
Abstract
Human telomeric DNA has the ability to fold into a 4-stranded G-quadruplex structure. Several G-quadruplex ligands are known to stabilize the structure and thereby inhibit telomerase activity. Such ligands have demonstrated efficient telomerase inhibition in dilute conditions, but under molecular crowding conditions mimicking physiological milieu, stabilization of the telomeric G-quadruplex is often lost. We attempted to demonstrate the enhanced G-quadruplex stabilizing ability under molecular conditions by using twisted intercalating nucleic acids (TINA)-modified oligonucleotides. We have shown using circular dichroism and ultraviolet spectroscopic methods that these TINA-modified short oligonucleotides function as G-quadruplex, inducing agents and participate in the formation of stabilized 3:1 G-quadruplex with the human telomeric oligonucleotide. Using enzyme-linked immunosorbent assay–based telomerase repeat amplification assay (TRAP) assay as well as nondenaturing polyacrylamide gel electrophoresis–based TRAP, we demonstrate remarkable enhancement in their anti-telomerase activity even under molecular crowding conditions. This is the first time in which a G-quadruplex stabilizing agent has demonstrated enhanced activity even under molecular crowding conditions.
Introduction
Here we present twisted intercalating nucleic acids (TINA)-modified oligonucleotides (Fig. 1), which are unique nucleic acids composed of bulged insertions of (R)-1-O-[4-(1-pyrenylethynyl)phenylmethyl]glycerol (monomer P in scheme 1) placed between nucleobases in the middle of oligonucleotides. TINA-modified oligonucleotides can stabilize parallel triplexes (Géci et al., 2007; Schneider et al., 2010) and can also participate in the formation of stable G-quadruplexes (Pedersen et al., 2011). However, the exact position of TINA influences the G-quadruplex stability with both increases (Cogoi et al., 2009) and decreases (Paramasivam et al., 2008) having been observed. Moreover, TINA has also exhibited efficient discrimination between double-stranded DNA and single-stranded nucleic acids (Cogoi et al., 2009).
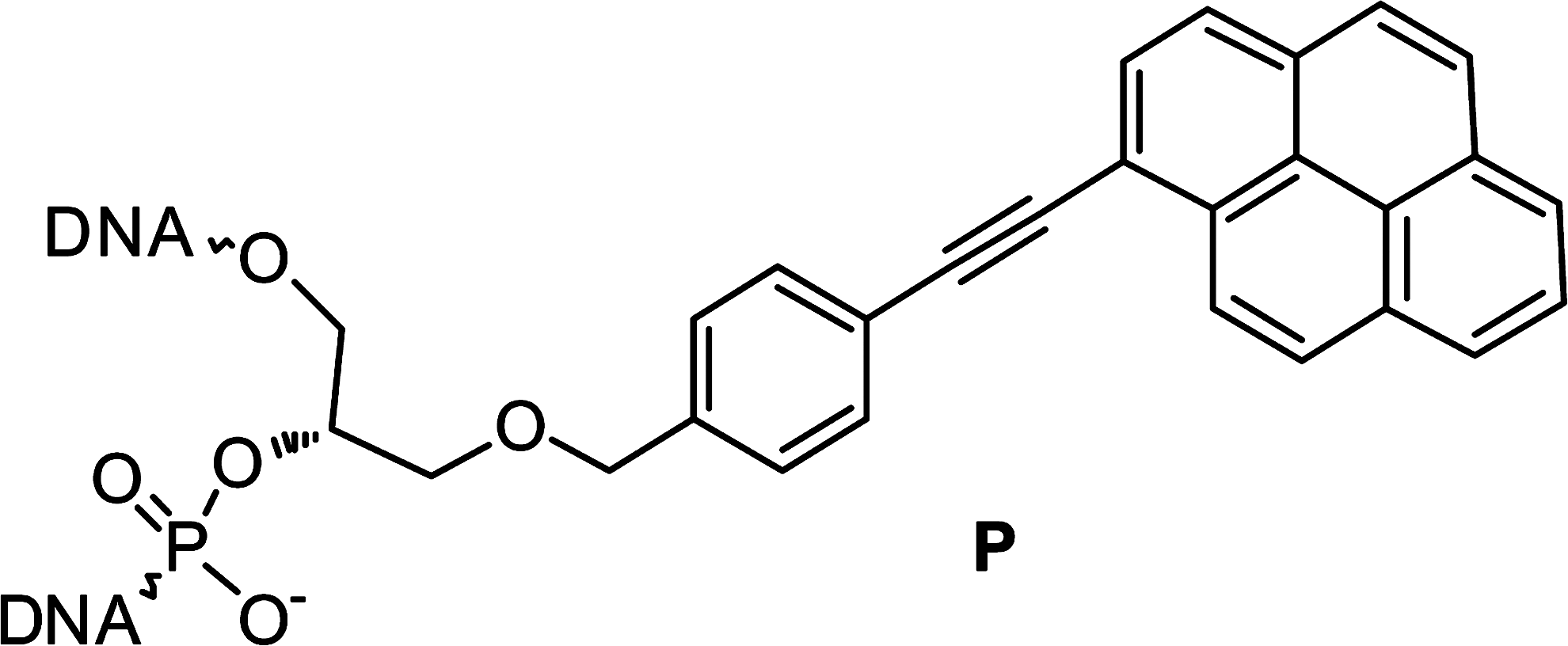
The twisted intercalating nucleic acids (TINA) monomer is denoted by “P.”
Although several studies have provided information on the impact of TINA insertion upon G-quadruplex stabilization/destabilization under in vitro dilute conditions, less is known about the influence of these monomers on G-quadruplex stabilization under molecular crowding conditions. Hence, in the present study we endeavor to investigate the impact of TINA-inserted oligonucleotides on the G-quadruplex stabilization, and hence, anti-telomerase activity in presence of poly ethylene glycol- mol wt. 200 (PEG-200), which