
Abstract
Dengue virus (DENV), a mosquito-borne flavivirus, causes serious diseases and threatens public health in tropical and subtropical areas worldwide. RNA interference (RNAi) is a prevailing strategy for antiviral therapy. In this paper, 6 single artificial microRNAs (amiRNAs) targeting the highly conserved regions of the DENV-2 genome were identified and inhibited virus replication efficiently. Then, effective tandem amiRNAs targeting 2 different DENV-2 genome regions were constructed and expressed simultaneously from a single microRNA-like polycistron to avoid virus variation or mutation escape. Finally, the most high-performance tandem amiRNA was embedded in a lenti-viral vector and inhibited DENV-2 virus replication stably and dose-dependently. Overall, these results indicated that RNAi based on multiple amiRNAs targeting viral conserved regions was an effective approach for improvements of nucleic acid inhibitors of DENV and provided a new therapeutic strategy for DENV infection in humans.
Introduction
To deal with this situation, researchers have investigated innovative methods to prevent dengue disease and RNA interference (RNAi) was one of them (Stein and Shi, 2008). David et al. have demonstrated that exogenous small interfering RNA (siRNA) targeting the highly conserved 5′ cyclization sequence (5′CS) region of DENV genome was a powerful inhibitor of all the four representative DENV serotype strains in Huh7 (human hepatoma-derived) cell cultures and in a mouse model (Stein et al., 2011). Synthetic dsRNA can imitate endogenous triggers and has great effect for rapid virus inhibition, but the toxicity and few delivery methods prevent its clinical application. The utilization of Pol2 promoter-mediated artificial microRNA (amiRNA) production could be safe both in vitro and in vivo without the cytotoxic effect of dsRNA and siRNA. Because siRNA without appropriate chemical modification will be sensitive to interferon systems, and short hairpin RNAs (shRNAs) expressed from Pol-3 promoters (especially U6) were possible to saturate the Dicer or other complexes in RNA silencing, which could cause serious effects (Grimm et al., 2006). It was exciting that Pol2 promoter mediated amiRNAs could avoid such side-effects (Giering et al., 2008; Bauer et al., 2009) and were utilized to fight against various viruses (Yekta et al., 2004; Grimm et al., 2006; Ying and Lin, 2006). On the other hand, the low efficient delivery of amiRNA is not always a question to be ignored in RNAi application. This difficulty can be overcome by lentiviruses, which even enabled up to 100% transduction and resulted in long-lasting RNAi effects (Peluffo et al., 2012).
In this study, the newly developed amiRNAs based on the natural structure of Mus musculus miR-155 (Chung et al., 2006; Li et al., 2012) were constructed to target the highly conserved sequences across the most of four serotypes of DENV, and these amiRNAs were then tested for their inhibition activity against DENV-2 infection in cell cultures. Then the 6 effective amiRNAs confirmed in experiments were tandem-linked and expressed simultaneously from the regular or a lentiviral vector for better DENV inhibition.
Materials and Methods
Cells and virus
BHK-21 cells (a baby hamster kidney-derived cell line), Huh7 cells (a human hepatoma-derived cell line), and HEK293T cells (a human embryonic kidney-derived cell line) were obtained from the Cell Bank of Type Culture Collection, Chinese Academy of Sciences. All cells were propagated in Dulbecco's modification of Eagle's medium (DMEM, Macgene) supplemented with 10% heat-inactivated fetal bovine serum (FBS, HyClone), 100 units/mL penicillin, and 100 g/mL streptomycin at 37°C in a humidified atmosphere containing 5% CO2. DENV-2 strain New Guinea C was preserved and maintained in Academy of Military Medical Sciences.
AmiRNAs design and plasmids construction
The overexpression and knockdown (pOK-basic) plasmid (Li et al., 2012), motivating the expression of amiRNA with polymerase 2 promoter and containing an ampicillin resistance gene, was used in this research. The amiRNA expressed from pOK-basic was based on the natural structure of M. musculus miR-155. We collected and aligned full-length genomic sequences of the four serotypes of DENV from National Center for Biotechnology Information references strains, then designed amiRNAs targeting the highly conserved regions across the DENV genome by using Invitrogen online tool Block-iT RNAi Designer (http://rnaidesigner.invitrogen.com/rnaiexpress). These oligonucleotides (Table 1) were annealed and inserted into the pOK-basic vector digested by SapI. The amiRNA targeting none human and mouse genes was also evaluated as a negative control (NC) in this study (McLaughlin et al., 2007). All of these amiRNAs were analyzed with blastn against human and mouse transcript sequences to decrease the probabilities of accidental hybridization incidents. Combination of multiple amiRNAs to a single miRNA polycistron construct was accomplished by digesting a single amiRNA unit with BamHI/XhoI and inserting the digested fragment into the BglII/XhoI sites of DENV-128. For the lentiviral system, plasmids pCMV▵R8.2 (a gift from Dr. Inder M. Verma), pVSV-G (Clontech), and pLNB-MP (containing the BamHI/XhoI sites after the mCherry-T2A-puromycin cassette for amiRNAs subclone) [unpublished data and modified from pLVX-DsRed-Monomer-N1 (Clontech)] were used. Construction of amiRNA expressing lentiviral vector was performed by digesting the multiple amiRNAs cassette with BamHI/XhoI and inserting it into the BamH I/Xho I sites of pLNB-MP.
