
Abstract
Plasmacytoid dendritic cells (pDCs) represent a highly specialized immune cell subset and are considered to be the main sentinels against viral infections and play an important role in the development of immune tolerance. pDCs are able to recognize cytosine-phosphate-guanosine (CpG) motifs within microbial DNA, which are unmethylated CG dinucleotides in a certain sequence context and trigger the secretion of interferon (IFN)-α and other proinflammatory cytokines. Here we used the typical class A CpG oligodeoxynucleotide (ODN) 2216, the B-class ODN 2006, and the newly synthesized CpG ODN TM64 to explore the potency and kinetics of IFN-α stimulation of pDC. TM64 CpG ODN has a hexanucleotide sequence TCGTGT that leads to an increased cellular uptake and features a CpG nucleotide within the sequence that leads to a potent specific B-cell stimulation, thus characteristics similar to a class B CpG. Our data reveals that all CpGs act as both dosage- and time-dependent stimuli of IFN-α secretion. The relationship between concentration of the stimulant and the secreted amount of IFN-α is not linear and results in a plateau formation, with saturation kinetics. Alteration to the backbone can change duration and quantity of overall IFN-α secretion.
Introduction
Under the influence of interleukin (IL)-3 and clusters of differentiation (CD40)-ligand they can also adjust the immune response to a tolerance induction. This happens by polarizing the secretion on cytokines, which lead to a TH2-specific immune response, like IL-4 and IL-10 (Rissoan et al., 1999).
Three different classes of CpG ODNs have been identified. They are grouped by their structural assembly, their ability to be stimulated by IFN-α, and by their effect on B-cells (Vollmer et al., 2004). Class A CpG ODNs consist of a chimeric phosphorothioate and phosphodiester backbone, 1 or more CpG dinucleotides in the form of a palindrome, and poly-guanosine (poly-g) motifs at either or both ends. Class A induces large amounts of IFN-α, almost as much as is induced by a virus infection of pDC, but has almost no effect on B-cells (Krug, et al., 2001). Class B CpG ODNs consist of 1 or more CpG dinucleotides on a pure phosphorothioate backbone. They mainly cause an activation of B-cells. The expression of antigen-presenting and costimulatory molecules will be upregulated, the synthesis of immunoglobulins increased, and the secretion of IL-6 and IL-10 stimulated (Krieg et al., 1995). On the contrary, the IFN-α induction is weak. The class C CpG ODNs consist of a 5′ stimulatory hexameric CpG motif linked by a T-spacer to 3′ palindromic sequences that are GC-rich but miss a chimeric phosphorothioate and phosphodiester backbone or poly-g endings. Class C combines the functional characteristics of classes A and B (Hartmann et al., 2003).
Native DNA, which has a phosphodiester backbone, is rapidly degraded by nucleases in vivo. Modified CpG motifs with a nuclease-resistant phosphorothioate backbone have an improved stability and show a more efficient cellular uptake (ZHAO, 1993; Temsamani et al., 1994). The poly-g motif at the end also leads to a better cellular uptake and immunostimulatory activity (Dalpke et al., 2002). It is known that TLR 9 is located intracellularly within the endoplasmic reticulum before stimulation (Latz et al., 2004; Leifer et al., 2004). This indicates that the uptake of CpG ODNs is an important step and regulatory mechanism of immunostimulation via TLR 9.
Although a number of investigations in vivo have been made to use the immunostimulatory qualities of CpG on pDC for therapeutic approaches, little is known of the kinetics of the induced IFN-α secretion. Here we used the typical class A CpG ODN 2216, the B class ODN 2006, and the newly synthesized CpG ODN TM64 to explore the potency and kinetics of IFN-α stimulation of pDC. TM64 CpG ODN has a hexanucleotide sequence TCGTGT that leads to an increased cellular uptake, and it features a CpG nucleotide within the sequence that leads to a potent, specific B-cell stimulation, thus characteristics similar to a class B CpG (Mende et al., 2007).
Material and Methods
Preparation and stimulation of plasmacytoid dendritic cells
Buffy coats were provided by the blood bank of the University of Lübeck and were used to isolate human PBMCs (peripheral blood monocytes). They were obtained from the blood of healthy, non-allergic, and medication-free donors. PBMCs were obtained from buffy coats by Ficoll-Hypaque density gradient centrifugation as described previously (Hartmann et al., 2003). PDCs were isolated by magnetically activated cell sorting using the BDCA-4 dendritic cell isolation kit from Miltenyi Biotec. Cell numbers were calculated by light microscopy and cell viability was determined by trypan blue staining of dead cells as well as using flow cytometry.
