
Abstract
ISIS 481464 is a constrained ethyl (cEt) modified phosphorothioate antisense oligonucleotide (ASO) targeting signal transducer and activator of transcription 3 (STAT3) studied in mice and monkey to support oncology clinical trials. Six-week toxicology studies were performed in mice and cynomolgus monkey (up to 70 and 30 mg/kg/week respectively). Reduction in STAT3 protein up to 90% of control was observed in monkey. Cynomolgus monkey was considered the most relevant species to human with respect to pharmacokinetic properties, but mice are useful in their relative sensitivity to the potential proinflammatory and hepatic effects of oligonucleotides. In monkeys, there was no impact on organ function at doses up to 30 mg/kg/week for 6 weeks. Minimal to slight proximal tubular epithelial cell degeneration and regeneration within the kidney was observed, which had no impact on renal function and showed reversibility at the end of the treatment-free period. Additionally, mild and transient activated partial thromboplastin time elevations and mild increases in complement Bb were observed at the higher doses by intravenous dosing only. In mice, the alterations at 70 mg/kg/week included spleen weight increase up to 1.4-fold relative to control, increases in alanine aminotransferase and aspartate aminotransferase up to 1.8-fold over control, interleukin-10 increases up to 3.7-fold, and monocyte chemoattractant protein-1 increase up to 1.9-fold over control. No significant clinical pathology or histopathology changes were seen in mice at 20 mg/kg/week or less. The toxicity profile of ISIS 481464 is consistent with effects observed with phosphorothioate ASOs containing 2′-O-methoxyethylribose modifications instead of cEt.
Introduction
One target for antisense therapy in the treatment of cancer is signal transducer and activator of transcription 3 (STAT3), which belongs to the STAT family of transcription factors. STAT3 is considered to be a promising cancer drug target because of its pleiotropic involvement in tumorigenesis. STAT3 not only regulates the expression of many genes directly important to the survival of tumor cells, but STAT3 is also an important factor in non-tumor cells of the tumor microenvironment involved in immune evasion of tumor cells, angiogenesis, and metastasis (Alvarez et al., 2005).
ISIS 481464 is a phosphorothioate-modified chimeric (P=S) ASO with 10 bases in the center that are DNA and thus supports RNAse H-mediated metabolism, flanked by 3 constrained ethyl (cEt)-modified nucleosides on both the 5′ and 3′ ends (Kortylewski et al., 2009; Seth et al., 2009). This chimeric design is called an cEt gapmer. The novel cEt chemical modification incorporated into ISIS 481464 is anticipated to confer similar pharmacokinetic properties to the compound as 2′-O-methoxyethyl (2′-MOE) chemical modifications have to other members of the class, but with a potential for enhanced potency.
ISIS 481464 is designed to target and reduce levels of human STAT3 messenger RNA (mRNA). The antiproliferative effects of ISIS 481464 were demonstrated in human tumor cells with concurrent decreases specifically in both STAT3 mRNA and protein levels in various xenograft models (Kim and MacLeod, in preparation).
Most toxicities induced by phosphorothioate ASO are generally independent of the specific sequence or the molecular target and are considered to be related to the class of compound (Henry et al., 2007). These toxicities include activation of complement in monkey (Henry et al., 1997b; Henry et al., 2000a; Henry et al., 2002) and inhibition of intrinsic clotting times (Henry et al., 1997c), lymphoproliferation in mice (Monteith et al., 1997; Henry et al., 2000b; Senn et al., 2005), and varying degrees of target organ toxicity in the kidney and liver due to the accumulation of drug in certain cell types (Henry et al., 1997a; Henry et al., 1997d; Monteith et al., 1998; Monteith et al., 1999). These effects typically occur at doses substantially greater than the doses employed in clinical trials.
In the first safety studies of a cEt-modified ASO, the toxicity of ISIS 481464 was assessed in cynomolgus monkeys and mice for 6 weeks. Mice were administered intravenous (IV) doses of ISIS 481464 up to 70 mg/kg/week and monkeys were administered IV and subcutaneous (SC) doses of ISIS 481464 as high as 30 mg/kg/week. Because of species differences in gene sequences, ISIS 481464 targeting human STAT3 is inactive in mouse. In contrast, the cynomolgus monkey sequence for STAT3 is fully homologous to human STAT3, allowing for the full characterization of the potential for human toxicity, including possible toxicologic effect and exaggerated pharmacology associated with profound decreased in STAT3 expression. In addition, the pharmacokinetics and tissue distribution were characterized in these studies to relate exposure (tissue and plasma concentrations) with toxicity.
Materials and Methods
Oligonucleotides
ISIS 481464 is the pentadecasodium salt of a 16-base residue (16-mer) phosphorothioate oligonucleotide (Fig. 1). Each of the 15 internucleotide linkages is a 3′-O to 5′-O phosphorothioate diester. Ten of the sixteen sugar residues are 2-deoxy-D-ribose, and the remainder are 2-O, 4-C-[(S)-ethylidene]-D-ribose (constrained ethyl, or cEt). The residues are arranged so that there are three cEt nucleosides at the 5′ and 3′-ends of the molecule flanking a gap of ten 2′-deoxynucleosides. All of the cytosine bases are methylated at the 5′-position.
Chemical structure of ISIS 481464.
ISIS 481464 can be named by accepted oligonucleotide nomenclature, showing each 3′-O to 5′-O-linked phosphorothioate diester internucleotide linkage as follows: 2′-O, 4′-C-[(S)-ethylidene]-5-methyl-P-thiocytidylyl-(3′-O→5′-O)-2′-O, 4′-C-[(S)-ethylidene]-5-methyl-P-thiouridylyl-(3′-O→5′-O)-2′-O, 4′-C-[(S)-ethylidene]-P-thioadenylyl-(3′-O→5′-O)-P-thiothymidylyl-(3′-O→5′-O)-P-thiothymidylyl-(3′-O→5′-O)-P-thiothymidylyl-(3′-O→5′-O)-2′-deoxy-P-thioguanylyl-(3′-O→5′-O)-2′-deoxy-5-thioguanylyl-(3′-O→5′-O)-2′-deoxy-P-thioadenylyl-(3′-O→5′-O)-P-thiothymidylyl-(3′-O→5′-O)-2′-deoxy-P-thioguanylyl-(3′-O→5′-O)-P-thiothymidylyl-(3′-O→5′-O)-2′-deoxy-5-methyl-P-thiocytidylyl-(3′-O→5′-O)-2′-O, 4′-C-[(S)-ethylidene]-P-thioadenylyl-(3′-O→5′-O)-2′-O, 4′-C-[(S)-ethylidene]-P-thioguanylyl-(3′-O→5′-O)-2′-O, 4′-C-[(S)-ethylidene]-5-methylcytidine, 15 sodium salt.
