
Abstract
The recent discovery of functional cell-free circulating microRNAs (miRNAs) in human body fluids has opened new avenues for the application of non-coding RNAs (ncRNAs) as noninvasive, specific and sensitive biomarkers for cancers and other human diseases. In this review, we explore the concept of circulating miRNAs as hormones, and discuss their potential functions in cellular communication and transferring of signals. We also provide a brief overview of their identification, processing, and potential functions and applications in human diseases.
Introduction: The Non-Coding Revolution
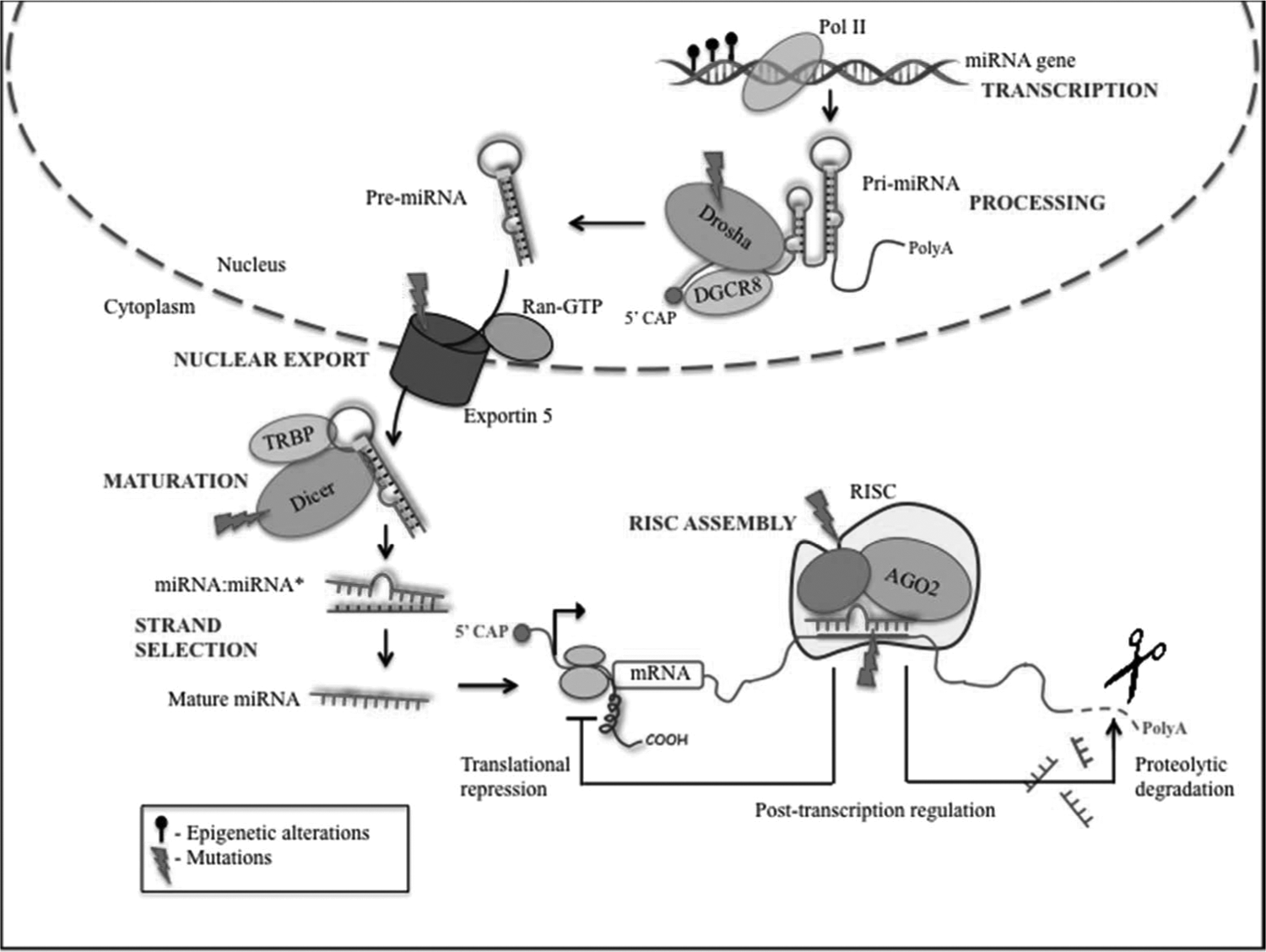
MicroRNA (miRNA) biogenesis and cancer deregulation in the cell. miRNAs are usually transcribed by RNA polymerase II, as autonomous transcription units, or as clusters from a polycistronic transcription unit, to give a stem-looped primary miRNA transcript (pri-miRNA), which is further cleaved by a microprocessor complex (composed of RNase 3 Drosha and double-stranded DNA binding domain protein DGCR8) to give hairpin-shaped precursors of microRNA (pre-miRNAs). The pre-miRNAs are then exported to the cytoplasm by the nuclear export factor Exportin-5/Ran GTP, where they are further processed to ∼22 nucleotide miRNA:miRNA* duplexes by the cytoplasmic RNase 3 enzyme Dicer, and its double-stranded RNA binding domain protein TRBP. Subsequently, the mature miRNA gets incorporated into RISC (RNA-induced silencing complex) that binds to the 3′-UTR of target mRNAs at specific seed sequences or microRNA response elements, and brings about cleavage or translational repression of the target mRNAs. The common alterations in the miRNA biogenesis that occur during cancer are also shown. AGO2, Argonaute2; HDL, high-density lipoprotein.
