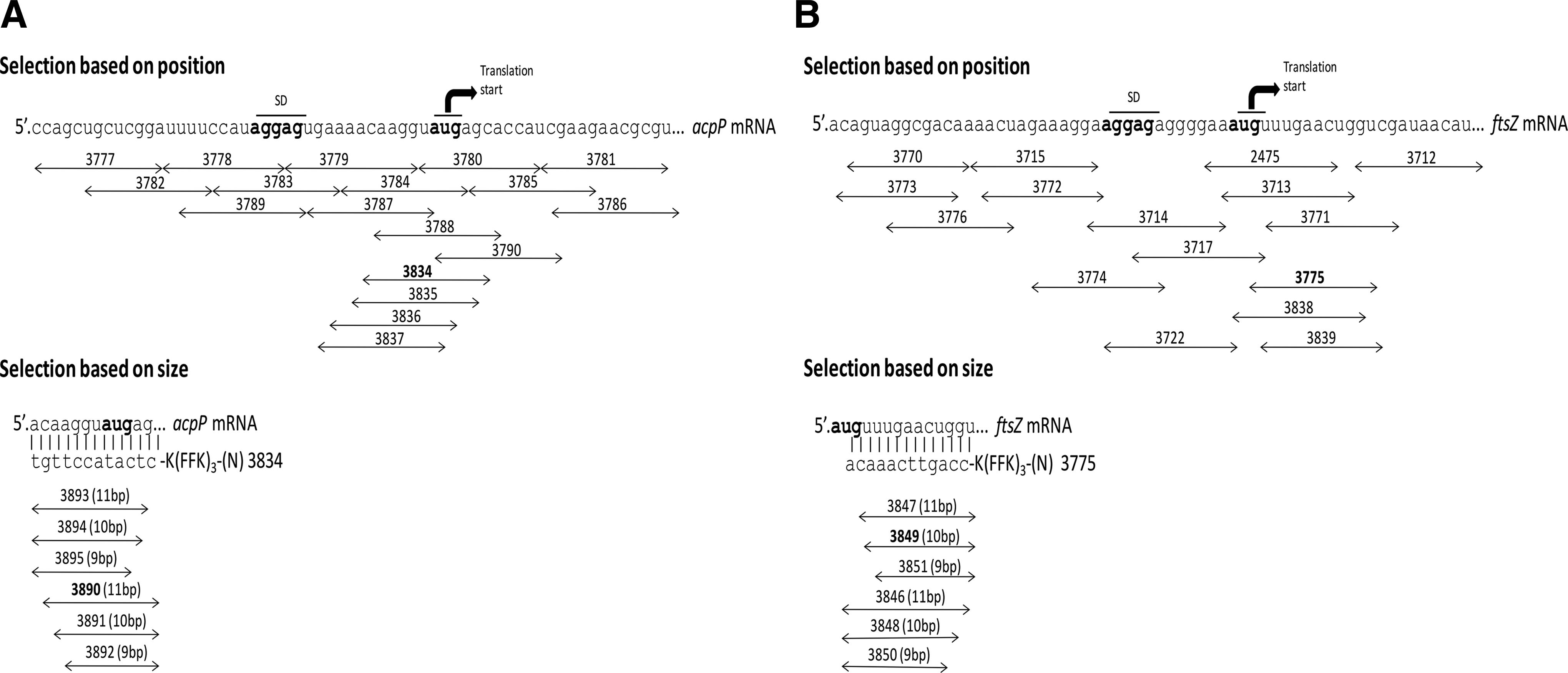
Schematic presentation of the binding region of antisense peptide nucleic acids (PNAs).
Amino acids are shown in uppercase letter and peptide nucleic acid (PNA) nucleobases in lowercase letter. The PNAs are written from their N- to C-termini, and the N-terminus corresponds to the 5′ end of a conventional oligonucleotide.
MIC values are reported as the concentration resulting in an Optical density (OD) value <0.15 after 8 hours incubation at 37°C.
Experimental growth inhibition data of the most active anti-acpP (KFF)3K–PNA conjugate and a corresponding mismatch control against Pseudomonas aeruginosa PA01.
Ahx, 6-aminohexanoic acid; MIC, minimal inhibitory concentration.
Amino acids are shown in uppercase letter and PNA nucleobases in lowercase letter. The PNAs are written from their N- to C-termini, and the N-terminus corresponds to the 5′ end of a conventional oligonucleotide.
MIC values are reported as the concentration resulting in an OD value <0.15 after 8 hours incubation at 37°C.