Toxicology
Mouse
CD1 mice were purchased from Orient Bio Inc. Mice were acclimatized for a minimum of 6 days and were approximately 6 weeks old prior to pretreatment. Groups of 10 or 16 male and 10 or 16 female mice were administered 0 (vehicle control), 2, 6, 20, and 70 mg/kg/week by IV injection on days 1, 3, 5, and 7 (loading dose phase) followed by once weekly dosing thereafter, on days 14, 21, 35, and 42 (males) or 43 (females). On day 44 (males) or day 45 (females), 20 mice (10 per sex) from each toxicology group were necropsied 48 hours after final dosing (Table 1). The reversibility of any treatment-related effects was evaluated following 6 weeks of recovery (day 84) at the 0, 20, and 70 mg/kg/week dose levels of ISIS 481464 (6 animals/sex/group). A standard toxicological evaluation was performed, which included daily clinical signs, weekly body weight, and food consumption, and end of study ophthalmic exams, clinical pathology, organ weights, cytokine analysis, and complete gross necropsy on all animals and light microscopic examination of a full range of tissues.
PBS vehicle control or ISIS 481464 will be administered intravenously (bolus) on Days 1, 3, 5, and 7 and weekly thereafter [on Days 14, 21, 28, 35, and 42 (males) / 43 (females)].
Groups 6 and 7 will be sacrificed at various time points (3 animals per time point).
F, females; M, males; PBS, phosphate buffered saline.
Monkey
Cynomolgus monkeys were obtained from Nafovanny. The potential toxicity and toxicokinetic profile of ISIS 481464 was examined following 6 weeks of treatment (day 44) by 1-hour IV infusion or SC injection on days 1, 3, 5, and 7 (loading dose) followed by once-weekly administration thereafter. Fifty-six naïve cynomolgus monkeys (28 males and 28 females) were assigned to 6 treatment groups (Table 2). The reversibility of any effects of treatment with ISIS 481464 was evaluated in a subset of monkeys assigned to 8 weeks of recovery (day 98) at the 0 and 30 mg/kg/week (IV and SC) dose levels of ISIS 481464 (2 animals/sex/group).
Groups 1–6: days 1, 3, 5, and 7, then weekly thereafter (days 14, 21, 28, 35, and 42).
Groups 1–4; 1-hour intravenous infusion followed by subcutaneous dosing.
Toxicokinetic analysis group.
One animal per sex necropsy on day 3 (this animal will not be dosed prior to necropsy), 9, 16, 30, 44, 58, and 72.
IV, 1-hour intravenous infusion; SC, subcutaneous injection.
In-life observations for animals assigned to each toxicology group included evaluations for mortality, clinical signs, body weight, food consumption, clinical pathology indices (hematology, coagulation, serum chemistry, urinalysis, and urine chemistry parameters), ophthalmology, electrocardiography, and complement activation (C3 and Bb split products). Post-mortem evaluations for these animals included organ weights as well as macroscopic and microscopic evaluations for all surviving animals. In addition, liver samples were also collected at terminal and recovery necropsies for STAT3 mRNA and protein analyses. Plasma, tissue, and urine samples for toxicokinetic analysis were collected from all treatment groups at various time points after single and repeated administration for ISIS 481464 concentration analyses.
RNA purification and quantitative real-time polymerase chain reaction analysis of STAT3
Approximately 100 mg of frozen liver tissue was homogenized in 3 mL of RLT lysis buffer (Qiagen) supplemented with 1% 2-mecaptoethanol (Sigma-Aldrich). The resulting homogenate was applied to Qiagen RNeasy 96-well plate for RNA purification according to manufacturer's protocols. After purification, the RNA samples were subjected to quantitative real-time polymerase chain reaction (qRT-PCR) analysis. The Perkin-Elmer ABI Prism 7700 Sequence Detection System, which uses real-time fluorescence RT-PCR detection, was used to quantify STAT3 mRNA. RNA transcripts were normalized to total RNA levels using either cyclophilin A or RiboGreen (Life Technologies). The sequences used for qRT-PCR include: STAT3: 5′-AAGTTTATCTGTGTGACACCAACGA-3′ (forward), 5′-CTTCACCATTATTTCCAAACTGCAT (reverse), 5′-FAM-TGCCGATGTCCCCCCGCA-TAMRA-3′ (probe); Cyclophilin A: 5′-TGCTGGACCCAACACAAATG-3′ (forward), 5′-TGCCATCCAACCACTCAGTC-3′ (reverse), 5′-FAM-TTCCCAGTTTTTCATCTGCACTGCCA-TAMRA-3′ (probe).
Western blot analysis of STAT3 protein
Frozen liver tissues (∼100 mg) were homogenized in 1 mL of ice-cold radioimmunoprecipitation assay buffer buffer (Sigma) containing inhibitor cocktails of both proteases and phosphatases (Roche Diagnostics). Total lysates were separated by 4%–12% Bis-Tris polyacrylamide gel electrophoresis (Life Technologies), transferred to a polyvinylidene difluoride membrane, and immunoblotted using the primary antibodies for STAT3 (Cell Signaling) and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Advanced Immunochemicals). Immunospecific bands were detected with the Enhanced Chemiluminescence Plus detection kit (GE Healthcare) and quantified using ImageJ software.
Cytokine and chemokine assay
Plasma was collected from mice by cardiac puncture. Interleukin (IL)-1β, IL-6, IL-10, and monocyte chemoattractant protein-1 (MCP-1) levels in the plasma were analyzed using a custom, multiplex immunosorbent assay according to the manufacturer's protocol (Meso Scale Discovery). Multiplex plates precoated with capture antibodies for specific analytes were presoaked/blocked with calibrator/sample diluent for 30 minutes prior to use. Diluted standards (4 replicates) and samples in duplicate were added to the wells and incubated for 2 hours. All incubations were performed at room temperature with vigorous shaking (300–1,000 rpm). Plates were washed three times with phosphate buffered saline + 0.05% Tween-20 before adding a cocktail of Sulfo-Tag® detection antibodies to each well. After incubating with detection antibodies for 2 hours, plates were washed 3 times. Read buffer was added to the wells and the plates immediately imaged using the MSD Sector 2400 imaging system. Data was analyzed using MSD Discovery Workbench® software. Concentrations of all unknown samples were back-calculated using results interpolated from the corresponding standard curve regression using a weighted, 4-parameter fit. Final sample concentrations (pg/mL) were calculated by factoring the dilution factor used for each sample.