Fifty thousand pDCs were cultured and stimulated in 96-well round-bottom plates in 100 μL of medium, comprising Dulbecco's modified Eagle's medium (Gibco), 10% heat-inactivated fetal bovine serum (FCS) (Gibco), nonessential amino acids (Gibco), sodium pyrovate (Sigma-Aldrich), and X-Vivo (Cambrex). Stimulation of the cells was performed for 12, 24, and 48 hours with 3 μg/mL and with 6 μg/mL CpG dissolved in endotoxin-free water. The CpGs used in this work are listed in Table 1.
Lowercase letters, phosphorothioate compound; uppercase letters, phosphodiester compound; accentuated, CpG dinucleotide; CMe, methylcytosine.
Institute of Molecular Medicine, University of Lübeck, Prof. Georg Sczakiel, PhD.
PTO, phosphodiester.
Detection of IFN-α
The IFN-α module set from Bender MedSystems was used to detect IFN-α in cell culture supernatants according to the instructions provided by the manufacturer. The photometric extinction was converted into pg/mL by inverse polynomic regression on the basis of the standard curve. The detection threshold is 3.16 pg/mL in accordance with the manufacturer.
Results
The CpG-induced IFN-α production is dosage dependent
To evaluate the correlation of the added amount of CpG ODN to the amount of secreted IFN-α, isolated pDCs were incubated with CpG ODN 2216 in steps of 0.5 μg up to 6μg/mL with a step to 10μg/mL. The total amount of IFN-α was measured by enzyme-linked immunosorbent assay (ELISA) photometrically after 24 hours of incubation and with help of the standard curve considering the dilution converted into pg/mL.
The smallest volume of 0.5 pg/mL ODN 2216 used in this analysis generated an ELISA detectable amount of 309 pg/mL per 5×104 pDCs. The largest volume, namely 10 μg/mL, generated 5745 pg/mL per 5×104 pDCs (Fig. 1). Nevertheless, the relationship between the concentration of the stimulant and the secreted amount of IFN-α is not linear. Doubling the concentration of the stimulant did not lead to a doubled amount of IFN-α. The highest increase of the secretion took place in the span between 1 μg/mL up to 4.5 μg/mL; subsequently the curve started to flatten and reached a plateau formation between 6 μg/mL and 10 μg/mL.

Correlation of the cytosine-phosphate-guanosine (CpG) oligodeoxynucleotides (ODN) 2216 dosage and the secretion of interferon (IFN)-α. Plasmacytoid dendritic cells (pDCs) were isolated from whole blood. Fifty thousand were incubated with the stated concentrations on the x-axis of CpG ODN 2216 in 96 U bottom well microplates for 24 hours. The quantity of CpG ODN is not proportional to the quantity of secreted IFN-α. It is a non-linear concentration-effect relation. Twice the concentration of the stimulus does not result in twice the amount of secreted IFN-α. The quotient between 3 μg/mL and 6μg/mL is 4.3, and from 5 μg/mL to 10 μg/mL it is 1.6. The slope of the curve is steepest in the area between 1 μg/mL and 4.5μg/mL. After that the increase is smaller and the curve flattens. Effective concentration 100 (EC100) is assumed at the arithmetic mean of the IFN-α concentration at 6 μg/mL to 10 μg/mL (arithmetic mean=5710.6 pg/mL). Therefore, the EC50 equals 2.6 μg/mL CpG ODN 2216. Shown is the arithmetic average of the repeat determination±standard error of the mean (SEM) from n=4.
The mean effective concentration (EC50) describes the effective concentration at which 50% of the maximal effect is observed. On the basis of the results of this study, the EC50 was calculated at 2.6 μg/mL.
The CpG-induced IFN-α production is time dependent
Isolated pDCs in vitro have only a limited lifetime, which can be artificially extended by growth stimulants (Staehelin et al., 1981). In this work we intentionally refrained from these in order not to cause additional changes in the phenotype of the pDCs, which are not due to the effect of CpG ODNs.