acts as a molecular crowding agent mimicking physiological conditions. We finally show that TINA insertions can efficiently participate in forming stable G-quadruplexes with human telomeric sequences in the presence of molecular crowding conditions generated using PEG-200. It was also found that TINA oligonucleotides could efficiently diminish telomerase activity even under molecular crowding conditions. This is the first report in which a G-quadruplex stabilizing agent demonstrates enhanced activity under molecular crowding conditions. TINA insertions were systematically introduced at different positions in the 5- or 6-mer oligonucleotide (Table 1), which could function as a structural component in the formation of an intramolecular G-quadruplex structure when present in a 1:1 complex (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/nat) with the 15-mer human telomeric sequence 5′-GGGTTAGGGTTAGGG-3′. Molecular crowding conditions were achieved with the addition of 30% PEG-200 as described in the literature (Arora et al., 2008; Chen et al., 2009). In this study we show that short TINA-modified oligonucleotides can invade and stabilize a model G-quadruplex system based on the human telomeric sequence under molecular crowding conditions that mimic the intracellular environment. Furthermore, we demonstrate their ability to elicit anti-telomerase activity using an enzyme-linked immunosorbent assay (ELISA)-based telomerase repeat amplification assay (TRAP) assay and nondenaturing polyacrylamide gel electrophoresis (PAGE)-based TRAP and that the anti-telomerase activity increases under conditions of molecular crowding.
P denotes TINA insertion.
n.d. denotes not determined.
Materials and Methods
Oligonucleotides
High performance liquid chromatography–purified 15-mer G-rich human telomeric sequence 5′-GGGTTAGGGTTAGGG-3′ was purchased from Sigma and used without further purification. Synthesis of TINA-modified oligonucleotides was performed as previously reported. Samples were prepared by mixing equimolar amounts of 15-mer human telomeric oligonucleotide with short unmodified or TINA-modified oligonucleotide to generate 1:1 complexes. Thirty percent PEG-200 was used to study changes in presence of molecular crowding.
Circular dichromism spectroscopy
Circular dichromism (CD) spectra were recorded in a Jasco spectropolarimeter 50 (model 715, Japan) equipped with a thermoelectrically controlled cell holder and a cuvette with a path length of 1 cm. Samples were heated at 95°C for 5 minutes followed by slow programmed cooling (0.2°C/minute) in 10 mM Tris buffer, pH 7.4, with 100 mM KCl. CD spectra for quadruplexes (3 μM) were recorded between 220 and 320 nm at 25°C, and the spectra obtained was the average of 3 scans.
