
Abstract
Our current understanding of the molecular events that functionally characterize cellular biology continues to be revised. Recent observations find that the vast majority of the human genome is transcribed and may be functionally relevant. Many of these previously unrecognized transcripts, both short and long non-coding RNAs, have been found to be active modulators of protein coding gene function. While such observations were in the past relegated to imprinted genes, it is now becoming apparent that several different genes in differentiated cells may be under some form of non-coding RNA based regulatory control. Emerging evidence suggests that some of these long non-coding RNAs are functional in controlling gene transcription by the targeted recruitment of epigenetic silencing complexes to homology-containing loci in the genome. Most notably when these repressor non-coding RNAs are targeted using small RNA-based inhibitors (such as with RNA interference), a de-repression of the targeted gene can occur resulting in activation of gene expression. Knowledge of this emerging RNA based epigenetic regulatory network has implications not only in cellular evolution but also for the development of an entirely new area of pharmacology.
Introduction
Evidence addressing this question of what ncRNAs are doing in the cell has begun to emerge with the notion that they are functionally relevant moieties, sometimes involved in the regulation of gene transcription (reviewed in Morris and Vogt, 2010). Interestingly, some of the recent observations that long non-coding RNAs (lncRNAs) are involved in the regulation of gene transcription arise from earlier studies in RNA interference (RNAi) and the paradigm of short RNA biology.
Small RNAs and the regulation of gene transcription
RNAi is best known for the work demonstrating that ∼22 nucleotide small interfering RNA (siRNAs) or microRNAs (miRNAs) could be used to functionally target and direct the degradation of a cognate messenger RNAs (reviewed in Hannon and Rossi, 2004). However, guide strands derived from siRNAs have also been observed to direct transcriptional gene silencing (TGS), via the targeting of particular chromatin/epigenetic remodeling proteins to the targeted loci (reviewed in Green and Weinberg, 2011; Turner and MORRIS, 2010). While much has been gleaned from studies whereby siRNAs were found to direct TGS, until recently it was unknown whether human cells generate endogenous TGS-driving siRNAs or whether there is an endogenous pathway in human cells actively utilizing TGS to control transcription.
Uncovering the endogenous mechanism of non-coding RNA based epigenetic regulation in human cells
During the course of our work detailing how siRNAs were capable of directing TGS in human cells, one striking observation was generated: we discovered, quite serendipitously, that only the antisense strand of the siRNA was required to drive TGS in human cells (Weinberg et al., 2006). Other later studies have validated this requirement (Turner et al., 2009), suggesting that the endogenous mechanism in human cells whereby ncRNAs regulate gene transcription may involve forms of RNA beyond double stranded RNAs.
Despite earlier observations indicating that de novo derived siRNAs could affect gene transcription, little was known as to whether or not there were active ncRNAs modulating gene transcription in human cells. Several studies provided evidence that miRNAs could function as endogenous regulators of gene transcription (Omoto et al., 2004; Omoto and Fujii, 2005; Klase et al., 2007; Kim et al., 2008; Tan et al., 2009). One recent study revealed that many aspects of the hematopoietic system are in fact under some level of miRNA directed TGS (Zardo et al., 2012). The vast majority of miRNA studies however point to a predominant role for these small RNAs in controlling genes in a post-transcriptional manner.