To clarify the kinetics of the CpG ODN–induced IFN-α secretion, isolated pDCs were incubated with 3 μg/mL CpG ODN 2216. The IFN-α concentration was measured after 2, 4, 6, 8, 12, 24, and 48 hours. The main part of the IFN-α secretion took place in the first 12 hours of incubation, with the lowest detectable amount of IFN-α after 4 hours of incubation and the highest increase after 8 hours. Considering the total amount secreted after 48 hours as 100%, the kinetic documents that about 50% of the total IFN-α will be secreted within the first 12 hours (Fig. 2).
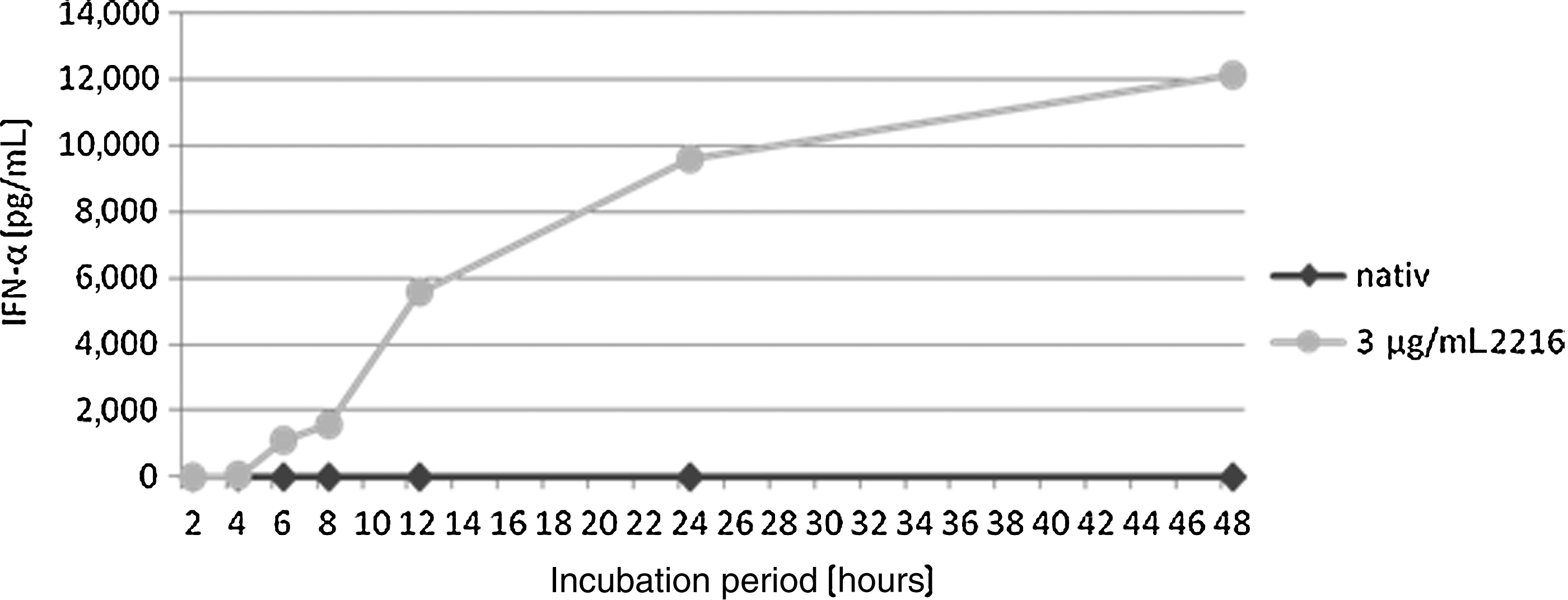
Time response of the CpG ODN 2216 induced IFN-α secretion. Fifty thousand pDCs were incubated with 3 μg/mL CpG 2216 for the periods of 2, 4, 6, 8, 12, 24, and 48 hours. The concentration of IFN-α in the supernatant was detected photometrically. The first measurable IFN-α secretion takes place >4 hours after incubation onset. Between 8 and 12 hours the highest increase occurs, and after 12 hours the progression flattens. Nevertheless, the overall amount still doubles in the following 36 hours. This one experiment run yielded exemplary results.