Mature artificial microRNA (amiRNA) sequences are underscored; the reverse complement sequence of mature amiRNA is italicized.
DENV, dengue virus; NC, negative control.
Packaging lentivector and viral tittering
HEK293T cells were seeded at 7.0–8.0×106 per 10 cm2 culture plate in 10 mL culture medium containing DMEM supplemented with 10% FBS and maintained for 18–24 hours at 37°C with 5% CO2, so that the cell density reached about 80% confluency at the time for transfection. For a plate of cells, 10 μg plasmids (the ratio of pLNB-MP-DENV/pCMV▵R8.2/pVSV-G was 2/2/1) were used and transfected following Sinofection (Sino Biological Inc.) product instructions. The cell culture supernatant was collected 48 and 72 hours after transfection, centrifuged at 3,000 g for 15 minutes and filtered through a 0.45-μm polyvinylidene difluoride filter (Acrodisc) to further remove cellular debris, aliquoted into eppendorf tubes and stored at −70°C until ready for use. HEK293T cells were plated at 1.0×104 per well in a 96-well plate and incubated at 37°C with 5% CO2 for 18–20 hours. One hundred microliter 10-fold serial dilutions of the lentivirus stock ranging from 10–1 to 10–6 were prepared and added to the plate. The medium containing virus was removed from wells 12 hours later and replaced with 200 μL DMEM with 2% FBS. The red fluorescent protein (RFP) positive cells were counted 72 hours later. The well that had more RFP-positive cells (number ≥5) but the RFP-positive cells were zero next it or the well that had fewer RFP-positive cells (number ≤5) was chosen to determine viral titer (recorded as 1 IU). Formula for virus titer calculation: titer=N×1 IU×(DF–1)/V (N, number of RFP-positive cells; DF, virus dilution factor; V, volume of the inoculum) (Jiang et al., 2003).
AmiRNAs transfection and DENV infection
AmiRNA transfection in a 96-well plate was employed for antiviral experiments. BHK-21 cells were trypsinized and plated on 96-well plates at a density of 1.0×104 cells per well for 24 hours before transfection. Each sample was repeated in 4 wells. For the same 4 wells, 0.5 μg amiRNA was used and transfected following GenJet™ Ver-II (SignaGen Laboratories) manufacturer's instructions. The transfection-reagent-containing medium was removed 6 hours later, and the cells were infected with DENV virus [multiplicity of infection (MOI) of 0.2, as indicated] in 100 μL DMEM (containing 2% FBS and antibiotics) for 2 hours with gentle rocking at 24 hours post-transfection. After that the virus-containing medium was removed, the cells were washed twice and replenished with 200 μL DMEM (containing 2% FBS and antibiotics).
Lentiviral transduction of BHK-21 cells
BHK-21 cells were plated in a 96-well plate at a density of 1.0×104 cells per well. After 24 hours, the appropriate volume of virus corresponding to the optimal MOIs (0.25, 0.5, 1.0) was added to the BHK-21 cells. The medium containing virus was removed from wells 12 hours later and replaced with 200 μL DMEM with 10% FBS. The cells were infected with DENV at 72 hours after transduction, as described above.
Cell counting kit-8
Cell culture, plasmid transfection, and DENV infection conditions were as described above. Cell viability was measured at 5 days after DENV infection by using the CCK-8 cell counting kit (Dojindo Laboratories) according to the manufacturer's instruction. The absorbance values of amiRNA-treated cells and virus-infected cells were compared with the uninfected samples containing no transfection reagent and amiRNA.