Experimental growth inhibition data of the most active anti-ftsZ (KFF)3K–PNA conjugate and a corresponding mismatch control against P. aeruginosa PA01.
Consequently, we decided to use the 2 identified acpP and ftsZ sequence targets in combination with a small series of alternative delivery peptides: H-KKHRKHRKHRKH- (Eckert et al., 2006), H-D(KFFKFFKFFK-), H-FWRIRIRR- (Sánchez et al., 2010), H-FWRRFWRR- (Sánchez et al., 2010), H-(RFR)4-Ahx-βala-, H-(R-Ahx)6-βala-, and H-(R-Ahx-R)4-Ahx-βala-. Of these the 3 latter arginine based peptides were originally identified as effective cell penetrating peptides (CPPs) in eukaryotic systems (Abes et al., 2007), but were subsequently found to also be active in E. coli (Mellbye et al., 2009) and various other gram-negative bacteria (Bai et al., 2012).
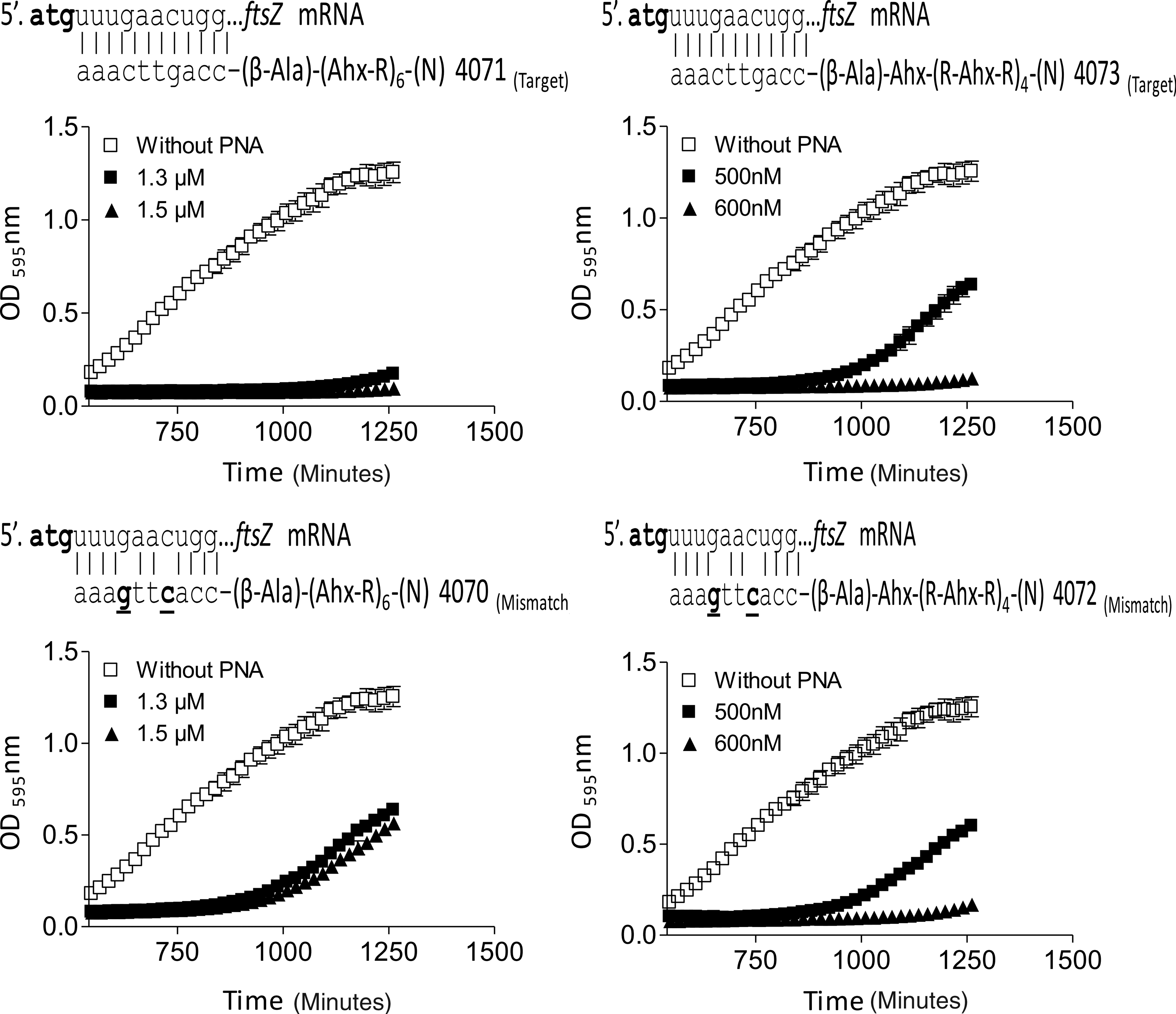
Among this series of peptide–PNA conjugates only PNAs 3969 and 3984 (anti-acpP) and PNAs 4071 and 4073 (anti-ftsZ) based on the H-(R-Ahx-R)4-Ahx-βala and H-(R-Ahx)6-βala peptides showed good activity (MIC≅1–2 μM) in P. aeruginosa PA01. Reassuringly, the corresponding mismatch controls (PNAs 3993, 3985, 4070, and 4072) were much less active both at the 8-hour time point (MIC≅5–20 μM) (Table 3, Fig. 2), as well as at a later time point (20 hours) (Table 4), thereby strongly supporting an antisense mechanism of action. These conjugates further exhibited comparable antibacterial activity against P. aeruginosa PA14 and LESB58 strains, while the corresponding mismatch PNAs were much less active in case of the anti-acpP PNAs (Table 4). However, the mismatch discrimination was less pronounced for the anti-ftsZ PNAs, for which it was practically absent in the LESB58 strain (Table 4 and Fig. 3). We note that a PNA–(KFF)3K conjugate targeting the motA gene was recently reported to inhibit biofilm formation of P. aeruginosa PA01 (IC50≅5 μM) (Hu et al., 2011). However, no mismatch controls were included in this study, and in view of the present results, the effects observed by Hu et al. (2011) may well have been caused by a non-antisense mechanism.