IFN-α secretion in response to different classes of CpG ODN
The pDCs were obtained from the blood of healthy, non-allergic, and medication-free donors via magnetic cell sorting. They were then incubated with 3 μg/mL CpG ODN 2216, CpG ODN 2006, and TM64. CpG tetraCH2 and plain medium were used as negative controls, with no detectable IFN-α secretion. Among the CpG motifs tested, CpG 2216 showed the highest capability of IFN-α stimulation. The average overall secretion per single pDC was 0.017pg within 24 hours of stimulation with CpG 2216.
The different classes of CpG-induced lower amounts of IFN-α secretion as anticipated. The amount of IFN-α after 24 hours of stimulation with 3 μg/mL CpG 2216 is about 22 times increased compared with stimulation with CpG 2006 (394 pg/mL) (Fig. 3); 2685 pg/mL were detected after stimulation with CpG TM64, which is about one-third compared to stimulation with the class A CpG. The newly synthesized CpG TM64, with the characteristics of a class B CpG but a better cellular uptake (Mende et al., 2007), shows a lower IFN-α induction compared with CpG 2216, but an improved stimulation compared with the typical class B CpG 2006.
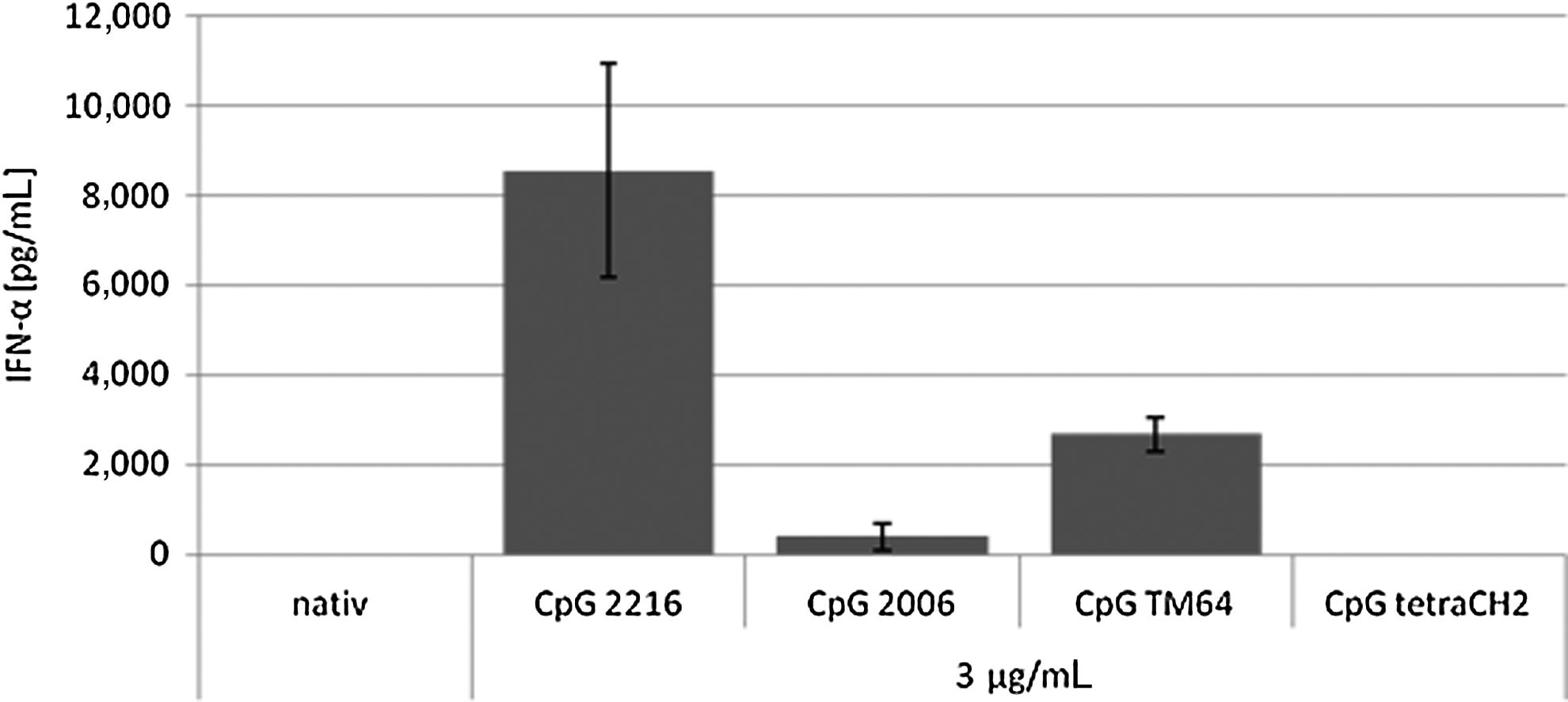
IFN-α secretion in response to different classes of CpG ODN in pg/mL. CpG 2216 reacts as a typical class A CpG with the highest amount of IFN-α secretion on stimulation with different classes of CpG. The class A CpG ODN 2216 induces about 22 times (8553 pg/mL) as much as the class B CpG 2006 (394 pg/mL). Three times less are secreted on stimulation with CpG TM64 (2685 pg/mL). Here we show the results of a 24-hour incubation period with 3 μg/mL ODN per 50,000 pDCs. Native cells function as a negative control and CpG tetraCH2 a nonsense CpG motif as negative control for a immunostimulatory effect. Shown is the arithmetic average of the repeat determination±SEM from n=20.