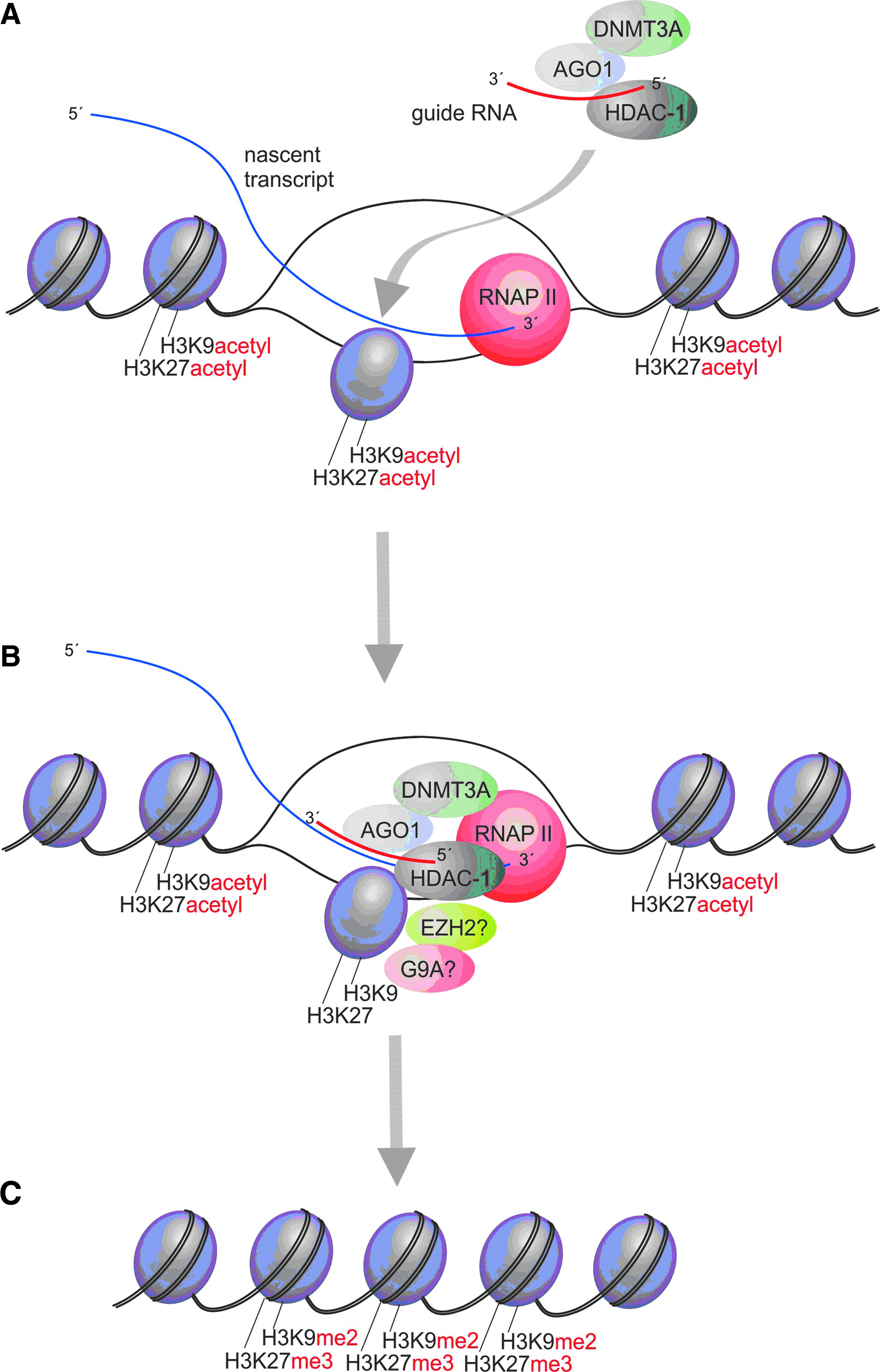
Hints to a deeper layer of RNA-based transcriptional regulation arose quite unexpectedly from studies that set out to target de novo derived siRNAs to the p21 and E-cadherin promoters, specifically targeting AT-rich regions in these promoters (Li et al., 2006; Place et al., 2008). Notably, AT-rich regions are generally found in the coding bodies of genes, while promoters tend to exhibit CpG islands and palindromic sequences with high GC content, oftentimes used for transcription factor or protein binding (Alberts, 1994). Interestingly, p21 and E-cadherin promoter targeted siRNAs were actually able to activate gene expression, although activation of transcription was not demonstrated (for example, as measured by nuclear run-on analysis) (Li et al., 2006; Place et al., 2008). These studies also determined that Argonaute 2 (AGO2) is required for activation, which is a noteworthy because siRNAs that modulate TGS had generally been observed to require the action of Argonaute 1, DNA methyltransferase 3a, and histone deacetylase 1 (reviewed in detail in MORRIS, 2009a) (Fig. 1). Though some investigators had previously observed a role for AGO2 in siRNA-directed TGS, these studies specifically targeted siRNAs to the TATAA start site in the candidate promoters and appeared to use a mechanism independent of targeted epigenetic modifications (Janowski et al., 2006; Napoli et al., 2009)—that is, the obstruction of RNA Polymerase II (RNAPII) activity by direct binding or via a DNA/RNA interaction at the TATAA sequence (discussed in MORRIS, 2009a; Green and Weinberg, 2011).