Ultraviolet melting studies
One-to-one complexed sample was prepared by heating the oligonucleotides in 10 mM sodium cacodylate buffer pH 7.4, 100 mM KCl, followed by slow cooling. Ultraviolet (UV) melting experiments were performed with 3 μM concentration in a Cary 100 (Varian) spectrophotometer equipped with Hitachi SPR-10 thermo programmer in the absence and presence of different molar ratios of ligands at a heating rate of 0.2°C/minute. Data was collected at 295 nm. Typically 3 replicate experiments were performed and average values are reported.
Nuclear magnetic resonance
Nuclear magnetic resonance (NMR) experiments were recorded on a Varian Inova 500 spectrometer. Samples were prepared by heating to 90°C followed by slow cooling in a 90% H2O/10 % D2O solution, 10 mM sodium phosphate buffer, pH 7, and 100 mM NaCl. The concentration of oligonucleotides was ∼0.15 mM. 1D Watergate NOESY was recorded for all oligonucleotides at 25°C; each spectrum was acquired with 8,192 scans and a repetition delay of 2.8 seconds.
TRAP assay
TRAP assay was performed according to manufacturer's protocol using Telo TAGGG Telomerase PCR ELISA kit from ROCHE. TINA oligonucleotides at 1μM concentrations were used in all cases. Porphyrin was used as the control ligand at 2-μM concentration to achieve visible inhibition of telomerase activity. Twenty percent PEG-200 was used in the mentioned samples to achieve visible results; as with 30% PEG, complete suppression was seen (data not shown). Post polymerase chain reaction amplification, samples were either run on 12% native PAGE and stained with 1×SYBR Gold from Invitrogen, or were processed suitably for ELISA-based detection. ELISA reader from TECAN was used to obtain ELISA readings.
Results
Firstly, CD spectroscopy was performed to investigate the potential of human telomeric DNA to fold into G-quadruplex structures. As illustrated in Fig. 2 and Fig. S2, all of the oligonucleotides in a 1:1 complex with 3 μM of 15-mer human telomeric sequence showed prominent signatures for G-quadruplex, typified by peaks at 295 nm and/or at 260 nm. This showed that the unmodified and all of the 7 TINA-modified oligonucleotides could efficiently participate in the formation of G-quadruplex structures. Moreover, a position dependent effect of the modified oligonucleotides was observed as TINA1, TINA4, and TINA5 showed two peaks at 260 nm and 295 nm indicative of a mixed structure/population, while TINA2, TINA3, TINA6, and TINA7 showed a single prominent peak at 295 nm, suggestive of a predominantly antiparallel G-quadruplex. Additionally, in the presence of 30% PEG-200, the profiles in each case showed stable G-quadruplex structures as illustrated in Fig. 2 (a, c, and e). It was also observed that in most cases the parallel conformation signified by the peak at ∼260 nm became more prominent with a general tendency toward a parallel fold of G-quadruplex.