The traditional view of human physiology limits the functions of cellular communication and signaling molecules mainly to proteins. However, with the discovery of multifaceted miRNAs, the disparities between the functionalities of RNAs and proteins are fading. The most recent asset to the world of ncRNAs has been the discovery of functional cell-free circulating miRNAs in human body fluids, which has introduced a more intricate level of cellular communication and regulation (Mitchell et al., 2008). Circulating miRNAs can act as functional hormones since they are secreted by donor cells into the body fluids as exosomes or free molecules, stably transported to other parts of the organism thus spreading the signals, and are finally taken up actively by recipient cells. The following review focuses on the putative role of circulating miRNAs as hormones and their diagnostic and therapeutic implications in human diseases.
ncRNAs as Stable Circulating Entities: Packaging, Transport, and Uptake
While cellular miRNAs and other ncRNAs were discovered more than 2 decades ago, recent evidence suggests that these RNAs also exist stably in human body fluids, including plasma, serum, saliva, urine, and milk. Circulating RNAs are also detectable in the serum and plasma of cancer patients, being surprisingly stable in spite of the high amounts of RNases circulating in the blood of cancer patients (Mitchell et al., 2008). However, little is known about the mechanisms by which circulating miRNAs are generated and the biological impact of these molecules in distant sites of the body. Currently, there are at least 3 different mechanisms explaining the origin and stability of circulating miRNAs in body fluids (Cortez et al., 2011) (Figure 2): (1) Passive release of miRNAs from broken cells and tissues following tissue injury, chronic inflammation, cell apoptosis or necrosis, or from cells with a short half-life, such as platelets. miR-208 is one such example that is elevated in serum after myocardial infarction (Ji et al., 2009). (2) Active secretion via cell-derived microvesicles, including exosomes and shedding vesicles, which are membrane-enclosed cell fragments released by cells under both normal and pathological conditions (Valadi et al., 2007). Exosomes are formed via inward budding of early endosomal membranes, giving rise to intracellular multivesicular bodies that later fuse with the plasma membrane and release the exosomes to the extracellular environment. Shedding vesicles are larger vesicles that are generated by outward budding and fission of the plasma membrane. Loading of miRNAs into the microvesicles is controlled by specific proteins of the RNA-inducing silencing complex (Gibbings et al., 2009). (3) Active secretion by cells as RNA-binding-protein conjugated complexes. Examples of RNA-binding proteins are nucleophosmin (Wang et al., 2010b), high-density lipoprotein (Vickers et al., 2011), and Argonaute2 (Arroyo et al., 2011).