Antisense RNA mediated regulation of gene transcription.
How do siRNAs targeted to AT-rich regions in the p21 and E-cadherin promoters utilize AGO2 to activate gene expression? Building on the work carried out by Li, Place, and colleagues (Li et al., 2006; Place et al., 2008), we addressed this question by determining that the activating siRNAs targeted to AT-rich promoter sites weren't actually targeting the promoter, but rather, previously undiscovered antisense lncRNAs (Morris et al., 2008). These lncRNAs overlapped the p21 or E-cadherin coding regions. This study found that these antisense lncRNAs were actively involved in regulating the transcriptional expression of p21 and E-cadherin (Morris et al., 2008). Other genes, such as the tumor suppressor gene p15 and the transcription factor Oct4, also appeared to be under this mode of antisense lncRNA based regulation (Yu et al., 2008; Hawkins and MORRIS, 2010). Interestingly, the regulation of Oct4 transcription appeared to be via the action of a pseudogene-expressed lncRNA (Hawkins and MORRIS, 2010). The latter represents the first such example of pseudogenes epigenetically regulating their protein-coding counterpart.
A model has begun to emerge, based on several different bodies of work, suggesting that antisense lncRNAs are active regulators of gene transcription in human cells (Fig. 2). Mechanistically, this model suggests that the siRNA targeting and degradation of the gene-specific antisense lncRNAs, affects the ability of these RNAs to interact and guide epigenetic silencing complexes to their particular targeted loci in the promoter of the protein coding gene, in this case p21, p15, or E-cadherin. The result of this unintended lncRNA targeting is the de-repression or activation of the lncRNA-targeted gene (Fig. 2). It is in essence “suppressing the endogenous suppressor,” which results in the activation or a loss of endogenous epigenetic brakes being instilled at the particular lncRNA targeted gene (Fig. 2 and reviewed in MORRIS, 2011).