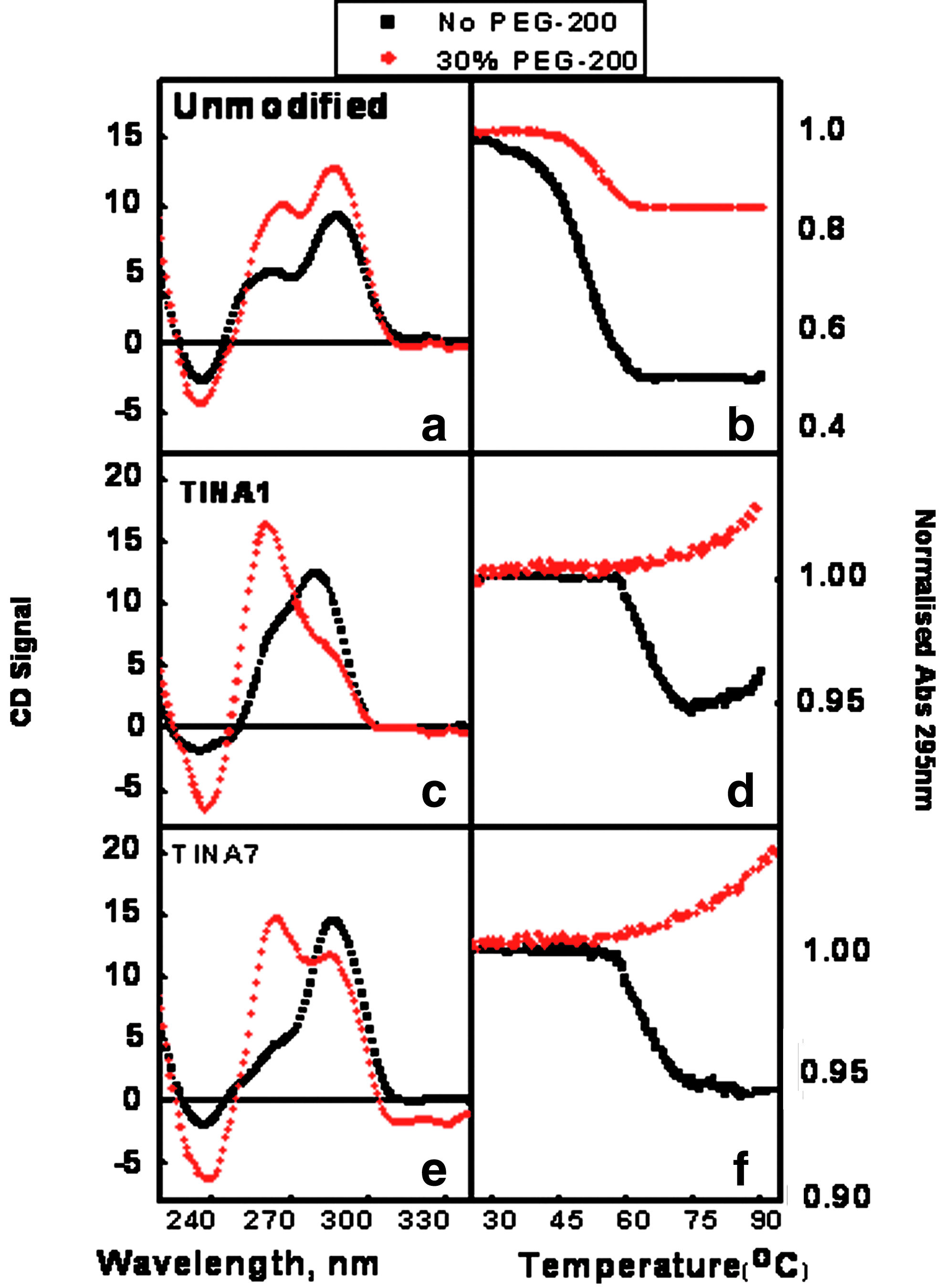
Circular dichromism (CD) and ultraviolet (UV) melting profile of 1:1 complexes. G-quadruplex formation in 1:1 complex of 15-mer human telomeric sequence and different TINA-modified 6-mer oligonucleotide sequences:
UV denaturation/renaturation studies are essential to understanding thermal stabilities of oligonucleotides. Thus, we performed a UV melting study with 1:1 complexes of 3μM of 15-mer human telomeric sequence and unmodified or TINA-modified short oligonucleotides. It was observed that all the sequences showed hypochromic signal with increasing temperature from 20°C to 100°C at 295 nm, characteristic to G-quadruplex forming sequences (Fig. 2b, d, e). The melting temperature of complexes was determined from the normalized absorbance signal (Table 1). Results indicated a position-dependent effect of TINA modification, such that resulting 1:1 G-quadruplex complexes had increased thermal stability. TINA1 and TINA7, where the TINA modification is placed adjacent to a guanine tetrad in a 6-mer, show the highest ΔTms (melting temperatures), 15°C and 16°C respectively. This result is in accordance with previous results reported by Xodo and co-workers, in which TINA insertions led to ΔTms of 22–32°C in G-quadruplexes from KRAS promoter (Cogoi et al., 2009). For all other oligonucleotides, TINA2, TINA3, TINA4, TINA5, and TINA6, either the TINA modification is placed away from guanine tetrads or the parent oligonucleotide is a 4- or 5-mer. These oligonucleotides all gave decreased stability with ΔTms between −7.5°C and −2.0°C This is also in agreement with another report of Xodo and colleagues, in which TINA insertions between guanines resulted in G-quadruplex with depleted thermal stability (Paramasivam et al., 2008). In the context of position dependent effect of TINA, it was shown by Boutorine et al. that the relative positions of TINA insertions could affect the activity of the triple helix formed in the polypurine tract of proviral human immunodeficiency virus (HIV) DNA (Boutorine et al., 2009). They have also indicated the formation and the plausible participation of G-quadruplex in this polyguanine tract in the activity of this triplex of proviral HIV DNA.