Toxicokinetics
Mouse
Plasma and tissue toxicokinetic profiles of ISIS 481464 were evaluated in separate toxicokinetic groups at the 6 and 20 mg/kg/week dose levels (51 males per group, 3 animals per time point; Table 1) following a single dose on day 1 or multiple doses to day 42. Animals assigned to the toxicokinetic groups received loading doses on days 1,3, 5, and 7 followed by a weekly maintenance regimen on days 14, 21, 35, and 42. Blood samples and tissues were collected at selected time points from 15 minutes through 48 hours on day 1 and day 42. Tissue collected and frozen for analysis included liver, kidney, spleen, lung, mesenteric lymph node, heart, testes, and brain. In addition, tissue samples were evaluated from ISIS 481464 concentrations from the main toxicology animals sacrificed 48 hours after the final dose.
Monkey
Plasma and tissue toxicokinetic profiles of ISIS 481464 were evaluated at the 3, 10, and 30 mg/kg/week dose levels (Table 2) following a single dose on day 1 or multiple doses to day 42 after both IV and SC administration. Tissue levels of ISIS481464 were also assessed in animals maintained for an additional 8 weeks of recovery. Exposure to ISIS 481464 was evaluated by determination of plasma toxicokinetics and by ISIS 481464 concentration analysis of selected tissues. Plasma and tissue samples for toxicokinetic analysis were collected from all treatment groups at various time points after single and repeated administration for ISIS 481464 concentration analyses.
Analysis of plasma and tissue sample for oligonucleotide
Plasma samples were analyzed using a quantitative, sensitive hybridization enzyme-linked immunosorbent assay method, which is a variation on the method reported previously (Yu et al., 2002). The assay was validated for precision, accuracy, selectivity, sensitivity, and stability of ISIS 481464 quantitation before analysis of mouse and monkey plasma samples. Plasma sample analyses were conducted at Korea Institute of Toxicology and were performed based on the principles and requirements described in 21 Code of Federal Regulation part 58. The assay was evaluated using synthesized putative shortened oligonucleotide metabolite standards and showed no measurable cross-reactivity, confirming the assays specificity for the parent oligonucleotide. The quantitation range of the assay was determined to be 1.0 to 100 ng/mL with 1.0 ng/mL defined the lower limit of quantification.
Tissue samples were analyzed using a quantitative, sensitive high-pressure liquid chromatography with UV detector (HPLC-UV) method validated for precision, accuracy, selectivity, sensitivity, and stability of ISIS 481464 quantification before analysis of mouse and monkey kidney and liver samples. Tissue sample analyses were also conducted at Korea Institute of Toxicology. Oligonucleotide peaks were detected by UV absorbance at 260 nm. Computer software was used to determine the areas under the oligonucleotide peaks for concentration determination. Good separation between full-length and chain-shortened oligonucleotides was demonstrated. The sensitivity of the HPLC-UV method was acceptable for determination of ISIS 481464 tissue concentrations with a quantification range of 10 to 2000 μg/g for ISIS 481464.
Statistical analysis
The data of in vivo studies are expressed as the mean±standard deviation or±standard error of the mean. Comparisons of in vivo data between 2 groups were analyzed with an unpaired Student's t-test. Multiple comparisons were evaluated with 1-way analysis of variance, followed by a post hoc Dunnett's test with JMP version 9 (SAS Institute). A P<0.05 was considered significant.
ISIS 481464 plasma mean concentration-time data were analyzed by noncompartmental methods using the computer program Phoenix™ WinNonlin®, Version 6.1 (Pharsight Corporation). The tissue disposition half-life (t1/2λz) associated with the apparent terminal elimination phase was calculated using a noncompartmental analysis IV infusion or extravascular input model (Model 200-202; Phoenix™ WinNonlin®) applied to the tissue concentration-time profiles of Group 6 for the purposes of obtaining a generally representative estimate only. The tissue disposition half-life (t1/2λz) values associated with the apparent terminal elimination phase from Groups 4 and 5 were calculated using a noncompartmental analysis model (Model 200-202; Phoenix™ WinNonlin®) applied to the tissue concentrations obtained 48 hours after the last dose and at the end of the recovery period (2 point curve) for the purposes of obtaining a generally representative estimate only. The t1/2λz is calculated by the noncompartmental analysis within Phoenix WinNonlin, as described in the text. To be specific, t1/2λz is calculated as t1/2λz=0.693/k, where k is determined by fitting the concentration v. time data using nonlinear regression to the equation C=C0e^(-kt). C0 is the concentration at time=0, and C is the concentration at time=t. For the terminal elimination half-life, t1/2λz, only the terminal elimination portion of the curve is used.
Results
The dose regimens evaluated were designed to equal or exceed the proposed clinical dosing. The high dose evaluated in these studies was selected based on previous experience with related compounds. In rodents, pro-inflammatory effects are common for phosphorothioate ASOs following 2 weeks of treatment at doses higher than 50 mg/kg (Sarmiento et al., 1994; Monteith et al., 1997). In monkeys, doses used in subchronic studies are limited by effects in kidney, which is the tissue that contains the highest oligonucleotide concentrations of any tissue measured. Doses of similar phosphorothioate oligonucleotides greater ≥40 mg/kg/week have been associated with tubular degeneration and associated functional changes (Henry et al., 2007).
Pharmacology
Following 6 weeks of treatment in monkey, liver STAT3 mRNA levels were decreased to 50% of control in monkeys treated with 30 mg/kg/week ISIS 481464 (Fig. 2A), and STAT3 protein levels in the liver were inhibited by approximately 90% compared with controls at doses ≥10 mg/kg/week ISIS 481464 (Fig. 2B, C). Because ISIS 481464 is not complementary with the mouse STAT3 mRNA binding site, there was no reduction of STAT3 expression in the mouse. The toxicities observed in mice for ISIS 481464 were attributed to general oligonucleotide interactions rather than inhibition of STAT3.