Growth inhibition activity of antisense peptide–PNA conjugates and mismatch controls. Dose dependent effect of anti-acpP and ftsZ peptide–PNA conjugates and their mismatch controls on the growth of PA01. Bacteria were grown in the absence of PNA (closed square) and different doses of peptide–PNA conjugates (open square, open triangle, inverted open triangle, open rhombus, and open circle). PNA 3993, PNA 3985, PNA 4070, and PNA 4072 are the mismatch controls of PNA 3969, PNA 3984, PNA 4071, and PNA 4073, respectively. PNA 3969 and PNA 3984 target the acpP mRNA sequence and PNA 4071 and PNA 4073 target ftsZ, as indicated.
Growth inhibition activity of antisense peptide–PNA conjugates and mismatch controls. Dose dependence effect of anti-acpP and ftsZ peptide–PNA conjugates and their mismatch controls on the growth of LESB58 strain. Bacteria were grown in the absence of PNA (open square) and different doses of peptide–PNA conjugates (closed square and closed triangle).
MIC values are reported as the concentration resulting in an OD value <0.15 after 8 hours incubation at 37°C.
Ahx, 6-aminohexanoic acid; βala: β-alanine.
MIC values are reported as the concentration resulting in an OD value <0.15 after 20 hours incubation at 37°C (PA14 growth inhibition data are shown in Figure S1).
Ahx, 6-aminohexanoic acid; βala, β-alanine.
In parallel, the traditional antibiotics gentamicin, ciprofloxacin, ofloxacin, and ceftazidime were tested under similar conditions for antimicrobial activity against these 3 strains of P. aeruginosa (Table 4). While all strains were sensitive to gentamicin and ciprofloxacin, ceftazidine was less active against PA14, and the virulent LESB58 strain exhibited resistance toward both ofloxacin and ceftazidine. Most interestingly, PNA3969 and PNA4071 showed excellent activity (∼2 μM) against this strain, thereby reemphasizing the potential of peptide–PNA conjugates for the treatment of (multidrug) resistant bacterial infections.
In addition, bactericidal activity of the 4 active PNA conjugates was characterized in PA01. The compounds were tested at twice the MIC (Srinivas et al., 2010). Formally, only PNA 4071 shows bona fide bactericidal behavior, although very significant (up to10-fold) reduction of viable bacterial cells was also observed for the 3 other conjugates (Fig. 4). However, we note a trend indicating that anti-ftsZ antisense agents may be more bactericidal in P. aeruginosa than are anti-acpP agents.