Further, in the presence of 30% PEG-200, as illustrated in Fig. 3, all of the above oligonucleotides were further stabilized and melted at temperatures at least 5–6°C higher compared with those in dilute conditions. This shows that TINA insertions could stabilize the 1:1 G-quadruplex complexes in a molecular crowding environment. In contrast, Chen et al. have shown that ligands like TmPyP4, BMVC, Daunomycin, and Hoechst, which could also stabilize G-quadruplex under dilute conditions, showed destabilization of G-quadruplex under molecular crowding conditions (Chen et al., 2009). Further, in our study, TINA1 and TINA7, which have stabilized the 1:1 G-quadruplex complex, exhibited a further stabilization such that the complex could not be melted within the experimental temperature range.
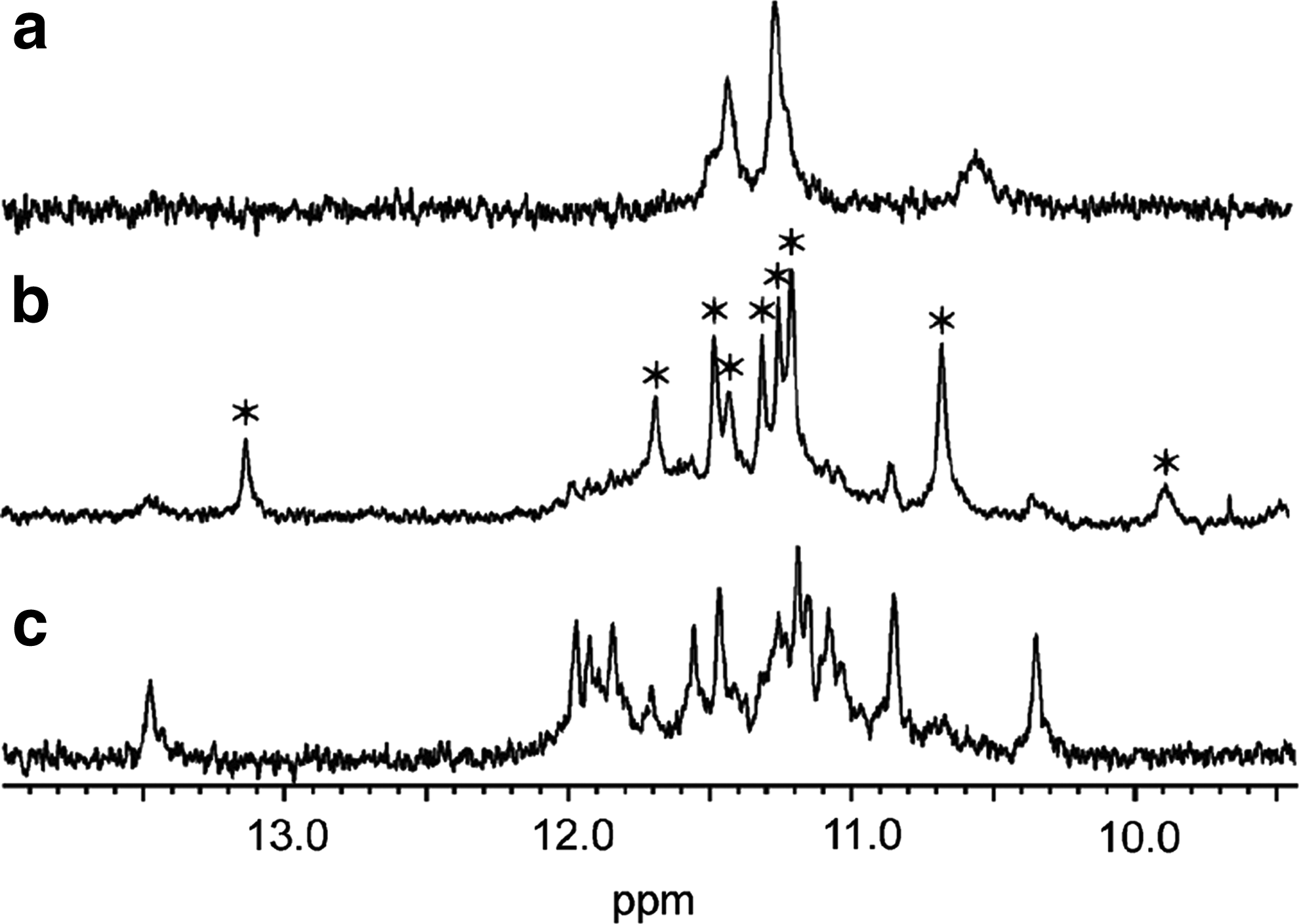
1D 1H nuclear magnetic resonance (NMR) spectra. The imino region of 1D 1H NMR spectra recorded in H2O, pH=7, and 100 mM NaCl at 25°C of
As we observed promising results with TINA1 and TINA7, we used NMR spectroscopy in an attempt to obtain detailed information about the nature of the complexes formed between these TINA-modified oligonucleotides and 5′-GIGTTAGGGTTAGGGT-3′ (I=inosine). The imino region in 1D 1H NMR spectra is informative, as guanine imino protons resonate between ∼10 and 12 ppm when guanosines are involved in G-tetrad formation. Unfortunately, the spectra recorded of 1:1 mixtures of the TINA7-modified oligonucleotides and the G-rich target display fairly broad and overlapping lines, making it impossible to draw conclusions on the formation of a 