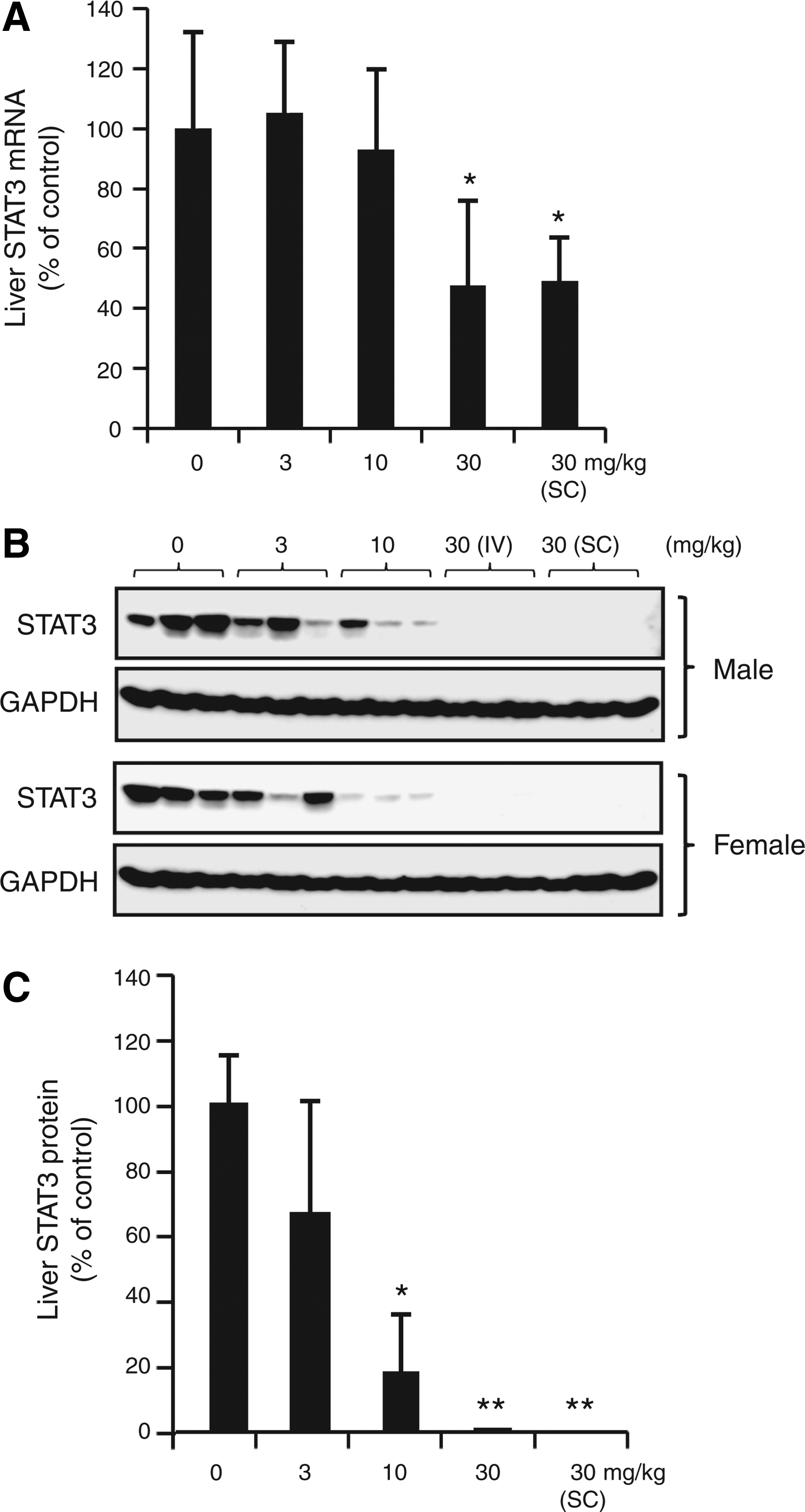
STAT3 levels in the livers of cynomolgus monkeys on day 44.
Toxicology
Clinical signs, organ weights, and gross observations: Mouse
Treatment with ISIS 481464 for 6 weeks at dose levels of up to 70 mg/kg/week was well tolerated in mice. No treatment-related mortality, clinical signs, ophthalmic abnormalities, body weight, or food consumption were observed during the study. The only gross findings were enlarged lymph nodes (mandibular, renal, iliac, and inguinal) observed in few females at 70 mg/kg/week. The enlarged lymph nodes were likely associated with the presence of hypertrophied/vacuolated histiocytes observed microscopically.
Proinflammatory effects of oligonucleotide included an approximately 1.3- to 1.4-fold increase in spleen weight (relative to control) at 70 mg/kg/week (Fig. 3A).
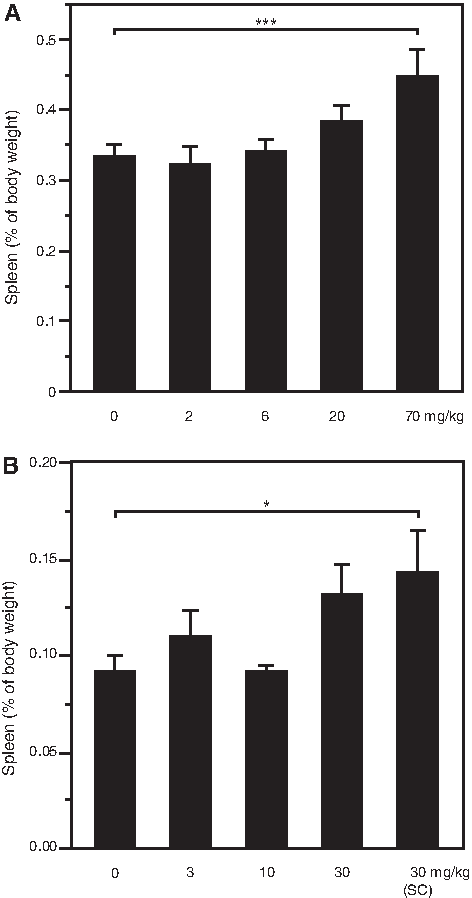
Spleen weight adjusted to body weight on day 44/45.
Clinical signs, organ weights, and gross observations: Monkey
Treatment with ISIS 481464 for 6 weeks at dose levels of up to 30 mg/kg/week was well tolerated and no treatment-related mortality was observed in cynomolgus monkeys. There were no treatment-related changes in clinical signs, food consumption, body weight, ocular abnormalities, electrocardiograms, heart rate, blood pressure, serum chemistry, hematology, or urinalysis parameters during the treatment or recovery periods.