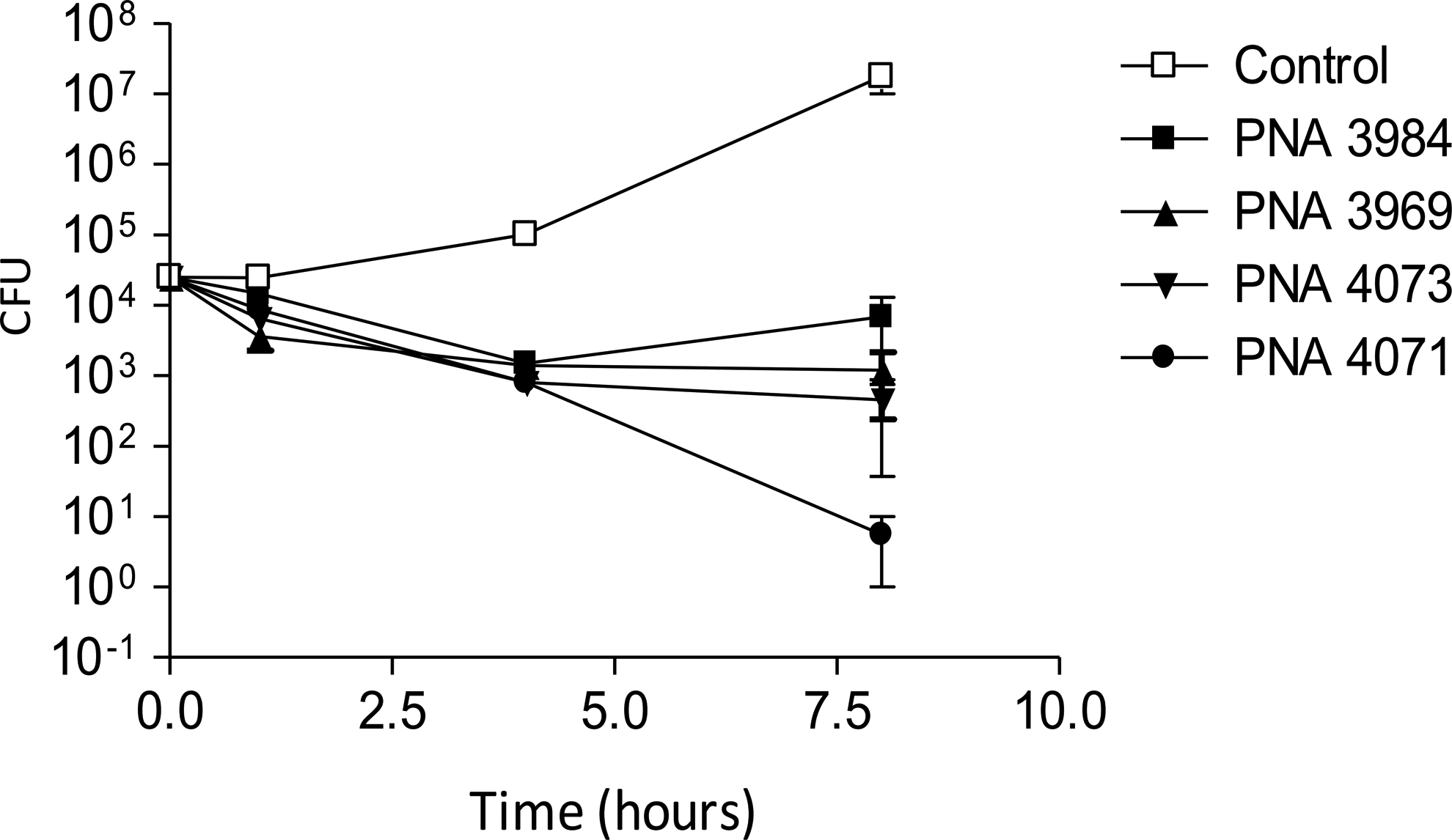
Killing kinetics of anti-acpP and ftsZ peptide-PNA conjugates against Pseudomonas aeruginosa PA01. Bacteria were grown in the absence of peptide–PNA conjugate (open square) or in the presence of 2×MIC of anti-acP PNA, 3984 (closed square) or 3969 (closed triangle), or anti-ftsZ PNA, 4073 (inverted closed triangle) or 4071 (closed circle). Standard error of the mean (SEM) of 2 determinations is indicated. Each point value represents the mean of 2 independent experiments with at least 2 technical replicates for each experiment. MIC, minimal inhibitory concentration.