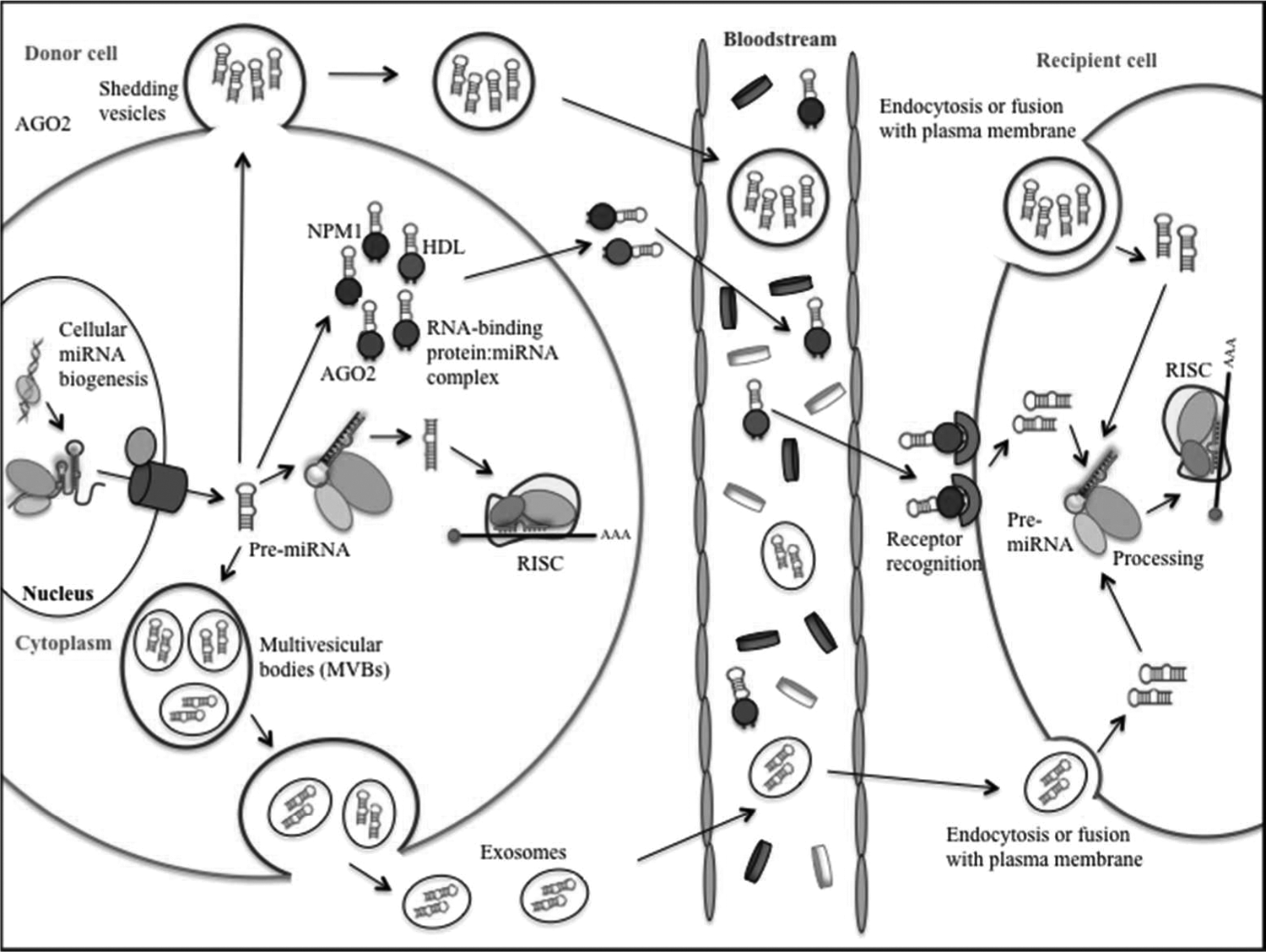
Circular miRNA packaging, transport, and uptake. miRNAs can enter the circulation through three pathways: (1) passive leakage from broken cells; (2) active secretion via microvesicles, including exosomes and shedding vesicles; and (3) active secretion in conjunction with the RNA-binding protein. Once secreted, they are transported across the body fluids in protective capsules and are delivered to the recipient cells. At the site of recipient cells, miRNAs can be taken up via internalization of microvesicles by endocytosis, phagocytosis, or direct fusion with the plasma membranes. RNA-binding protein-associated secreted miRNAs could be taken up via specific receptors on the cell surface.
The diagnostic and prognostic potential of miRNAs as cancer biomarkers relies mainly on their high stability and resistance to storage handling. The molecular basis of the high stability of circulating miRNAs remains largely unknown, although several possible mechanisms have been proposed (Cortez et al., 2011). Firstly, circulating miRNAs might have unique modification, such as methylation, adenylation (Katoh et al., 2009), and uridylation (Jones et al., 2009) that increases their stability and thus protects them against RNases. Secondly, circulating miRNAs are protected by encapsulation in the cell-derived microvesicles. And thirdly, circulating miRNAs that are associated with specific RNA-binding proteins are protected by them. Recently, (Kosaka et al., 2010) showed that the release of miRNAs is controlled by neutral sphingomyelinase 2 and through ceramide-dependent secretory machinery. These results suggest the existence of a specific miRNA export system, wherein certain miRNAs are targeted for export in one cell, and are recognized, taken up, and utilized by another.