Treatment-related gross findings included enlarged spleen, which was limited to only 2 males at 30 mg/kg/week (single animals, each treated by the IV or SC routes). Absolute and body weight relative spleen weights for these animals were increased (approximately 2- to 2.5-fold relative to control) and contributed to slight increase in the group spleen weight at 30 mg/kg/week (Fig. 3 B). Thymus atrophy was also observed in 2 females at 30 mg/kg/week (a single animal in either IV or SC), however, this finding was considered secondary to stress and not directly related ISIS 481464 treatment.
Clinical chemistry and hematology: Mouse
Changes in hematology parameters were limited to the 70 mg/kg/week dose level and included an approximately 2.4-fold increase (relative to control) in relative monocyte counts in females only (Fig. 4A).
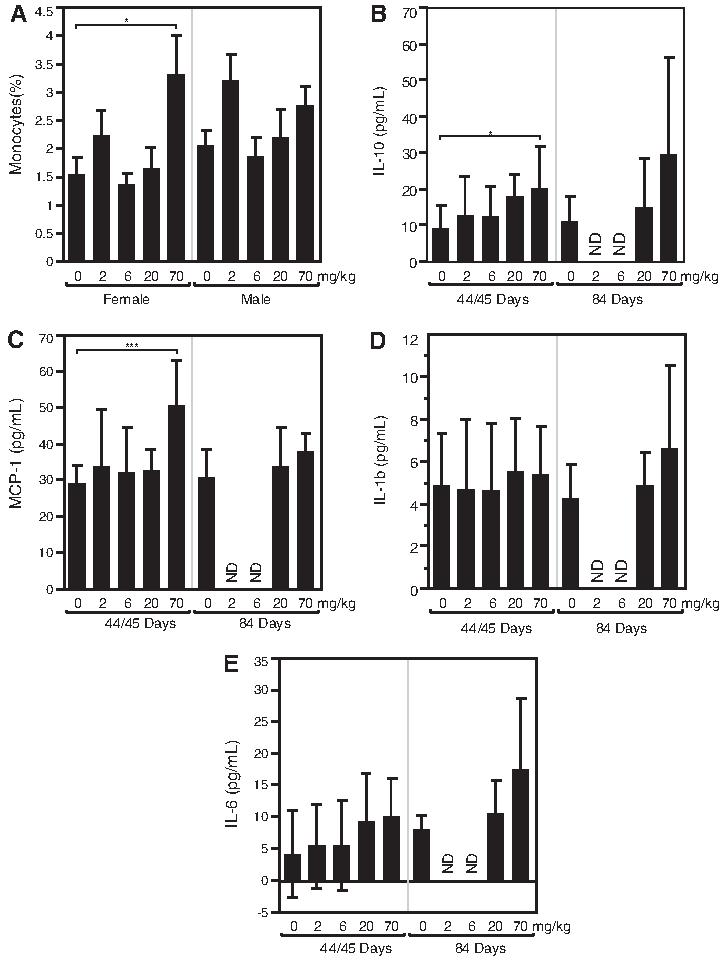
Other changes included increased circulating IL-10 (3.7-fold relative to control) and MCP-1 (1.9-fold relative to control) in plasma samples collected 48 hours following the last dose (Fig. 4 B, C) while IL-1β and IL-6 were unchanged (Fig. 4 D, E).
Minimal increases in liver enzymes were observed at 70 mg/kg/week, including a 1.4-fold increase (relative to control) in alanine aminotransferase (ALT) and a 1.7-fold increase (relative to control) in aspartate aminotransferase (AST) (data not shown). No change in bilirubin was observed.
Clinical chemistry and hematology: Monkey
There were no changes in serum chemistry or hematology parameters, but there were transient prolongation of intrinsic clotting times and transient alternative pathway complement activation. Transient and minimal increases in activated partial thromboplastin time (aPTT) clotting times were observed following IV administration of ISIS 481464 (Fig. 5). The change was similar on days 1 and 42 at the 30 mg/kg/week dose level (IV only). Peak elevations occurred at the end of 1 hour IV infusion (approximately 1.5-fold relative to predose levels) or 4 hours post dose for SC (approximately 1.1-fold relative to predose) administration, and returned to baseline by 24 hours post dose. The greater increase in aPTT after IV infusion compared to SC dosing is consistent with higher plasma observed mean plasma concentration (Cmax) (276 vs. 96 μg/mL respectively) and no effects on aPTT were observed in monkeys at the lower dose levels. No other changes in clotting time parameters (fibrinogen or PT) were observed in both sexes at dose levels of 30 mg/kg/week. There were no sequelae associated with aPTT prolongation. Thus, the increase in clotting time was not considered clinically significant based on the rapid reversibility, absence of hemorrhage, the selective effect on the intrinsic pathway, and the absence of changes in PT or in fibrinogen levels. This transient effect on aPTT is a known class effect of ASOs that is dependent on plasma ASO concentrations (Henry et al., 1997c; Farman and Kornbrust, 2003).
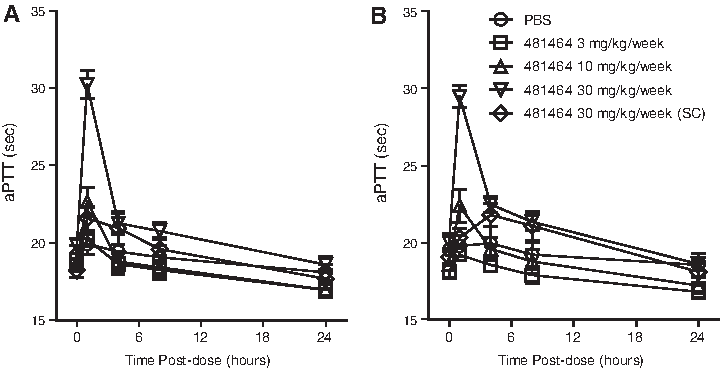
Effect of ISIS 481464 on clotting times in cynomolgus monkeys. The values presented are the mean and standard error of activated partial thromboplastin time (aPTT) measured relative to initiation of treatment on
The transient production of complement activation split product (Bb) was an anticipated effect with oligonucleotides in the monkey. Complement split product (Bb) concentration increased approximately 5.6- to 8-fold at 1 to 4 hours on days 1 and 42, respectively, at the 30 mg/kg/week dose level (IV) and returned to baseline levels by 24 hours (Fig. 6 A, B). The increases in Bb were similar at both time points and thus not cumulative over the course of treatment. Such increases in complement split product are indicative of alternative complement pathway activation, and only observed at the highest IV doses evaluated and associated with the highest plasma concentrations of ISIS 481464. Minimal decreases in C3 levels (approximately 10% to 15% reductions relative to baseline) were also observed at ≥10 mg/kg/week 48 hours after last dose on day 42 (Fig. 6C). The decrease in C3 observed did not fully correlate with the elevations in Bb observed at 30 mg/kg/week (IV only), suggesting an alternative etiology for the C3 decreases observed. The pattern of C3 reduction was better correlated with the reduction in STAT3 protein (Fig. 2B, C). Activation of the alternative complement pathway has been previously observed with antisense oligodeoxynucleotides and shown to be a threshold drug plasma concentration phenomenon in monkeys (Henry et al., 1997b; Henry et al., 2002).