The concentration (pg/mL) of IFN-α can be directly converted into the activity (U/mL) (Staehelin et al., 1981): 1 U/mL=2.5 pg/mL and 1 pg/mL=0.4 U/mL respectively. Fifty thousand pDCs were incubated in 100 μL total volume per well with 3 μg/mL respectively 6μg/mL ODN for 12, 24, 48, and 72 hours. Native cells only in medium and the negative control ODN tetraCH2 were also tested. There was no medium change under the overall incubation period.
The kinetics of the IFN-α secretion and the time dependence is different in all 3 of the tests of CpG ODN (Fig. 4). The class A CpG ODN 2216 induces the highest amount of IFN-α within the first 12 hours; the total concentration rises in the subsequent 36 hours with a lower secretion rate per time. After 48 hours the IFN-α concentration declines, to 50% after 72 hours. The mRNA has a half-life of 30 minutes; the IFN-α, a biologic half-life of 2–4 hours (Weidle and Weissmann, 1983; Zeuzem and Welsch, 2003).
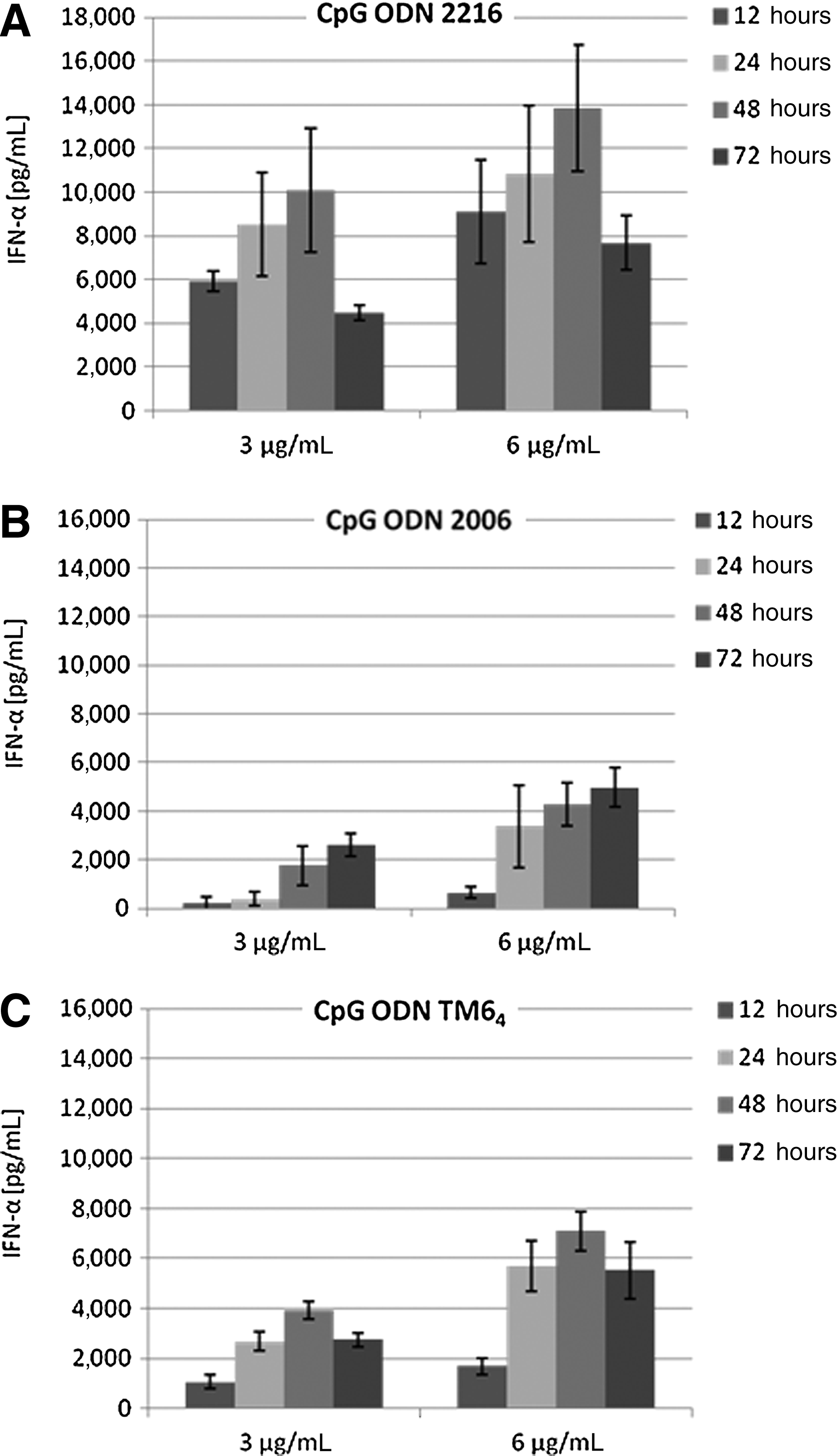
IFN-α concentration (pg/mL) in relation to incubation period (hours] with
The stimulation with the class B CpG 2006 shows different time lapses. The highest secretion takes place after 12 hours (6 μg/mL ODN) decreasing gradually after 24 hours (3 μg/mL). Thus, the concentration does not, unlike ODN 2216, decline after 48 hours. A possible explanation could be the growth-stimulating feature of ODN 2006 that is apparent in an extended lifetime of the pDCs