Cell-based indirect immunofluorescence assay
Cell culture conditions were as those described above, except that Huh7 cells were used. Culture medium was removed by gentle suction 48 hours after DENV infection and the cells were washed 2 times with phosphate-buffered saline (PBS) under gentle shaking for 5 minutes. Then the cells were fixed with 4% paraformaldehyde at room temperature (RT) for 20 minutes. Cells were washed 3 times with PBS under gentle shaking for 5 minutes and then permeabilized in 0.25% Triton X-100 for 10 minutes at RT. Cells were washed 3 times with PBS under gentle shaking for 5 minutes and then blocked with PBS containing 2% normal goat serum plus 0.4% Triton X-100 at RT for 30 minutes. Cells were then washed 3 times with PBS, as above, and then incubated in primary antibody (mouse monoclonal to DENV E glycoprotein, Abcam) at a dilution of 1:10 in a humidified chamber at 4°C overnight. Cells were washed 3 times with PBS, as above, and then incubated with secondary antibody (goat anti-mouse immunoglobin G conjugated with DyLight™ 594 or DyLight™ 488) diluted 1:200 with PBS for 1 hour at RT in the dark. Cells were then washed 3 times with PBS and mounted with slow-fade gold antifade reagent with 4′,6-diamidino-2-phenylindole (Invitrogen).
Virus plaque assay
BHK-21 cells were plated in a 6-well plate, and 2.0×106 cells in 2 mL growth media were added to each well. The cells should be 100% confluent after 12-–4 hours. One milliliter of each supernatant dilution containing virus was collected after 48 hours infection and was added to each well. The infected monolayers were incubated at 37°C with 5% CO2 for 1 hour and mildly shaken several times during this adsorption period. Then 3 mL of the 2% low-melting agarose/growth media mixture were added to each well and the plate was allowed to sit for 15 minutes at RT until the agarose overlay turned solid. After growing for 5 days at 37°C with 5% CO2, cells were fixed with 3 mL 4% formaldehyde at 37°C for 1 hour. Then the formaldehyde and agarose cover were discarded and cells were washed 3 times with PBS under gentle shaking for 5 minutes. Cells were stained for 1 hour at 37°C by adding 1 mL 1.5% crystal violet solution. Cells were washed 3 times with PBS, as above, and then plaques were counted to calculate the concentration of the initial viral in plaque-forming units (PFU) per mL. [PFU=N×DF/V (N, number of plaques; DF, virus dilution factor; V, volume of the inoculum).
Semiquantitative reverse transcription-polymerase chain reaction
BHK-21 cells were plated in a 6-well plate at a density of 2×105 cells per well. After 24 hours, the appropriate volume of amiRNA-expressing lentivirus corresponding to the optimal MOIs (0.25, 0.5, 1) was added to the plated BHK-21 cells. The medium containing virus was removed from wells 12 hours later and replaced with 2 mL DMEM with 10% FBS. The cells were infected with DENV at 72 hours after transduction, as previously described; 48 after hours infection, cell total RNA was extracted by TRIzol Reagent (Invitrogen) following the manufacturer's instruction. Then reverse transcription was performed according to the protocol of FastQuant RT Kit (TIANGEN), and the polymerase chain reaction (PCR) was performed under the following conditions: 94°C for 5 minutes, followed by 30 cycles of 94°C for 30 seconds, 55°C for 30 second, and 72°C for 1 minute, with the final cycle of 72°C for 5 minutes. PCR products were analyzed using 1.5% agarose gel electrophoresis and visualized by ethidium bromide staining, and the bands intensities were measured by ImageJ software. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA expression levels were used for normalization. The primers used for DENV-2 and GAPDH were: DENV-2-F, 5′-AATATGCTGAAACGCGAGAGA-3′; DENV-2-R, 5′-GGG ATTGTTAGGAAACGAAGG-3′; GAPDH-F, 5′-GAGTCAAC GGATTTGGTCGT-3′; and GAPDH-R, 5′-TTGATTTTGGAG GGATCTCG-3′, respectively.
Interferon induction assay
Huh7 cells were plated in a 6-well plate at a density of 2×105 cells per well. Twenty-four hours after plating, the appropriate volume of amiRNA-expressing lentivirus corresponding to the optimal MOIs (0.25, 0.5, 1.0) was added to the Huh7 cells. Nontransducted cells added with 2,000 IU/mL of interferon (IFN)α-2a (3SBio Inc.) for 72 hours served as the positive control. The medium containing virus was removed from wells 12 hours later and replaced with 2 mL DMEM with 10% FBS. Sixty hours later, cell total RNA was extracted using TRIzol Reagent (Invitrogen) following the manufacturer's instruction. Then reverse transcription was performed according to the protocol of FastQuant RT Kit (TIANGEN). The PCR was performed under the following conditions: 94°C for 4 minutes followed by 27 cycles of 94°C for 30 second, 56°C for 30 seconds, and 72°C for 35 seconds, with a final cycle of 72°C for 10 minutes. PCR products were analyzed using 1.2 % agarose gel electrophoresis and visualized by ethidium bromide staining. The primers for the IFN-inducible genes 2′,5′-oligoadenylate synthetase 1 (OAS1), MxA, interferon-stimulated gene 15, and signal transducer and activator of transcription 1 are shown in Table 2 (Sadler and Williams, 2008; Wang et al., 2013).