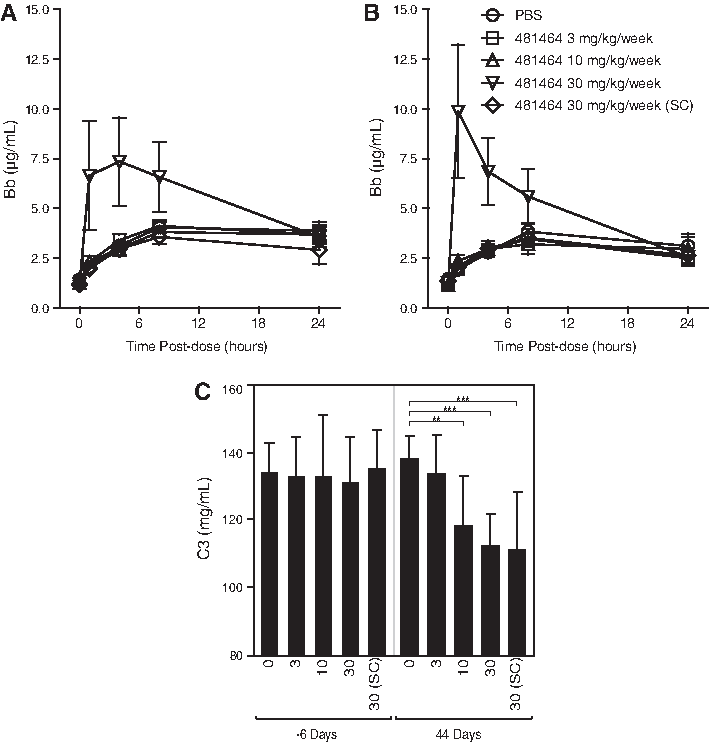
Effect of ISIS 481464 on complement split products Bb in cynomolgus monkeys on day 1
Histopathology
Mouse
Microscopic findings mainly consisted of the presence of minimal to moderate basophilic granules in kidney, liver, and histiocytes in multiple tissues that increased in severity and/or incidence in a dose-dependent manner (Table 3). The accumulation of basophilic granules was dose dependent and accompanied by minimal increase in hypertrophy/hyperplasia of Kupffer cells at doses ≥20 mg/kg/week. Oligonucleotides like ISIS 481464 stain with hematoxylin, and therefore, their uptake into cells can be visualized microscopically as basophilic granules (Butler and Murray, 1997). Thus, basophilic granules in tissue is indicative of the presence of oligonucleotide in organs where compound exposure is the greatest (liver and kidney) and is not considered a toxicologically significant observation (Henry et al., 2000b).
Values are mean±standard deviation.
Intact ISIS 481464: mouse tissue concentrations were measured at 48 hours after the last injection (n=6).
Recovery groups were sacrificed 42 or 43 days following final dosing.
LLOQ, lower limit of quantitation; NOAEL, no observed adverse effect level.
Lymphohistiocytic cell infiltration in the liver related to pro-inflammatory effect of the oligonucleotide was also observed in both sexes at ≥20 mg/kg/week. The remaining changes observed following 6 weeks of treatment with ISIS 481464 were related to the cellular uptake of oligonucleotide into histiocytes in multiple organs. The hypertrophied and vacuolated histiocytes observed in multiple organs [kidneys, urinary bladder, liver (within Kupffer cells), heart, spleen, salivary glands (submandibular and parotid), pancreas, skeletal muscle, tongue, lung, lacrimal glands, esophagus, stomach, duodenum, jejunum, ileum, cecum, colon, rectum, skin, mammary gland, femur, sternum, injection site, lymph node (inguinal, mandibular, mesenteric, renal, and iliac), testes, epididymides, prostate, seminal vesicle, ovary/oviduct, uterus/cervix, and vagina] with dose-dependent increase in incidence and severity at dose levels ≥20 mg/kg/week. With the exception of the liver, no cell infiltrates were observed in other tissues. This observation might be a reflection of the short duration of treatment, as a mild increase in infiltrate would be expected with increased duration of treatment as suggested by mild elevation in chemokines observed at 6 weeks. Chemokine increases generally accompanied multiorgan cell infiltrates in longer-term treatment.
Following a 6 week treatment-free period, the findings identified at 6 weeks of ISIS 481464 treatment were completely (20 mg/kg/week) or partially (70 mg/kg/week) resolved. At the 70 mg/kg/week dose group, the presence of basophilic granules in major tissues (Kupffer cells of the liver, tubular epithelium of the kidney) and Kupffer cell hypertrophy/hyperplasia were noted with reduced incidence and severity after the 6-week recovery period. The continued but reduced presence of basophilic granules is indicative of continued but decreased presence of drug in those tissues.
Monkey
Treatment-related microscopic findings were dose dependent and observed in the kidneys, liver, lymph nodes, urinary bladder, and subcutaneous injection sites following 6 weeks of treatment. Despite an absolute and relative spleen weight increase in some animals (Fig. 4), there was no microscopic correlation to splenomegaly. Basophilic granules reflecting the cellular uptake of ISIS 481464 were mainly observed in the high dose groups in the tubular epithelium of the kidney and in Kupffer cells of the liver.
In the kidney, minimal to moderate basophilic granule accumulations in the cytoplasm of tubular epithelial cells in the outer cortex were observed at 30 mg/kg/week (Table 4). Minimal to slight periglomerular fibrosis accompanied with slight tubular degeneration/regeneration was observed in two of 12 animals (a single male and female) at 30 mg/kg/week (SC only). Minimal tubular degeneration/regeneration was also observed in control (a single male and female) suggesting the degeneration/regeneration does not appear to be solely related to ISIS 481464 treatment (Table 5). The association of the periglomerular fibrosis with treatment was not clear based on the low incidence and the absence of this finding in a prior independent pilot study (data not shown).