in vitro Krug et al., 2001. CpG TM64 shows like ODN 2006 a later increase of the IFN-α concentration likewise after 12 hours of incubation. The induction capacity is thus higher which may be due to the enhanced cellular uptake Mende et al., 2007. The IFN-α concentration declines after 48 hours likewise after stimulation with ODN 2216 but to a lesser extent (about 20%–30% depending on CpG concentration).
IFN-α secretion in response to CpG ODN sequence modifications
As positive control for the effect of the IFN-α induction, CpG ODN 2216 was used. As negative control, ODN tetraCH2 and native, unstimulated plasmacytoid dendritic cells were used. The ODNs were used both unmodified and with additional phosphodiester (PTO) in their backbone or poly-g ends.
The poly-g elongation with 15 guanosine positions at 3′ of the typical class A ODN 2216 sequence, named ODN 2216-15G, has no effect on the stimulatory activity of the CpG motif and did not show any significant effect on the secreted amount of IFN-α (Fig. 5A).
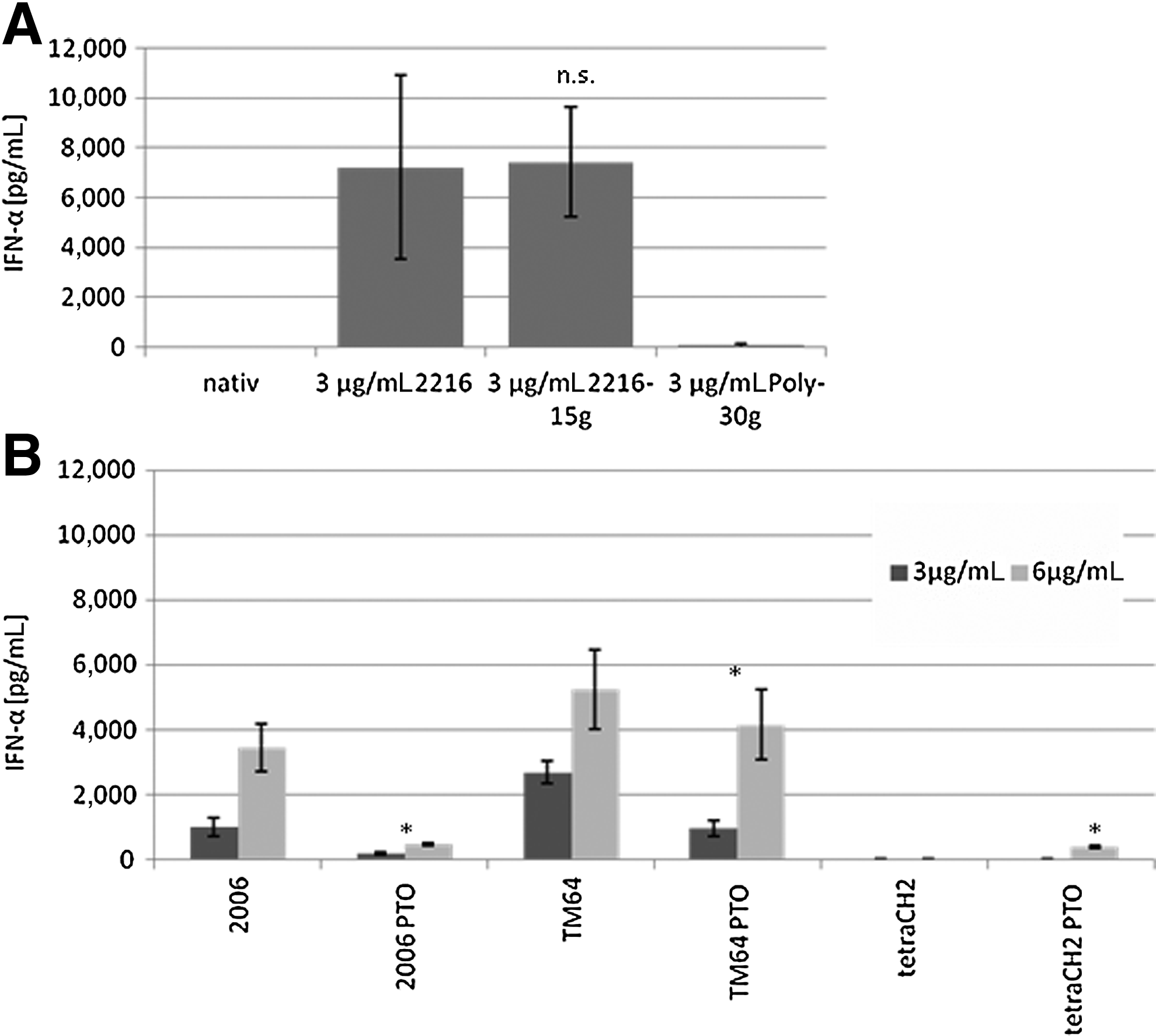
The sequence named poly-g used in this experiment consisted of 30 guanosine positions and did not have any influence on the IFN-α induction.
Comparing non-modified CpG ODN with the PTO modified, the non-modified showed a higher capacity of IFN-α, even though the PTO modifications have their own stimulatory activity. This becomes apparent in the stimulation of the pDC with the nonsense ODN tetraCH2 (negative control) and ODN tetraCH2-PTO (+modified phosphodiester backbone). ODN tetraCH2 does not show any IFN-α activity, whereas the same sequence with PTO modification shows little IFN-α induction (Fig. 5B).
Discussion
CpG motifs as ligands of TLR 9 cause the induction of IFN-α in plasmazytoid dendritic cells. Depending on the sequence/class, they exhibit different kinetics in IFN-α secretion. The class B ODN 2006 and the newly synthesized ODN TM64 show a slower increase and a extended kinetic of the IFN-α secretion as well as a lower total concentration (Fig. 3) compared with the class A CpG ODN 2216.
The class A ODN 2216 reaches its highest increase in secretion within 12 hours, the class B ODN 2006 not until 24 hours. CpG ODN TM64 CpG ODN has a hexanucleotide sequence TCGTGT that leads to an increased cellular uptake, and it features a CpG nucleotide within the sequence that leads to a potent specific B-cell stimulation, thus showing characteristics similar to a class B CpG (Mende et al., 2007). The characteristics of both classes are reflected in the kinetics of the IFN-α secretion, where it combines the features of a class A CpG (an increased IFN-α secretion compared with a classical class B ODN) and class B (an in-time prolonged increase of IFN-α secretion) (Fig. 4). The overall amount of the CpG-induced IFN-α depends on the incubation period and the amount of added ODN. This applies to all CpG ODN classes comparably.