Statistical analysis
Results were graphed, with error bars indicating the standard deviation. Statistical analyses were done with Prism 5 (GraphPad Software), and statistical significance was determined using Student's t-test or one-way analysis of variance (P<0.001, P<0.01, and P<0.05).
Results
Design and identification of recombinant amiRNA plasmids
All amiRNAs used in this study were defined in Table 1, and the 6 effective amiRNAs targeting regions against DENV genomic sequences are shown in Fig. 1A. Some of these target sequences were highly conserved across the GenBank reference sequences for the 4 serotypes of DENV, especially for serotype 2. All 21 amiRNAs were synthesized and cloned into pOK-basic vector. Six effective amiRNAs were obtained using primary screening tests (data not shown), and 6 miRNA-like polycistron constructs expressing different combinations of amiRNAs were also constructed using the strategy illustrated in Fig.1B. Specifically, the other 5 single amiRNAs were constructed respectively into a chain with DENV-128, whose target sequence was the most conserved across the 4 serotypes of DENV. The amiRNA combination of DENV128-1382, which was the most efficient at inhibiting DENV infection, was selected and inserted into the lentiviral vector pLNB-MP. These recombinant positive clones were selected by ampicillin resistance and confirmed by DNA sequencing.
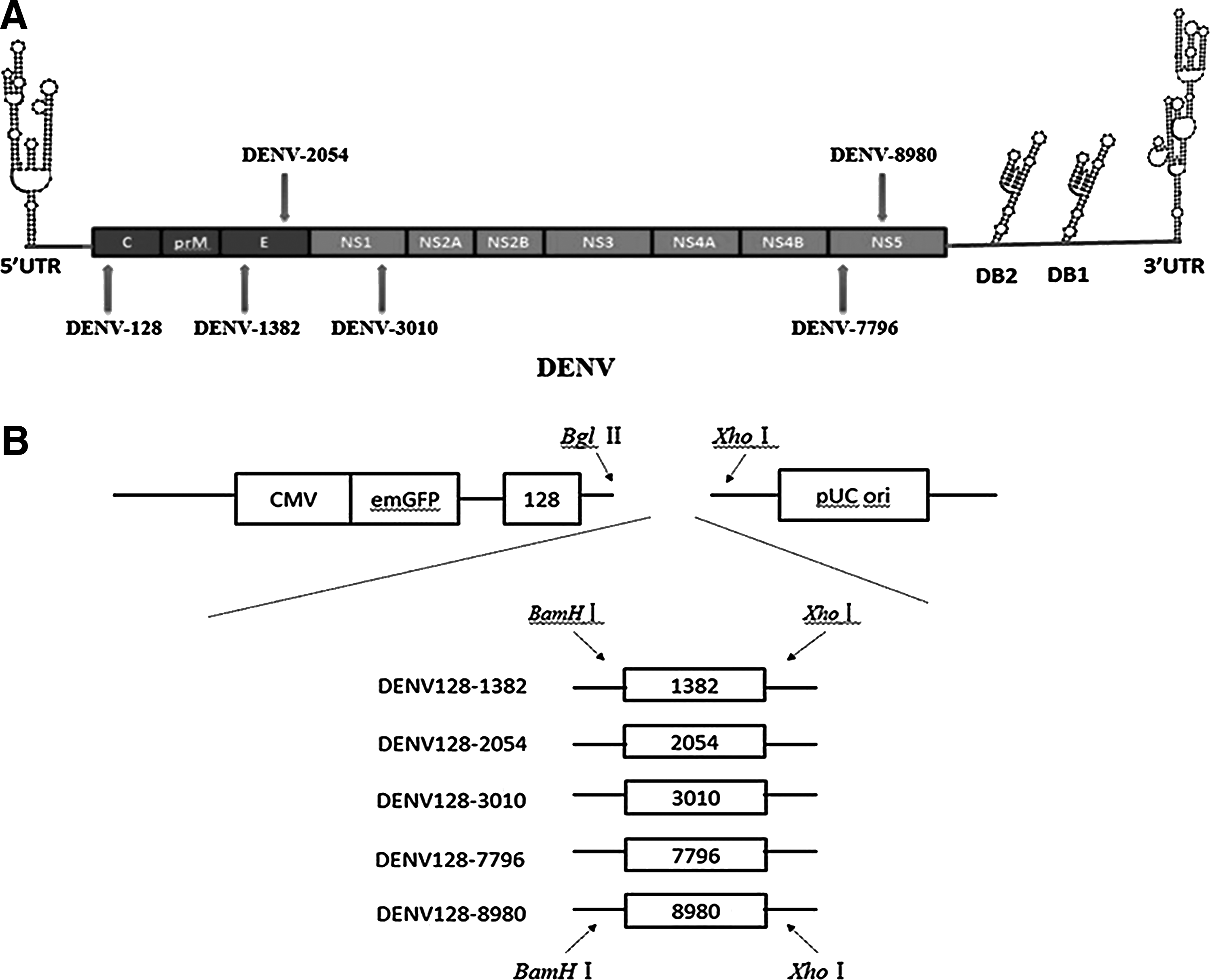
Schematic representations of the dengue virus (DENV) genome structure and the tandem artificial micro RNA (amiRNA) construction strategy.