Values are mean±standard deviation.
No changes in renal or hepatic function at doses up to 30 mg/kg/week.
Intact ISIS 481464: monkey tissue concentrations at 48 hours after the last infusion or injection (n=4 or 6).
Kidney=kidney cortex for monkey.
n=2, values reported as median (minimum, maximum).
Combined data from toxicology and toxicokinetic dose groups.
Recovery group were sacrificed 54 days following final dosing.
Kidney=kidney cortex for monkey.
Presented as mean with standard deviation.
Minimal tubular degeneration/regeneration was also present in 2 of 6 control monkeys.
- No findings.
In the liver, minimal to slight accumulation of basophilic granules within Kupffer cells was observed at 30 mg/kg/week and limited to 1 male monkey at 10 mg/kg/week. No functional changes were noted in the liver.
In the mesenteric and inguinal lymph nodes, minimal to moderate basophilic granule accumulation in the cytoplasm of histiocytes were observed at doses ≥3 mg/kg/week in a dose-dependent manner. Basophilic granules were also observed in histiocytes in urinary bladder and ovaries at 30 mg/kg/week (minimal to slight).
In the injection site skin, minimal to slight inflammatory cell foci were observed only in males at the 30 mg/kg/week dose level (SC).
Findings at the recovery necropsy were similar to those observed at the terminal sacrifice but with decreased incidence and/or severity, consistent with partial to full recovery following the cessation of treatment.
Toxicokinetics
Mouse
The highest concentrations of ISIS 481464 were observed in kidney with tissue concentrations ranging from 37 to 378 μg/g following 6 weeks of treatment (Table 3). As such, basophilic granule staining was observed in the proximal tubular epithelium (minimal) at 70 mg/kg/week. The second highest concentrations of ISIS 481464 were observed in liver with tissue concentrations ranging from 38.1 to 317 μg/g following 6 weeks of treatment at doses of 6 to 70 mg/kg/week, respectively. Data from the toxicokinetic portion of the study demonstrated dose-dependent exposure of mice to ISIS 481464 following IV administration. Maximum Cmax and area under the curve (AUC) values were dose dependent and increased nearly, or slightly less, than dose proportionally over the dose range evaluated (Table 6). Plasma disposition of ISIS 481464 was multiphasic and relatively rapid, mainly representing distribution to extracellular fluid and tissues. As an index measure to reflect the initial rapid and almost complete distribution of ISIS 481464 from plasma to tissues within the first 48 hours post dose, mean residence time (MRTlast) was calculated after the first (day 1) and ninth (day 42) doses. Following IV bolus administration, mean MRTlast values ranged from 2.5 to 4.5 hours in mice for both evaluated dose levels. Rapid clearance from plasma correlated with extensive uptake into kidney and liver. Tissue concentrations of ISIS 481464 were dose dependent and concentrations generally increased less than dose proportionally in kidney, while liver concentrations increased nearly dose proportionally. Kidney concentrations ranged from approximately 37 to 317 μg/g at doses of 2 to 70 mg/kg/week, respectively. Liver concentrations (intact ISIS 481464) ranged from approximately 38.1 to 317 μg/g at doses of 6 to 70 mg/kg/week, respectively (Table 3).
n=3 mice per time point were used to achieve a single pooled plasma profile for PK parameter determination.
IV loading doses administered on days 1, 3, 5, and 7, followed by weekly injections starting on day 14 and continuing until day 42.
AUC0–48 was calculated as AUC from 0 to 48 hours post dose.
(MRTlast) calculated from 0 to 48 hours after dosing.
CLp(0–48hr)=(dose*1000*body weight)/AUC0–48; where body weight=0.033 kg for day 1 and 0.038 kg for day 42).
AUC, area under the plasma concentration time curve; C0, extrapolated plasma concentration at time=0; CLp, plasma clearance; Cmax, observed maximum plasma concentration; MRTlast, mean residence time.
Monkey
Data from the toxicokinetic evaluation demonstrated dose-dependent exposure of ISIS 481464 in plasma, urine, and systemic tissues following intravenous or subcutaneous administration. Plasma toxicokinetics demonstrated dose-dependent and nearly dose-proportional peak (Cmax) and total (AUC) exposure of monkeys to ISIS 481464 following 1-hor IV infusion doses ranging from 3 to 30 mg/kg/week. The median (range) time of maximum observed plasma concentration (tmax) after SC administration at 30 mg/kg/week was observed at 3 (2 to 3) hours after the injection. Observed mean plasma concentration (Cmax) was approximately 3-fold lower with SC dosing compared with 1-hour IV infusion dosing, while AUC were similar at 30 mg/kg/week following both routes of administration (Table 7), indicating nearly complete bioavailability after SC dosing. Plasma concentrations rapidly declined following IV or SC administration. At all dose levels and routes of administration, ISIS 481464 distribution appeared to be multiphasic and indicative of rapid distribution to tissues. As an index measure to reflect the initial rapid and almost complete distribution of ISIS 481464 from plasma to tissues within the first 48 hours post dose,MRTlast was calculated after the first (day 1), fourth (day 7), and ninth (day 42) doses. Following 1-hour IV infusion administration, mean MRTlast values ranged from 1.8 to 3.9 hours in monkeys across all doses (Table 7). The estimated terminal elimination half-life (t1/2λz) values in the 10 mg/kg (IV) and 30 mg/kg (IV or SC) groups ranged from 20.4 to 31.8 days. The highest tissue concentrations of ISIS 481464 were found in kidney cortex followed by liver (Table 4). ISIS 481464 cleared slowly from kidney and liver with an estimated elimination half-life of 13.8 to 30.3 days in kidneys and 14.2 to 25.2 days in liver (Table 8). The exposure estimates demonstrate a positive correlation between plasma trough and tissue levels and target organ concentration. These results are consistent with rapid and extensive distribution of ISIS 481464 from plasma to tissues, and subsequent slow tissue clearance of the parent compound as mainly chain-shortened metabolites and also intact drug.
MRTlast calculated from 0 to 48 hours after dosing only.
CLp(0–48hr)=[dose (mg/kg)*1000]/AUC0–48; note: CLp(0–48hr)/F calculated for SC dosing.
tissue disposition half-life (t1/2λz) calculated for group 6, day 7 using the sparse sampling function in Phoenix™ WinNonlin®.
SC.