Another question of interest is how secreted miRNAs are taken up into recipient cells and whether this process occurs randomly or involves targeting of specific cells (Cortez et al., 2011; Chen et al., 2012). The mode of uptake and its specificity will influence the functional role of secreted miRNAs. It is postulated that microvesicle-enclosed secreted miRNAs are internalized by endocytosis, phagocytosis, or direct fusion with the plasma membranes of the recipient cells. RNA-binding protein-associated secreted miRNAs could be taken up via specific receptors on the cell surface. Cell recognition molecules on microvesicle surfaces or specific receptors on recipient cell membranes could allow specificity in uptake. Though most of the current research on secreted RNAs is focused on miRNAs, these pathways can also be important in transfer of other ncRNAs.
ncRNAs in Intra- and Inter-Cellular Communication
The ability of a cell to communicate with neighboring cells and to modulate its local microenvironment forms the basis for coordinated cellular activity in multicellular organisms. Findings over the past several years suggest the existence of a novel form of intercellular communication: horizontal (cell-to-cell) and vertical (parent-to-offspring) transfer of secreted miRNAs. The role of circulating miRNAs in horizontal transfer of information between cells has been demonstrated in (Skog et al., 2008). They show that glioblastoma-specific proteins and RNAs (such as EGFRvIII and miR-21) are secreted in microvesicles and are converted by recipient normal cells into functional signals to stimulate proliferation and promote tumor progression. Other studies indicate that tumor-secreted miRNAs display the ability to influence the surrounding microenvironment for their own benefit. miR-150 is one such miRNA that is derived from a cultured human monocyte/macrophage cell line and effectively enhances cell migration in human microvascular endothelial cells (Zhang et al., 2010). Moreover, circulating miRNAs might modulate immune responses. For example, (Mittelbrunn et al., 2011) showed that antigen-dependent, unidirectional transfer of miR-335 from T-cells to antigen-presenting cells is important during immune synapse formation. Recent studies have demonstrated that placenta-derived exosomes (containing miR-517a) from trophoblasts are secreted into maternal circulation where they function as immune regulators in fetal–maternal crosstalk, thus improving maternal acclimatization to pregnancy and promoting fetal allograft survival (Luo et al., 2009). This suggests that circulating miRNAs might also contribute to vertical transfer of genetic information. Essentially, secreted miRNAs represent a newly discovered regulatory mechanism by which donor cells can influence the gene expression of recipient cells, and thus impact physiological and pathological processes.
Extracellular ncRNAs as Biomarkers in Human Diseases
Secreted miRNAs have many essential features of useful biomarkers. miRNAs are stable in various bodily fluids, they can be extracted and measured non-invasively, the sequences of most miRNAs are conserved among different species, the expression of some miRNAs is specific to tissues or biological stages, and the level of miRNAs can be easily assessed by various methods, such as quantitative polymerase chain reaction, the gold standard for microRNA quantification in clinically certified laboratories. Alterations of miRNA levels in plasma, serum, urine, and saliva have already been associated with several diseases. The identification of cell-free miRNA in body fluids was first described in patients with diffuse large B cell lymphoma, where high levels of tumor-specific miR-21 correlated with improved relapse-free survival (Lawrie et al., 2008). Hundreds of similar studies have been performed since then, and several other circulating miRNA have been listed as useful potential biomarkers for cancer diagnosis and prognosis (Table 1). Circulating miRNAs are also associated with inflammatory diseases. Circulating miR-146a and miR-223 were significantly reduced in septic patients when compared with healthy controls (Wang et al., 2010a); while miR-150 levels in leukocytes and plasma correlate with the aggressiveness of sepsis (Vasilescu et al., 2009). Reduced plasma levels of miR-132 were observed in patients with rheumatoid arthritis and osteoarthrosis compared with healthy controls (Murata et al., 2010). Furthermore, during acute myocardial infarction, plasma levels of the cardiac-specific miR-208 are elevated (Ji et al., 2009). Recently, an unusual role of let-7b in neurodegeneration in Alzheimer patients via direct interaction with Toll-like receptor 7 (TLR7) was proposed (Lehmann et al., 2012). In a similar study, Fabbri et al. showed that tumor-secreted miR-21 and miR-29a act as ligands to TLR7 (murine) and TLR8 (human) in immune cells and trigger a paracrine prometastatic inflammatory response that ultimately leads to tumor growth and metastasis (Fabbri et al., 2012). In addition to diagnosis, circulating miRNAs are also important as prognostic [miR-21 in diffuse large B-cell lymphoma (Lawrie et al., 2008)] and predictive [miR-125b as a predictor of resistance to adjuvant chemotherapy for breast cancer (Wang et al., 2012)] biomarkers and show correlation to histological subtypes, different stages of cancer, and response to therapy.
lncRNA, long non-coding RNA; miR, microRNA; ncRNA, non-coding RNA.