Antiviral activity of transiently transfected single-amiRNAs
We chose BHK-21 cells as host cell for evaluating the antiviral efficacy of amiRNAs due to their high infection rate by DENV and relatively high amiRNA transfection rate. The negative-control amiRNA (NC), the pOK-basic vector control (BC), and the virus infection control (containing DENV without amiRNA and transfection reagent) were included in each experiment. Cells were transfected with 0.125 μg amiRNA per well 1 day before DENV infection at MOI 0.2. We obtained 6 amiRNAs more effective than the other 15 amiRNAs by comparing their cytopathic effects. We took pictures of amiRNA-treated cells by microscope to detect the cell morphology of BHK-21 cells challenged with DENV (Fig. 2A). Furthermore, the CCK-8 assay was used to assess the protection effect of each amiRNA on viability of infected BHK-21 cells at 5 days after DENV infection. These results indicated that there was a significant protection effect on cell survival rate for amiRNA-treated group (Fig. 2B). All 6 amiRNAs except for DENV-8980 markedly increased the cell survival rate, especially, the cell survival rate of DENV-128 (0.90±0.05), which was 1.81 times the virus infection control rate. This result was generally consistent with the indirect immunofluorescence assay for DENV E (envelope) protein expressed in BHK-21 cells 48 hours post-infection (Fig. 2C).
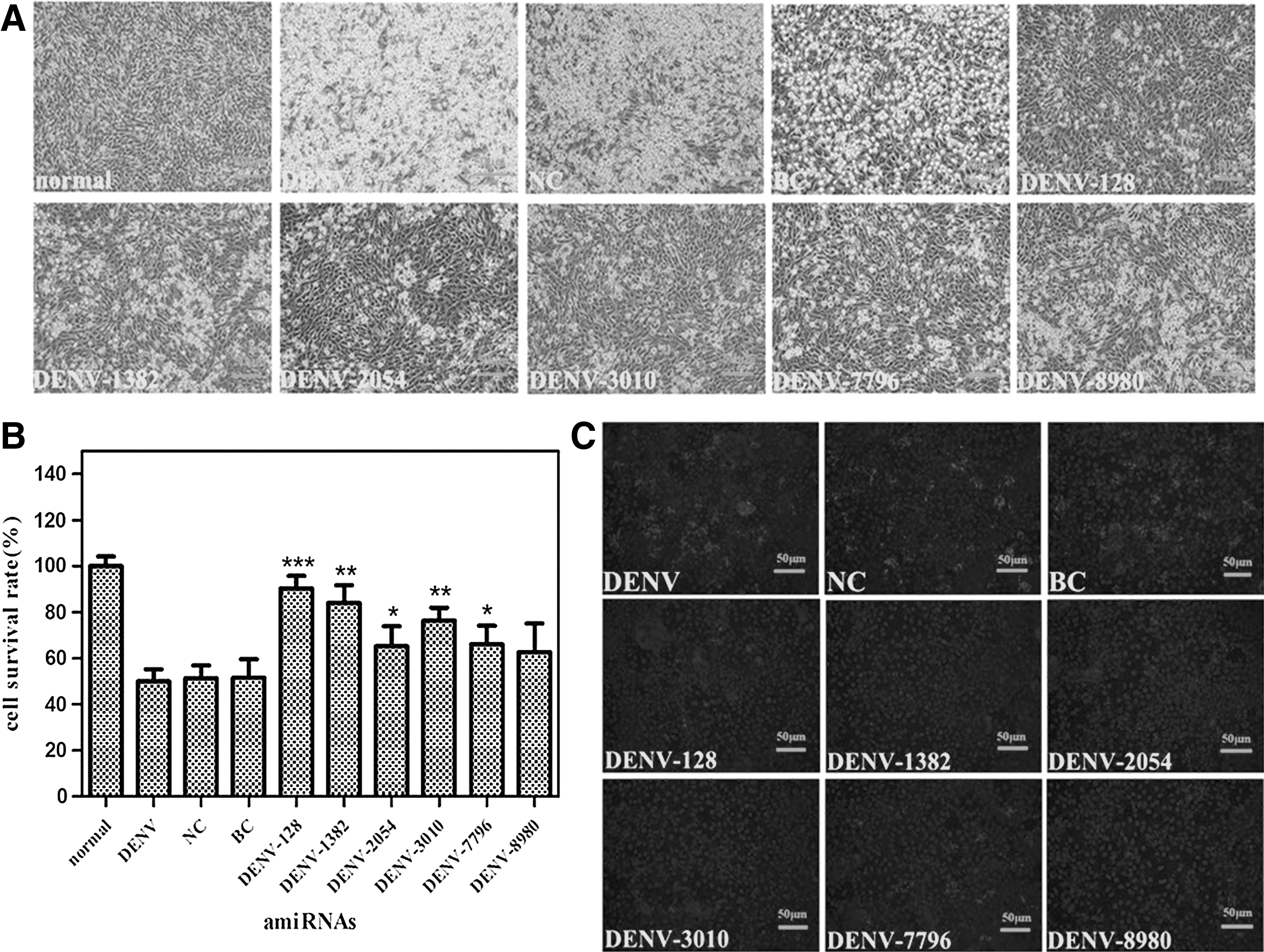
Effects of 6 single amiRNAs by transient transfection on cell viability and DENV-2 replication. Cells seeded in a 96-well plate were transfected with 0.125 μg of single amiRNA per well, then the cells were infected with DENV at multiplicity of infection (MOI) 0.2 one day post-transfection.
Antiviral activity of transiently transfected amiRNAs polycistrons