1:1 complex (Supplementary Figs. S2–S3). However, TINA1spectra recorded with 100 mM Na+ were reasonably well resolved, which provided us with evidence that complexes are indeed formed. In the spectrum of the TINA 1 complex, 8 sharp imino lines show that at least 2 G-tetrads are formed, and the sharp inosine signal at 13.1 ppm is evidence that the complex is formed between the TINA-modified oligonucleotide and the G-rich target with a 1:1 stoichiometry (Fig. 3). The broader line at 9.9 ppm could be either a further guanine imino or a thymine imino protected from exchange with water. Several broad lines that line up with signals from the free G-rich target are also present in the spectrum. These lines might come from a slight excess of the G-rich target. Unfortunately, low concentration of the TINA-modified oligonucleotide precluded analysis of 2D NMR spectra.
Finally, we performed the TRAP to assess the anti-telomerase activity of the TINA-modified quadruplex forming short oligonucleotides. For clarity, we performed both native gel and ELISA-based detection of the amplification products in both the absence and presence of PEG-200. In the native PAGE-based detection of TRAP activity (Fig. 4), it was found that the ladder of telomeric repeat adducts in samples containing PEG-200 generally comprised fewer bands, signifying further decrease in the telomerase activity as compared with the dilute samples without PEG-200. Results were also well in line with previous reports, which showed that Porphyrin stabilizes human telomeric G-quadruplex but loses this efficiency under molecular crowding.