In order to further characterize the mechanism of action of the antibacterial conjugates, mRNA analysis by quantitative real-time-PCR (QRT-PCR) was performed. Although the molecular antisense mechanism of PNA oligomers in bacteria is believed to rely on translation inhibition via steric blockage of ribosome assembly on the mRNA (Dryselius et al., 2003a), this does lead to specific down-regulation of the targeted mRNA, presumably due to the cellular response of degradation of non-translated mRNA (Goh et al., 2009). Accordingly, we also find a very dramatic reduction (up to 90%) in acpP and ftsZ mRNA after treatment with anti-acpP and -ftsZ PNA, respectively, in the PA01 strain, while only a minor reduction was observed with the corresponding mismatch control PNAs (Fig. 5A, B). (In order to avoid interference of the PNA with the PCR amplification, the primers were chosen so that the amplified RNA does not include the PNA target).The QRT-PCR results were fully corroborated by reverse transcription-PCR (RT-PCR) gel electrophoresis analysis (Fig. 5C), as well as by a dose response experiment using PNA 3969 at shorter incubation time (1 hour) and higher starting bacterial density (requiring higher PNA concentrations). As a final control we detected no change in ftsZ mRNA level in the cells treated with the anti-acpP PNA (Fig. 5 E).
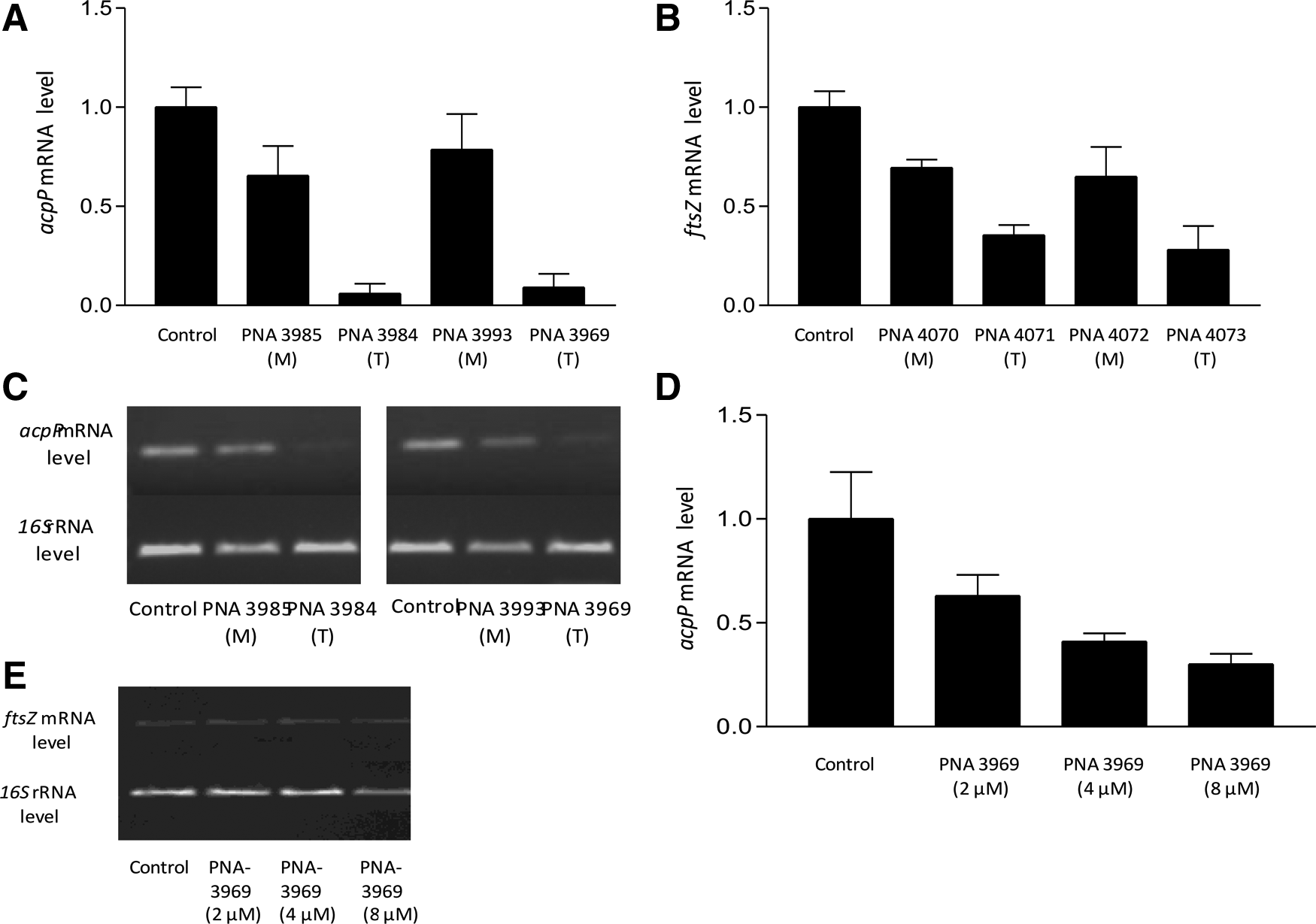
mRNA analysis of antisense treated P. aeruginosa. Relative expression levels of acpP
The combined evidence strongly support a predominant antibacterial antisense mechanism for PNAs 3969, 3984, 4071, and 4073, but an underlying general toxicity of these cationic and slightly amphiphilic peptide–PNA conjugates could be present, analogous to the cell membrane/wall disruptive activity of many traditional antimicrobial peptides such as polymyxin-B. The fluorescent, DNA binding Sytox dye, which is not taken up by live, intact bacteria, is used as an indicator of cell membrane (wall) disruption (Roth et al., 1997; Srinivas et al., 2010). However, a Sytox uptake assay did not give any indication of membrane disruption activity of the peptide–PNA conjugates even at 20 μM concentration, in sharp contrast to the effect of polymyxin-B (Fig. 6).