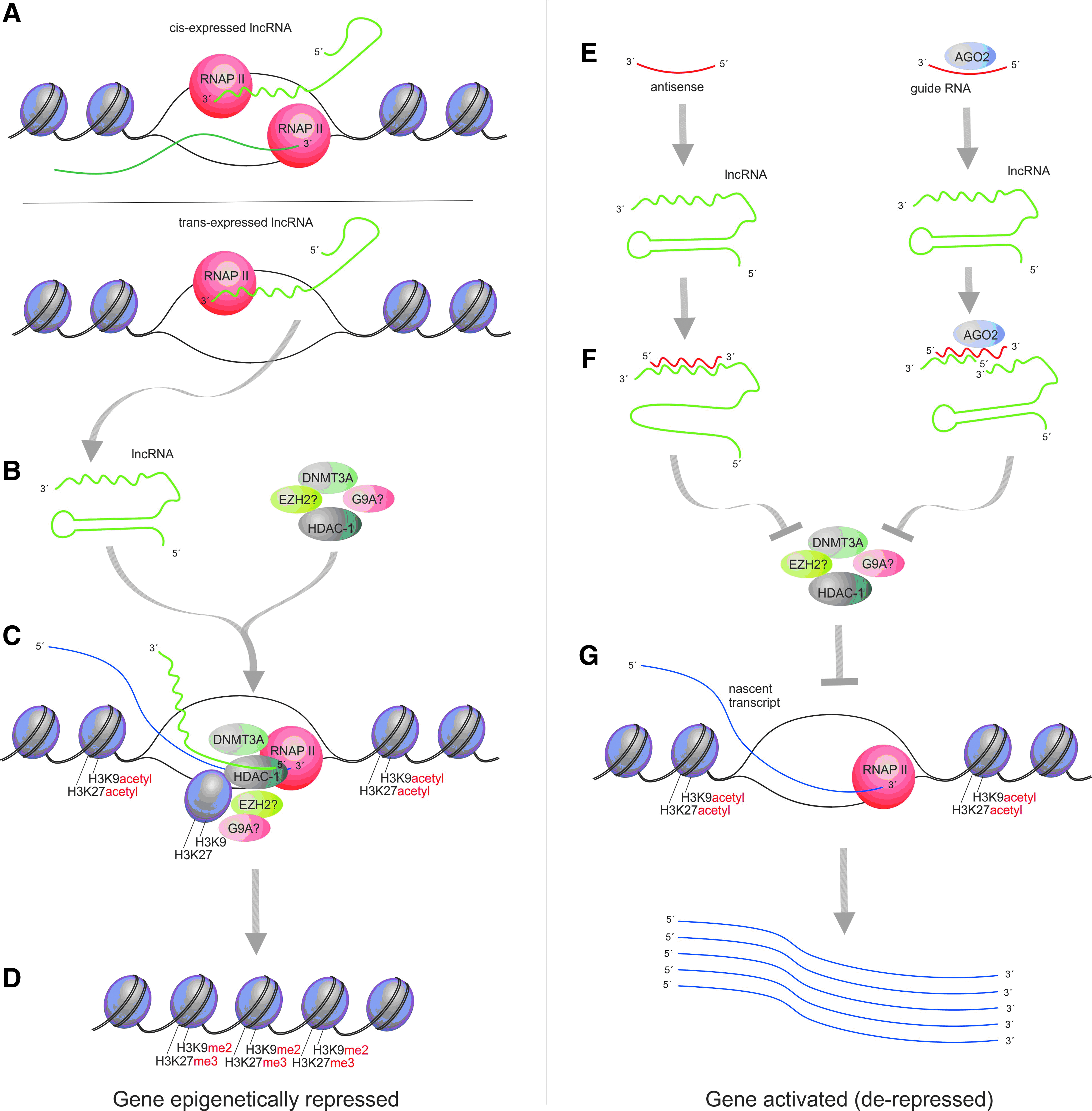
Antisense non-coding RNA mediated transcriptional silencing and activation.
The list of functional lncRNAs in human cells continues to expand with antisense lncRNAs having a regulator role in several different genes, ranging from genes associated with cancer (Morris et al., 2008; Yu et al., 2008; Tahira et al., 2011), autism (Kerin et al., 2012), and neurodegenerative disease (Lipovich et al., 2012). Recently, the targeting of a brain-derived neurotrophic factor (BDNF)-associated antisense lncRNA results in physiologically relevant changes in vivo (Modarresi et al., 2012). In this study Modarresi and colleagues found that targeting of the BDNF-associated antisense lncRNA using chemically modified oligonucleotides or siRNAs resulted in 2- to 7- fold–increased expression of BDNF in the mouse brain. Importantly, this increased BDNF expression had a direct effect on downstream BDNF targets (Modarresi et al., 2012). Such observations suggest that the juxtaposition of knowledge of the endogenous lncRNA TGS pathway with siRNA targeting can lead to applications of targeted gene activation/de-repression.