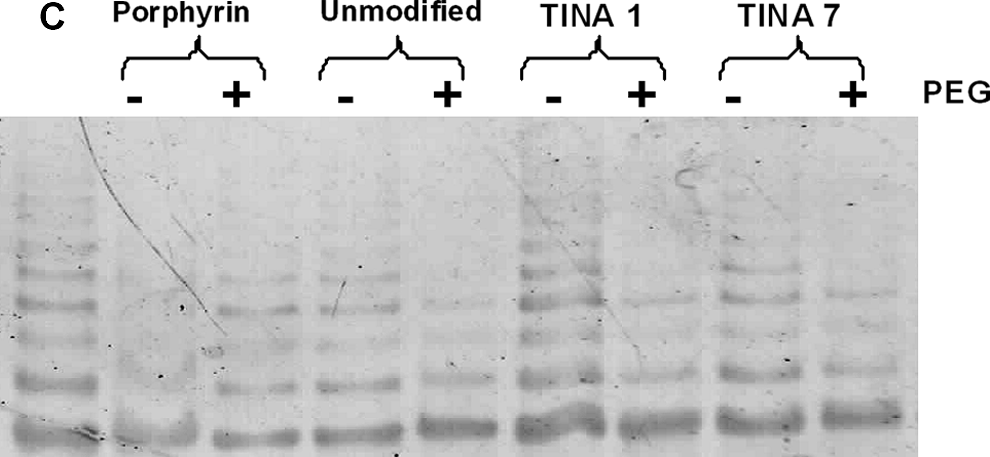
Native gel-based detection of telomerase repeat amplification assay (TRAP). TRAP amplicons show enhanced suppression of telomerase activity in the presence of 20% PEG-200 (see Materials and Methods section) in the case of TINA-modified oligonucleotides. Lanes from left to right show (left) positive control (C) with no ligand; lanes 2 and 3 (Porphyrin) show telomerase activity with porphyrin at 2μM concentration in the absence and presence of PEG-200. The subsequent lanes show the effect of unmodified oligonucleotide, TINA 1, and TINA7 (1 μM concentration each) on telomerase activity in the absence and presence of PEG-200.
Similarly, in the ELISA-based detection of TRAP activity (Fig. 5), porphyrin (TmPyP4), which was used as the positive control, showed inhibition of telomerase activity. TINA1 and TINA7 also showed potent anti-telomerase activity under dilute conditions. Under molecular crowding conditions, the anti-telomerase activity of porphyrin was decreased by as much as 2 times, whereas in case of the TINA oligonucleotides the opposite effect was observed, as both TINA1 and TINA7 showed even further suppression of telomerase activity in the presence of PEG-200.
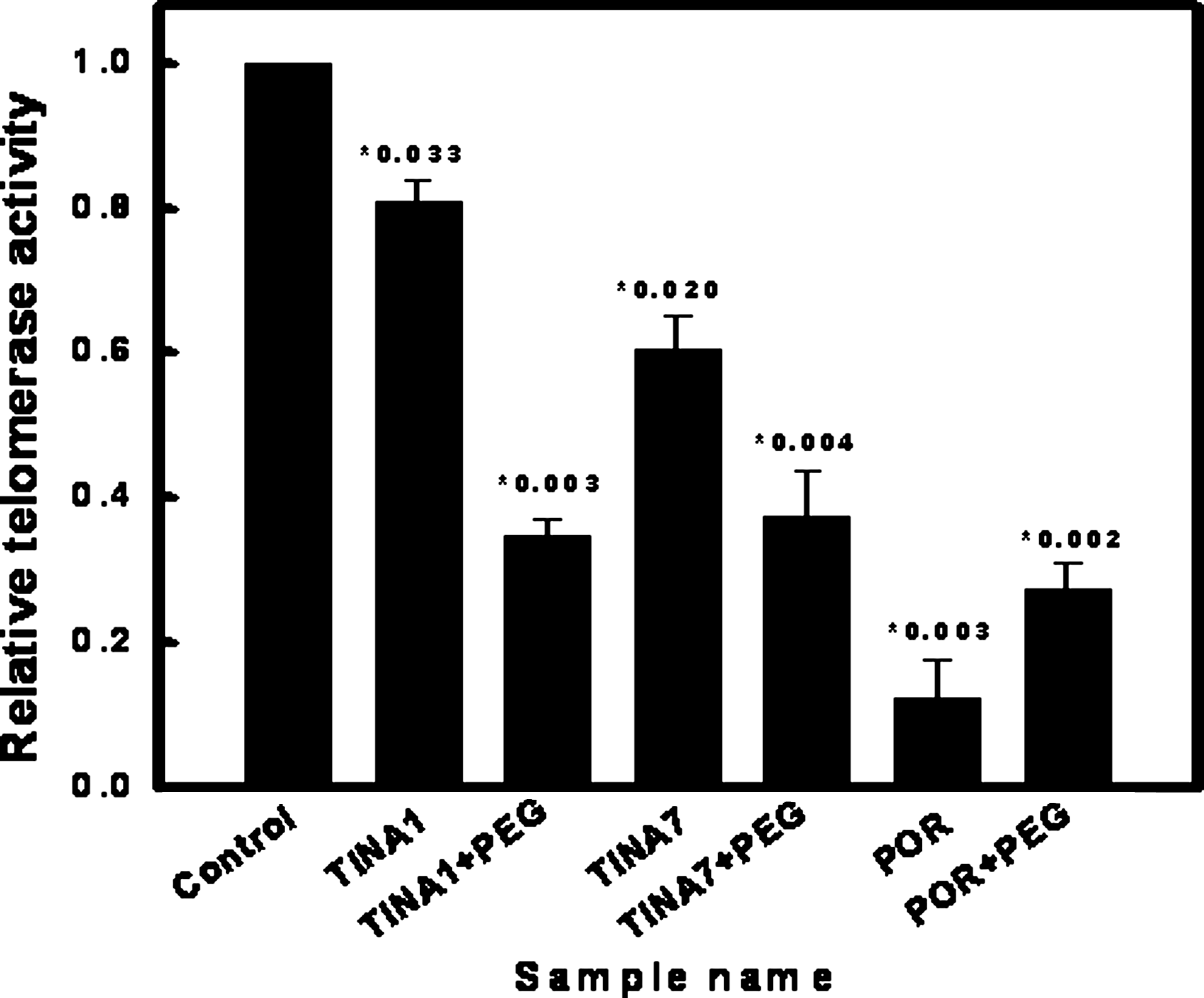
Enzyme-linked immunosorbent assay–based TRAP assay. The relative telomerase activity in the presence of TINA oligonucleotides, in either the absence or presence of 30% PEG-200, is shown. POR denotes porphyrin at 2μM concentration. Each experiment was performed in triplicate and P values for Student's t-test are mentioned along with each bar.
Discussion
This is the first time that G-quadruplex stabilizing agents have been demonstrated to not just tolerate molecular crowding, but rather to perform with increased potency in the presence of PEG-200. Molecular crowding can effectively diminish the activity of external stabilizing agents on account of their altered binding affinity with target G-quadruplex and/or environmental factors contributed by PEG-200, including a changed dielectric constant, changed viscosity, changed water activity, and excluded volume (Chen et al., 2009). In the case of small molecular ligands, like Porphyrin, it