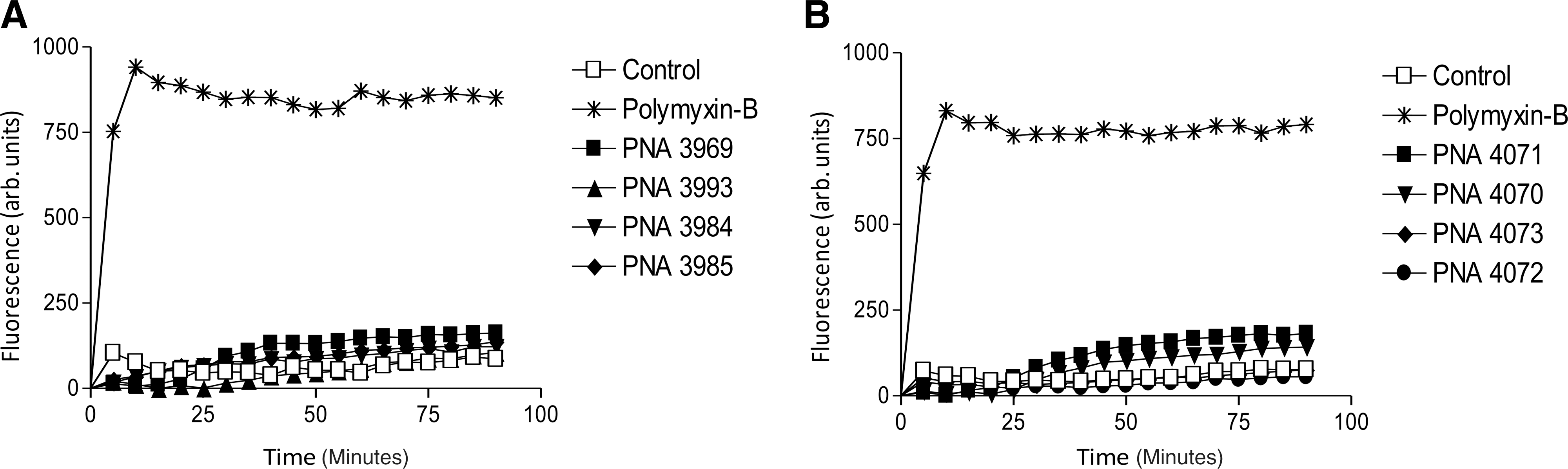
Membrane disruption Sytox assay. Membrane disruption activity in PA01 of anti-sense peptide–PNA conjugates measured by fluorescence of internalized Sytox. Polymyxin-B was used as positive control. All compounds were tested at a concentration of 20 μM.