Cells were challenged with DENV at MOI 0.2 after transfection with 0.125 μg of the tandem amiRNAs vector per well for 24 hours. The morphological changes in the cells were examined by microscope 96 hours post-infection (Fig. 3A), and cell viability assays were employed to analyze the cell proliferation conditions by CCK-8 at 5 days after DENV infection (Fig. 3B). Most of the cells were alive compared with the normal control [noninfected baby hamster kidney cells (BHK-21) cells without the aimRNA], and approximately 50% of cells were killed in the NC, BC, and the virus infection control. The E protein expression of DENV-2 was detected by indirect immunofluorescence 48 hours post-infection and was significantly reduced in amiRNAs-treated groups compared with the NC, BC, and the virus infection control (Fig. 3C). These results showed that DENV128-1382 was the most effective amiRNA combination for DENV inhibition.
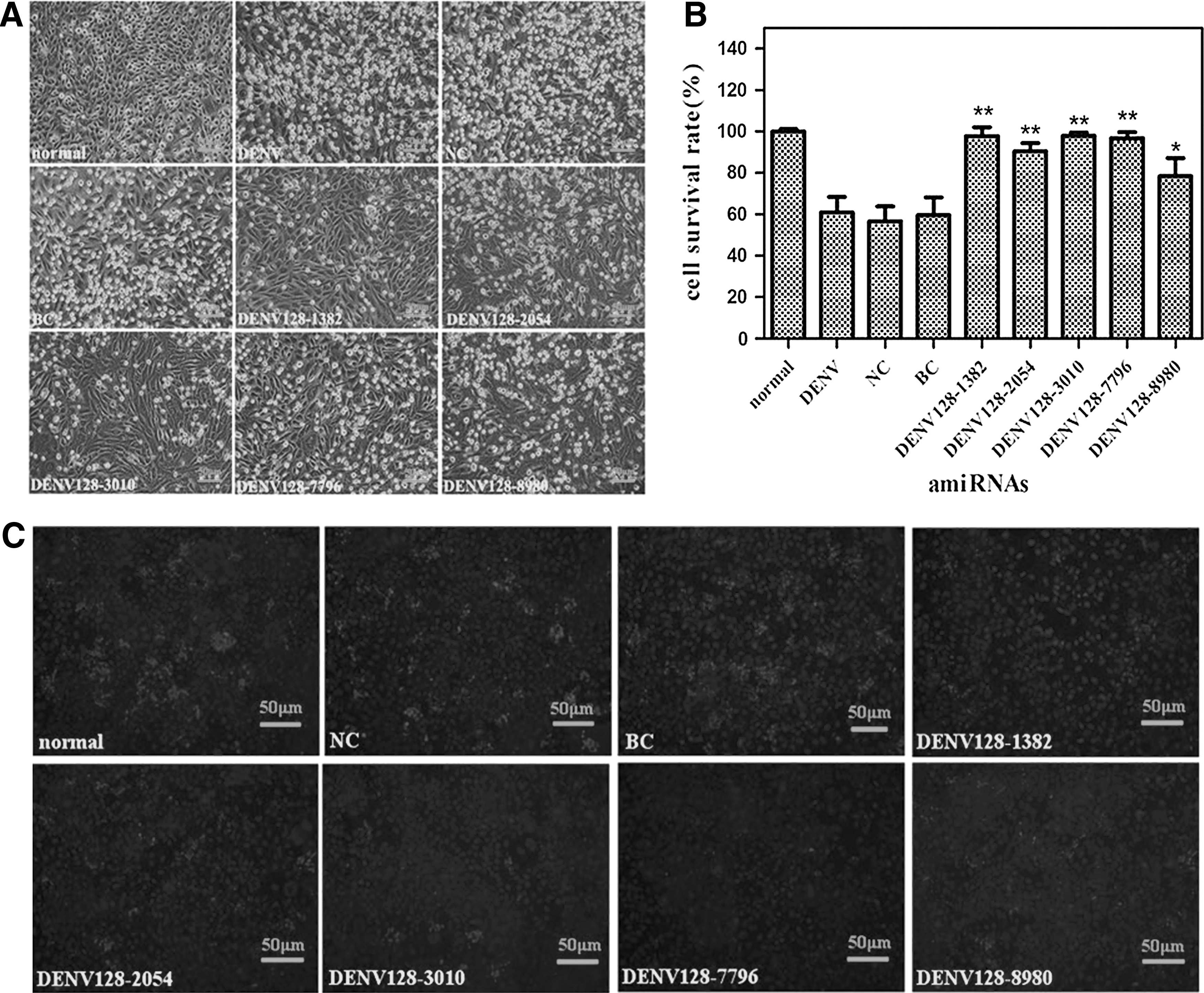
Effects on cell viability and DENV-2 replication by transiently transfected tandem amiRNAs polycistrons. Cells seeded in a 96-well plate were transfected with 0.125 μg miRNA-like polycistrons per well, then the cells were infected with DENV at MOI 0.2 one day post-transfection.
Antiviral activity of the lentivector expressing amiRNAs
We built lentivectors stably and serially expressing DENV-128, DENV128-1382, NC, and BC to diminish the variables associated with transient transfection. DENV-128 was the most effective single amiRNA, and DENV128-1382 was the most effective amiRNA combination. Lentiviral titers were calculated by fluorescence expressing cells (Jiang et al., 2003). BHK-21 cells were seeded in 96-well plates and infected with the amiRNA-expressing lentiviruses at a range of different MOIs. The cells were challenged with DENV at MOI 0.2 three days after lentiviral transduction. Cell viability assay was utilized to analyze the cell proliferation conditions by CCK-8 at 5 days after DENV infection. For the DENV-128 and DENV128-1382 treated cell groups, most of the cells infected with DENV were alive compared with the NC, BC, and virus infection control, for which over 60% of cells were dead. The cell survival rate of pLNB-MP-DENV128-1382 (MOI=1) was 3.1-fold higher than the virus infection control (Fig. 4A). The culture supernatants were collected 48 hours post-infection for subsequent viral plaque assay analysis. The viral titers of pLNB-MP-DENV128-1382 (MOI=1) group dropped drastically to 9.5×103 PFU/mL, while the viral titer l of the virus infection control was 8.5×104 PFU/mL (Fig. 4B, C). Simultaneously, DENV RNA and E protein levels in the DENV-128 and DENV128-1382 stable expressing groups were reduced remarkably compared with the BC, NC, and virus infection control (Fig. 4D, E, F).

Effects on cell viability and DENV-2 replication by lentiviral-expressed amiRNAs.
These results of cell viability, viral titer, viral RNA, and E protein expression indicated that the lentiviral expressing amiRNAs DENV-128 and DENV128-1382 were effective agents for DENV-2 replication. In addition, we concluded that the antiviral activity of these amiRNAs was sequence-specific from the results of endogenous IFN response experiment (Fig. 4G). However, mRNA expression of OAS1 gene in amiRNA groups increased to some extent compared with the normal control but was identical with the BC group (no amiRNA expression).
Discussion
MicroRNA induced RNA silencing has generally become an effective tool to study gene functions (Chung et al., 2006; Schwab et al., 2006). Previous studies showed that a synthetic siRNA could capably inhibit representative DENV strains replication in cell cultures and prevent pathogenic progression in a murine model of severe dengue disease (Stein et al., 2011). In this paper, amiRNA induced RNA silencing was utilized to suppress DENV-2 replication and expression. These 21 amiRNAs targeting DENV genes were constructed into pOK-basic vectors respectively based on M. musculus miR-155 backbone. There were 6 single-amiRNA expression plasmids that proved to be effective, but to different extents. By the transient transfection, DENV-128 is the most effective in inhibiting DENV replication among the 6 single amiRNAs.