has been documented that the binding affinity and stabilization of G-quadruplexes in a molecular crowding environment is notably compromised (Arora and Maiti, 2009). Hence, the efficiency of G-quadruplex ligands under molecular crowding conditions is an important criterion to determine the potency of the ligand as a suitable drug candidate. The present TINA ligands can apparently overcome the impedance offered by molecular crowding and efficiently afford their G-quadruplex stabilizing effect as well as displaying anti-telomerase effect. The stabilization of G-quadruplex by TINA monomers is attributed to the increased stability of the Hoogsteen bonds under molecular crowding effect. In the same context, formation of stable and selective triplexes involving Hoogsteen linkages with TINA monomers has also been demonstrated previously (Filichev and Pedersen, 2005). Additionally, it has been reported by Sugimoto and co-workers that molecular crowding can significantly increase the stability of Hoogsteen bonds in DNA secondary structures due to loss of hydration (Miyoshi et al., 2009). Moreover, hydration significantly affects the thermodynamics of G-quadruplex folding under molecular crowding conditions (Miyoshi et al., 2006), and hence, the same is reflected in the observed thermal stabilization.
Collating all the above results, we can substantiate that the TINA-modified oligonucleotides can be employed for stabilizing G-quadruplexes under both dilute and molecular crowding conditions. Moreover, the telomerase activity that keeps malignant cells in a proliferative and undifferentiated state can be potentially down regulated by these ligands. Optimization of TINA monomers and their position dependent activity will be instrumental to devise appropriate tools to suitably modulate telomerase inhibitory activities of therapeutic oligonucleotides.
Footnotes
Acknowledgments
This work was financially supported by Department of Science and Technology, (Swarnajayanti project to S.M.) Government of India and research fellowship from Council of Scientific and Industrial Research, Government of India, (to T.A.) is gratefully acknowledged.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.