It is also well established that cationic peptides in general exhibit mammalian cellular toxicity. Thus a preliminary toxicity characterization was performed in a standard in vitro HeLa cell culture assay based on mitochondrial activity. These results showed a surprising difference in toxicity between the 4 “lead” conjugates (Fig. 7). While PNA 3969 was relatively nontoxic (EC50≅14 μM) the 3 other PNAs were significantly more toxic (EC50<5 μM), and in particular PNA 4073 seems very toxic (EC50<< 5 μM). Most interestingly, there is no direct correlation between the peptide or the PNA sequence and the toxicity, but based on the very limited data there seems to be a trend that the ftsZ conjugates are more toxic than the acpP conjugates and that the H-(R-Ahx-R)4-Ahx-βala conjugates are more toxic than the H-(R-Ahx)6-(βala) conjugates. The latter conclusion is supported by the observation that a fully parallel trend in toxicity was found for the corresponding four mismatch peptide–PNA conjugates (Fig. 7). Previous studies have reported limited toxicity (up to 40 μM) of (R-Ahx-R)4–PMO conjugates based on a propidium cell membrane disruption assay (Abes et al. 2006), while some toxicity at 3 μM was observed for a (R-Ahx-R)4–PNA conjugate (similar to the ones used in this paper, but with an 18-mer PNA) using HeLa cells and a mitochondrial activity assay (Saleh et al., 2010), which is essentially analogous to the one used in this study. Both (R-Ahx-R)4 and the (R-Ahx)6 peptide also function as effective CPPs in eukaryotic cells (Abes et al., 2006; Amantana et al., 2007). Therefore it cannot be ruled out that (part of) the toxic effects of the present PNA conjugates (Fig. 7) could be due to “off-target” antisense activity, as theoretically 800–3,000 sequence targets for the 10- to 11-mer PNAs exist in the human genome. However, only a few percent of these would be present in mRNA and only a fraction of these (primarily found in the 5′-UTR region and in intron–exon junctions) would be sensitive to antisense inhibition. Finally, it would be highly unlikely that toxicity would correlate primarily with peptide rather than PNA sequence (compare PNAs 3984, 3985, 4073, and 4072 with PNAs 3969, 3993, 4071, and 4070; Fig. 7). Taken together these results indicate that antisense (R-Ahx-R)4- and (R-Ahx)6- conjugates can (do) show eukaryotic cellular toxicity, but they also emphasize that the toxicity (not surprisingly) is dependent on the conjugated ligand. Therefore one cannot conclude that (R-Ahx-R)4- and (R-Ahx)6- conjugates are nontoxic, and each new conjugate should be investigated as a novel, individual compound.
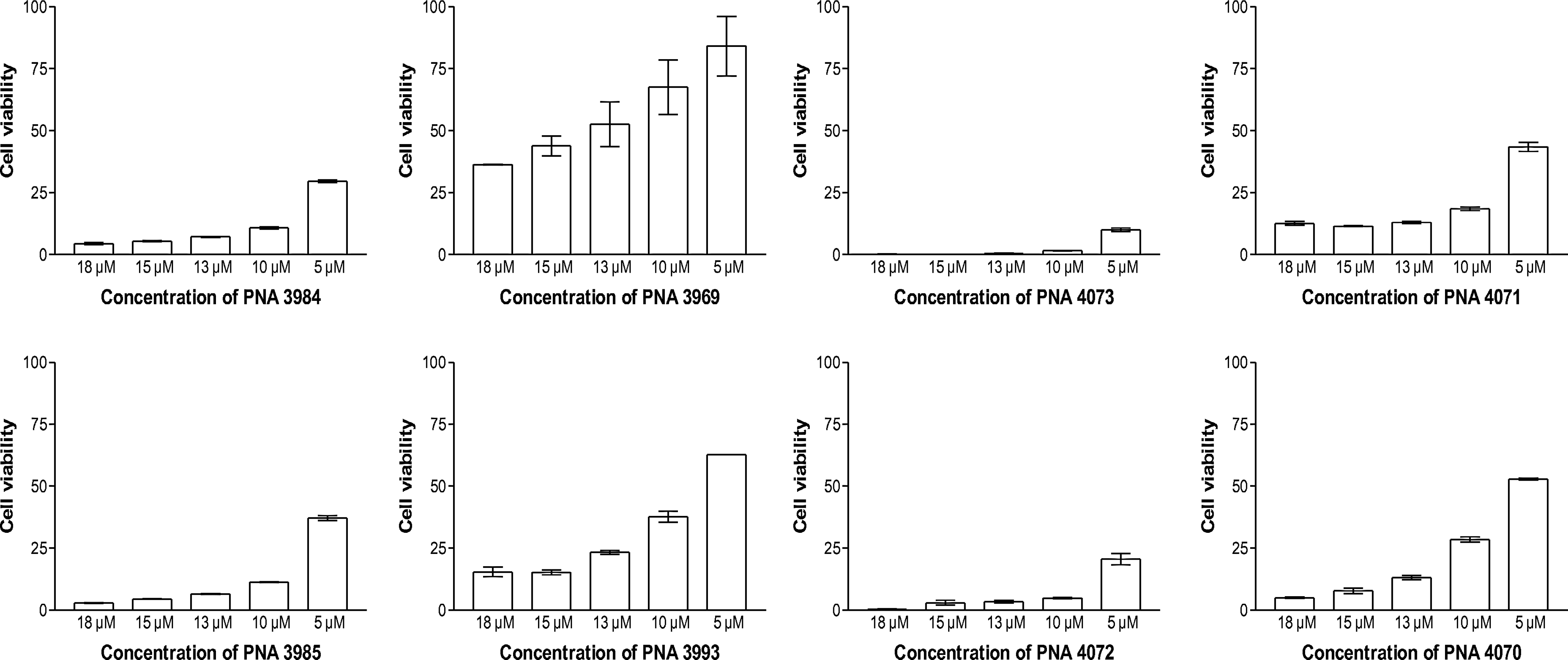
Cytotoxicity of peptide–PNA conjugates. Toxicity assay of anti-acpP PNAs 3984 and 3969 and anti-ftsZ PNAs 4073 and 4071 in HeLa cell culture. Cell viability was quantified by ATP level. The data are presented as the mean of normalized percentage of 2 individual experiments (each comprising triplicate determinations) relative to untreated cells (error bars represent SEM). ATP, adenosine triphosphate.