At present, few other ncRNAs have been characterized as potential biomarkers in human fluids. The long non-coding RNA (lncRNA) PCA3 (prostate-cancer antigen 3) is highly expressed in the urine of prostate cancer patients and serves as a more specific and sensitive biomarker than prostate-specific antigen (Lee et al., 2011). Another study reports the measurement of lncRNA HULC (highly upregulated in liver cancer) from the blood samples of the patients with hepatocellular carcinoma (HCC) (Panzitt et al., 2007) and colon adenocarcinomas (Matouk et al., 2009). Comparative studies of lncRNAs in serum from cancer patients and from normal subjects using unbiased high-throughput technologies such as genome tiling expression microarrays or RNA-seq deep-sequencing will possibly reveal novel circulating lncRNAs as potential biomarkers in many types of cancers.
ncRNAs as Attractive Therapeutic Targets
One important aspect of circulating miRNAs is that these molecules affect cellular systems elsewhere in the body and can produce conditions favorable for either disease progression (destructive effect) or prevention (protective effect). In this regard, therapeutic strategies that involve serum miRNAs emerge as an interesting field to be explored. Using this knowledge, some research groups are now exploring miRNA antagonists (antagomirs) as potential therapeutic molecules. Several studies have reported reduction of the corresponding miRNA levels after intravenous administration of antagomirs against miR-16, miR-122, miR-192, miR-194 (Krutzfeldt et al., 2005), miR-122 (Esau et al., 2006) and miR-221/222 (Mercatelli et al., 2008). Intuitively, miRNAs could be suitable candidates to manipulate the microvesicles' target cells by regulating their RNA stability and translation. miRNA-mimics of tumor-suppressor miRNAs can also be administered intravenously to decrease the expression of their oncogenic targets. However, before this strategy can be assessed in humans, more studies are necessary to identify the role of target miRNAs in each type of disease, to provide biological validation of potential miRNA targets and to measure pharmacokinetic features such as biodistribution and dose response.
Future Perspectives and Challenges
miRNA detection in body fluids is an active field of research in the world of biomarkers. However, the biological context and significance of several of these non-coding transcripts still needs to be evaluated. A simple standard assay for the rapid detection and quantification of circulating miRNAs in various body fluids, along with appropriate normalization controls, need to be established. Additionally, the physiological significance of the information conveyed by circulating miRNAs needs to be evaluated. A well-characterized panel of circulating miRNAs specific to each type of tumor, tumor grade, response to treatment, patient outcome, and recurrence is needed. A combination of known biomarkers, such as cancer-related mutations, or chromosomal aberrations, together with miRNAs levels, can increase the specificity and sensitivity of cancer detection. Large prospective clinical studies will then be required to validate the usefulness of these biomarkers. With the identification of circulating miRNAs being just the beginning, more research on other ncRNAs might help identify the potential gold mine of biomarkers. Thus, further understanding of the molecular nature and functions of these regulatory RNAs would help uncover if circulating ncRNAs can serve as novel minimally invasive biomarkers for management of human cancers and other diseases.
Footnotes
Acknowledgments
Dr. Tom Cech profoundly influenced the career of George Calin—years ago (o tempora!) when a medical student in a former Communist Country, G.A.C. sent Dr. Cech his first ever essay on RNAs; his detailed corrections and the written words to continue doing science, although the rough time, were enough to convince G.A.C. that this is his path! Thank you heartily!
G.A.C. is The Alan M. Gewirtz Leukemia & Lymphoma Society Scholar. He is also supported as a Fellow at The University of Texas M.D. Anderson Research Trust, as a University of Texas System Regents Research Scholar, and by the CLL Global Research Foundation. Work in Dr. Calin's laboratory is supported in part by an National Iinstitutes of Health/National Cancer Institute grant (CA135444), a Department of Defense Breast Cancer Idea Award, Developmental Research Awards in Breast Cancer, Ovarian Cancer, Brain Cancer, Prostate Cancer, Multiple Myeloma, and Leukemia SPOREs, the Laura and John Arnold Foundation, the RGK Foundation and the Estate of C. G. Johnson, Jr. M.Y.S. is a Rosalie B. Hite fellow.
Author Disclosure Statement
No competing financial interests exist.