DENV-128 outshined other DENV strains by the antiviral experiments among a series of amiRNAs designed against the highly conserved regions in the genomes of genetically various DENV. In view of the DENV-128 targeting region, we conclude that DENV-128 targeted the 5′CS region of DENV genome. Carter et al. earlier reported that anti-DENV group 1 trans-splicing introns (αDENV-Grp1s) targeting the 5′CS region repressed virus replication efficiently (Carter et al., 2010). David et al. also showed that the designed siRNAs inhibited West Nile virus moderately and yellow fever virus potently by targeting their 5′CS regions (Stein et al., 2011). Korrapati et al. showed that a siRNA targeting a 5′NTR conserved sequence across multiple DENV serotypes suppressed DENV infection efficiently (Korrapati et al., 2012). The structure of the 5′CS region has been known to be indispensable for DENV replication (Alvarez et al., 2008). Overall, these researches demonstrated that the flavivirus 5′CS domain is sensible to RNAi; therefore, it offers an attractive target for improvements of nucleic acid inhibitors for flavivirus.
In theory, amiRNA, targeting against a region that is highly conserved across the virus genomes of the 4 DENV serotypes would inhibit representative strains of DENV efficiently. However, in our study, some amiRNAs, such as DENV-10662 that targeted against the highly conserved 3′UTR region, were consistently inactive (data not shown). A possible explanation for this inefficiency could be steric blockade, barrier of RNA-binding protein, or the target complex secondary structure. Xu and colleagues showed that shRNA silencing efficacy was strongly affected by the secondary structure of the target RNA site (Tan et al., 2012) and the 3′-terminal region of the DENV genome indeed forms RNA secondary structure (Gebhard et al., 2011). This indicated that target RNA secondary structure is a key determining factor for the amiRNA silencing efficacy.
The transient transfection of single amiRNAs could inhibit the replication of DENV to a certain extent. However, these single amiRNAs could not be sufficient to prevent from DENV infection owing to RNAi resistant escape mutants of DENV. Just as miRNAs could transcribe from polycistronic pri-miRNAs, a reconstructed expressing vector could also express 2 or more different amiRNAs from a single transcript, which has the potential to inhibit 2 or more different target sequences effectively. Liu et al. showed that the miRNA polycistron could express functional mature miRNAs that inhibited HIV-1 replication efficiently and selectively (Liu et al., 2008). Our miRNA-like polycistron constructs, all of which contained DENV-128, were screened by primary experiment. It was shown that the combination of DENV128 and DENV1382 was the most effective one.
Furthermore, the antiviral effects of amiRNAs may be limited for low transfection efficiency. Indeed, the most challenging trouble in RNAi application is the delivery system. Lentiviral vectors are efficient vehicles for gene stable transfer. Numerous developments have been made in vector design to raise transgene expression efficiency and biosafety, and these advancements bring about the agreement of lentiviral vectors for use in clinical researches (Liu et al., 2013). Therefore, the lentiviral vector-based delivery strategy was applied in this study. We constructed lentiviral vectors expressing the representative amiRNA transcripts to validate their inhibitory effects further. From the results of cell viability, plaque assay, and E protein analysis, we confirmed that the selected amiRNAs could effectively inhibit the replication of DENV. But in the plaque assay analysis, the viral titers of NC and BC groups were also reduced compared with the virus infection control (Fig. 4C). In the interferon induction assay, the mRNA expression of OAS1 in amiRNA groups increased to some extent compared with the normal control but was identical with the BC group (no amiRNA expression). So we presumed that the second-generation lentiviral vector itself might have OAS1 induction potency and disturb DENV replication although more evidence should be demonstrated (Jackson and Linsley, 2004; Kenworthy et al., 2009). Also, we noticed that the MP128 (MOI 0.5) lowered viral titers much more effectively than the MP128-1382 (MOI 0.5) in the single versus polycistronic amiRNA functional study (Fig. 4C). The reason may be that each amiRNA generation rate in the polycistronic amiRNAs was reduced than the single amiRNA (Israsena et al., 2009; Zhou et al., 2005). Although the DENV inhibition efficiency of MP128-1382 was not enhanced more than the single MP128 amiRNA, the tandem amiRNA were more practical for the virus variation and mutant escape strains.
In conclusion, a further progression in the improvement of amiRNA inhibitors for DENV infection was provided here. These novel amiRNAs expressed from a single miRNA-like polycistron provide us a striking alternative way for the antiviral or transgenic suppression of DENV in future.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China 81202562 and the Chinese Science and Technology Key Projects grants 2012ZX10004503-011 and 2012ZX09103301-044.
Author Disclosure Statement
No competing financial interests exist.