Conclusions
In the present study we have employed sequence target and carrier peptide optimization in order to identify a series of antisense peptide–PNA conjugates with potent antibacterial activity against P. aeruginosa. The activity of 4 of the most active conjugates was studied in more detail, and the results provide compelling evidence for a bona fide antisense mechanism of action. Our results also emphasize the importance of careful, critical, and stringent data interpretation and the need for carrier optimization. It seems clear that the KFFKFFKFFK carrier peptide, which has successfully been used for a decade in other gram-negative bacteria (Good and Nielsen, 1998; Good et al., 2001; Dryselius et al., 2003a; Dryselius et al., 2003b; Nekhotiaeva et al., 2004; Nikravesh et al., 2007), particularly E. coli, is not efficient for delivery of PNA (and by inference PMO) antisense agents to P. aeruginosa, and that “Ahx-Arg” peptides (Mellbye et al., 2009; Bai et al., 2012) are more promising in this case. Because the mechanism(s) for bacterial uptake of these peptides is not understood, the reason for this difference is not clear. However, recent results have implicated the inner membrane protein SbmA in the uptake of KFFKFFKFFK–PNA conjugates in E. coli (Ghosal et al., in preparation), and sbmA is not found in P. aeruginosa. Further studies are required in order to resolve this question as well as unravel a more detailed understanding of the mechanisms of uptake. Despite the poor uptake of KFFKFFKFFK–PNA conjugates in P. aeruginosa as indicated by the weak activity discrimination between match and mismatch PNAs 3890-3815 and 3849-3873 (Tables 1, 2), it is noteworthy that most of the KFFKFFKFFK–PNA conjugates, including the mismatch PNAs, in fact exhibit low micromolar MIC values. Furthermore, the antibacterial activity of these conjugates differing only in PNA sequence span an order of magnitude (Tables 1, 2). We conclude from this that KFFKFFKFFK–PNA conjugates inherently exhibit general non-antisense mediated antibacterial activity, which is significantly higher than that of the naked peptide (Good et al., 2001 and unpublished data) but is dependent on the sequence and length of the conjugated PNA. The molecular mechanism behind this toxicity could be of both mechanistic and antibacterial drug discovery interest in its own right and is most likely related to ionic interactions with the bacterial cell membrane/wall. However, according to the present results it is not a consequence of direct cell membrane/wall disruption as seen for polymyxin-B (Fig. 5), for example, and thus the mechanism remains obscure at this stage.
Previous studies have demonstrated antibacterial activity of anti-acpP 11-mer morpholino PMO H-(R-Ahx-R)4-Ahx-βala and H-(R-Ahx)6-βala conjugates in E. coli, as well as in vivo activity of these conjugates in a mouse model (Mellbye et al., 2009). Thus by analogy it is conceivable that PNA3969 will also show activity against P. aeruginosa in vivo. Additionally, the CPP activity of the R-Ahx ligands may eventually allow treatment of conditions that involve intracellular infections. Almost certainly, further medicinal chemical optimization in terms of potency (bacterial uptake), toxicology, and pharmacokinetic profile in relation to indications and optimal routes of administration will be required in a drug discovery/development phase, but the present results provide the first step toward antisense antibiotics against P. aeruginosa. We finally note that the specific acpP target is well conserved in other Pseudomonas species for which the sequence data are available (Table 5), thereby potentially allowing for more broad spectrum action of a putative antisense drug.
Anti-acpP and anti-ftsZ PNA sequence alignment in various species of Pseudomonas (from http://string-db.org).
Footnotes
Acknowledgments
This work was supported by the European Commission Seventh framework NABATIVI project grant No. 223670 and by the Danish Council for Independent Research/Medical Sciences. The technical assistance of Ms. Jolanta Ludvigsen is gratefully acknowledged.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.