These collective observations—that lncRNAs are involved in regulating gene expression in differentiated cells—adds to the growing complexity of known lncRNAs involved in cellular regulation. Examples include the antisense Xist lncRNA TSIX, which is involved in dosage compensation and X-inactivation in undifferentiated primordial cells (reviewed in LEE, 2009). Other examples of lncRNAs involved in epigenetic regulation can be found in the imprinting genes (reviewed in Latos and Barlow, 2009). These lncRNAs also appear to function by targeting the recruitment of different epigenetic regulatory complexes to their intended targets. Clearly, the emerging evidence suggests that in human cells many more genes than previously envisioned, beyond X-inactivation and imprinting related genes, might actually be regulated by lncRNAs. In fact, lncRNAs might be actively switching on and off genes in an orchestral regulation that governs the fidelity of the cell and functions in cellular adaption (discussed in (MORRIS, 2009a, 2009b). Furthermore, a possible, underappreciated function for miRNAs might also be that they function as active regulators of antisense lncRNAs by sensing the ratios of sense to antisense compliments and adjusting the efficacy of antisense lncRNA targeting by interacting with the antisense RNAs in a manner that is suppressive of their targeting function (discussed in MORRIS, 2009a). Supportive of miRNA regulation of lncRNAs is the observation that double-stranded RNAs can actually target both sense and antisense strands (Wei et al., 2009), and maybe the reason miRNAs are abundantly expressed and linked to several forms of cancer in human cells is due to their role in regulating bidirectionally transcribed discordantly regulated loci (discussed in detail in MORRIS, 2009b). As the majority of those discordantly regulated bidirectionally transcribed genes detected and/or characterized to date appear to be involved in cell cycle, the loss of which can result in oncogenic transformation, it is not too far of a stretch to envision a role for miRNAs in regulating antisense lncRNAs.
Conclusion
Over the last decade, a lot of interest has been generated around RNAi and the ability of double-stranded RNAs [siRNAs, short hairpin RNA (shRNAs), and miRNAs] regulate gene expression in a post-transcriptional manner (Tomari and Zamore, 2005). The observation that double stranded RNAs can modulate messenger RNA expression post-transcriptionally is no doubt a paradigm shift. However, over the last ∼2 years it has become increasingly apparent that RNAi is only one part of a much larger story with regards to RNA-based regulation in human cells. Studies carried out to discern how siRNAs are able to modulate gene transcription in human cells have led to insights suggesting that antisense lncRNAs can transcriptionally regulate gene expression via the specific targeting of epigenetic silencing complexes to loci in the genome.
One might ask, why should these relatively low-expressed natural antisense transcripts be of any interest? The answer to this question lies in the fact that antisense lncRNAs appear in some instances to regulate gene transcription via the targeting of epigenetic changes to particular loci in the genome. This is noteworthy as for over ∼4 decades epigenetic changes to the genome have been observed in various cell types and cancers with little understanding as to what is guiding these complexes to their respective target loci. Epigenetics has been mechanistically implicated in cancer, but the mechanism of action for directing epigenetic changes has for the most part been lacking. The emerging observations with antisense lncRNAs playing a role in guiding epigenetic silencing complexes to target loci is suggestive of a mechanism whereby epigenetic silencing complexes are targeted to particular loci in the genome. These findings open up an entirely new discipline of scientific inquiry. This new area of inquiry has immense potential with regards to targeted interventions aimed at controlling the epigenetic state of particular genes. The future impact of this to novel therapeutics is immense.
Footnotes
Acknowledgments
This project is funded by National Institute of Allergy and Infectious Diseases R56 AI096861-01 and P01 AI099783-01 to Kevin V. Morris and National Cancer Institute R01 CA151574, National Institutes of Health R01 CA153124 to Peter K. Vogt.
Author Disclosure Statement
No competing financial interests exist.