When comparing our results with further data in the literature, the average quantity of secreted IFN-α per single pDC differs. The average overall secretion per single pDC was calculated at 0.017 pg within 24 hours of stimulation with CpG 2216, a maximum of 0.22 pg after 48 hours. Guiducc et al. stated ∼0.4 pg per pDC in their examinations, and Hartmann et al. reported 5 pg per pDC Krug, et al., 2001; Guiducci et al., 2006). A variety of influences can be assumed as an explanation for this wide range of difference.
Even though the plasmacytoid dendritic cells were obtained from the blood of healthy, non-allergic donors without any medication, some influencing factors and bias cannot be eliminated. The ability of secreting IFN-α is increased in times of an acute viral infection and decreases again after convalescence (BERNIER, 1984). Increased IFN-α levels can also be detected in patients with rheumatism or angiitis (Grau et al., 1989). In contrast, the levels are decreased in patients with diabetes or allergies. The expression of TLR 9 is not extenuated but pDCs of individuals with allergies secrete six times less IFN-α on stimulation with CpG than in healthy persons (Tversky et al., 2008). The pDC in the umbilical cord of newborns likewise show a reduced receptiveness on stimulation with CpG ODN (De Wit et al., 2004). By stimulating pDC with CpG ODN they enter a maturation process in which the IFN-α secretion decreases in favor of the IL-12 secretion (Duramad et al., 2003). IL-4, IL-5, and IL-17 alone or in combination with IL-3 in contact with the pDC even before stimulation can decrease the IFN-α secretion (Gary-Gouy et al., 2002). The ligation of blood dendritic cell antigen2 (BDCA2) can also block the secretion of IFN-α (Wu et al., 2008). Oxidative stress is also an influencing factor. It causes the up-regulation of IFN regulatory factor 7 and a reduced IFN-α secretion (Stout-Delgado et al., 2008). All these factors are known to influence the CpG-induced IFN-α secretion, and there are assumably more factors that are yet unknown.
In order to increase the IFN-α secretion and in order to improve the pharmacodynamic characteristics on TLR 9, stimulation modifications of the CpG ODN were examined. Phosphorothioate modifications are a pharmacodynamic adjuvant, which enhance the cellular uptake and make the ODN more stable against enzymatic degradation (ZHAO, 1993; Temsamani et al., 1994). However, changes like this on the native CpG matrix cause in vitro, for example, the activation of transcription factor Sp1, the inhibition of cell proliferation, and cell migration, and also the disruption of the ligand binding of epidermal growth factor receptor and the vascular endothelial growth factor receptor (Perez et al., 1994; Wang et al., 1996; Rockwell et al., 1997). In clinical studies with phosphorothioate-modified CpG motifs, more side effects were observed (Yuen and Sikic, 2000). Besides this, the phosphorothioate modifications feature an intrinsic immunostimulatory function that is autonomous from the CpG motif. Besides the effect on the IFN-α stimulation we were able to show, they also have an effect on B-cell proliferation and the specific immune reactions of PBMCs (Hartmann, et al., 1996; Liang et al., 1996). Still, this kind of modification is of big interest concerning therapeutic applications due to higher stability (KRIEG, 2006).
The second modification we took a look at was poly-g ends. These modifications also lead to a higher cellular uptake in vivo and show less side effects like lymphadenopathia or an extended cytokine boost compared with the PTO modification (Dalpke et al., 2002; Haas et al., 2009). In addition, they show in vitro an increased induction of naïve T-cells via CD8+ T-cell with reduced suppression capability via TLR8 (Dalpke et al., 2002; Moseman et al., 2004).
In our experiments, the addition of 15 guanosine positions at the 3′ end did not have any effect on the stimulatory activity of the class A CpG 2216—neither an increase or decrease of the IFN-α secretion. The sequence with bare poly-g positions did not show any activity at all in terms of IFN-α induction (Fig. 5).
Footnotes
Acknowledgments
We would like to thank Ewelina Szymanski and Brigitte Wollmann for their skillful support in some parts of this work. We are grateful to all members of the Department of Otorhinolaryngology for helpful discussions and a comfortable atmosphere. This work was supported by grants from the Mildred-Scheel-Stiftung (Deutsche Krebshilfe), the Werner and Klara Kreitz-Stiftung, the Monika-Kutzner-Stiftung, and the Rudolf-Bartling-Stiftung.
Author Disclosure Statement
No competing financial interests exist.