Median range.
nr, not reported; Tmax, time at which the maximum concentration is observed.
t1/2λz determined using only end of dosing and recovery tissue concentrations (2-point curve) for groups 4 and 5; see text for details,
Discussion
This is the first report of the toxicity profile of a new chemical subclass class of antisense oligonucleotides called cEt gapmers (Seth et al., 2008; Seth et al., 2009). The main differences in the structures were a slightly shorter length (16 nucleotides compared with the more common 19 to 21 nucleotides) and the presence of bicyclic nucleic acid (BNA) modification. This modification is similar to locked nucleic acid, but distinct to the 2′-alkyl modifications (Seth et al., 2008; Seth et al., 2009). These BNA modifications afford greater hybridization affinity to RNA, which allows them to be shorter in length. The most noteworthy findings observed for this single-stranded phosphorothioate cEt-modified 16-nucleotide antisense compound were, in general, nonspecific class effects that are typical of those for the related 2′-MOE-modified antisense oligonucleotide class (Henry et al., 2007). The pharmacokinetic properties were also very similar to 2′-MOE-modified antisense oligonucleotide (Geary et al., 2007). No toxicologically relevant findings were considered related to the pharmacologic inhibition of STAT3 were observed in these normal animals.
Effects of STAT3 inhibition and reduction
The possibility that pronounced or prolonged inhibition of STAT3 expression might have deleterious effects on normal animals was addressed by evaluating the toxicity of ISIS 481464 in monkeys. Levels of STAT3 protein were reduced by approximately 90% at 10 mg/kg and above, demonstrating that the doses were sufficient to reduce STAT3 expression in the absence of toxicity. While there was no apparent toxicity associated with 90% decrease in STAT3, decreased levels of STAT3 may weaken the immune system and increase sensitivity to pathogens (Bode et al., 2012). It must be noted that these were normal animals, and therefore the potential for increased risk of infections should be carefully monitored in the clinic. Interestingly while liver STAT3 protein levels were decreased by more than 90% of control in monkey, STAT3 mRNA was decreased only by 50% of control. This discrepancy between mRNA and protein level reduction after ASO treatment has been observed before and is not unique to ISIS 481464. The difference in reduction between protein and mRNA level might be explained by multiple terminating mechanisms by which ASO binding to RNA can potentially regulate mRNA level including RNaseH-mediated degradation, but also through nondegradative mechanisms such as translation arrest, inhibition of splicing, and others (Chiang et al., 1991; Kole et al., 2012).
Transient aPTT prolongation and complement activation
Increases in aPTT and serum Bb concentrations were observed at the 30 mg/kg dose of ISIS 481464 typical of other phosphorothioate oligonucleotide, but seemed to be of lesser magnitude. This was particularly evident in the case of complement where activation was only observed after the 1-hour IV infusion, and not following the 30 mg/kg SC injection, which would ordinarily be expected to activate complement. This may reflect the shorter oligonucleotide length and resulting decrease in nonspecific protein interactions.
One unexplained change in complement parameters was in the absence of complement split product (Bb) elevations in 2 dose groups where reductions in C3 were observed (10 mg/kg/week IV and 30 mg/kg/week SC). C3 has been shown to decrease as a consequence of repeated activation by antisense oligonucleotides. One possible explanation of the data is that the observed C3 reduction at doses ≥10 mg/kg/week may be related to STAT3 reduction rather than a result of complement activation. STAT3 inhibition would be expected to decrease inflammatory signals and it is possible that a decrease in C3 could be part of reducing acute phase processes.
Renal effects
The renal effects observed for ISIS 481464 in monkeys are very similar in nature to those observed with other related 2′MOE modified oligonucleotides (Henry et al., 2012). Kidney is the organ that contains the highest concentrations of oligonucleotides, and all phosphorothioate oligonucleotides are almost exclusively taken up into proximal tubular epithelium at the brush border and processed into endosomes and eventually lysosomes. The most consistent microscopic observation in both species treated with ISIS 481464 was the presence of basophilic granules in the renal proximal tubular epithelial cells. Severity of basophilic granules was minimal to moderate and only observed at the highest dose and limited in incidence. They reflect the oligonucleotide uptake by these cells and compartmentalization into phagolysosomes and alone are not considered toxicologically significant (Monteith et al., 1999; Henry et al., 2000b; Henry et al., 2007). Oligonucleotide concentrations in the renal cortex of monkey at the 30 mg/kg/week dose were up to 2075 μg/g, which is somewhat lower than often reported for 20-nucleotide MOE ASO. While ISIS 481464 has a similar tissue distribution properties than 2′MOE modified oligonucleotide, ISIS 481464 appears to have less effect on tubular epithelial cells at similar concentrations because no frank tubular degeneration was apparent. This possibly reflects the shorter oligonucleotide length and lower concentration.
Isolated instances of minimal to slight periglomerular fibrosis epithelium were observed following 6 weeks of treatment and were not observed in an independent study. No functional changes in the kidneys, as measured by evaluation of standard serum chemistry and urinalysis parameters, were associated with periglomerular fibrosis, indicating that proximal tubules were functioning properly. Therefore, the clinical significance or toxicological relevance of the periglomerular fibrosis is uncertain. The kidney changes were also reversible with the clearance of ISIS 481464 and its metabolites following the treatment-free period.
Hepatic effects
Liver is also a target organ for oligonucleotide distribution. In mouse liver, dose-dependent hyperplasia/hypertrophy (minimal to moderate) in Kupffer cells with basophilic granules were observed at doses ≥20 mg/kg/week. In addition, minimal to moderate lymphohistiocytic cell infiltrates as well as minimal changes in serum transaminases, ALT, and AST were observed at 70 mg/kg/week. The elevations in liver enzymes returned to predose levels 6 weeks following cessation of treatment. The changes were not associated with any microscopic evidence of hepatocellular injury or changes in tissue weight at 6 weeks. These changes were considered secondary to the pro-inflammatory effects; as mild elevations of ALT and AST are known to be exacerbated by the pro-inflammatory effects of oligonucleotides in rodents (Henry et al., 1997d; Monteith et al., 1998). These changes are not related to the inhibition of STAT3, since the ISIS 481464 is not pharmacologically active in mice. These findings were partially or fully reversible following the cessation of treatment, consistent with clearance of oligonucleotide from tissue.
In monkey, changes in the liver were limited to the presence of minimal to slight basophilic granules within Kupffer cells at doses ≥10 mg/kg/week. Macrophages are known to phagocytize oligonucleotides and cellular uptake can be visualized histologically as basophilic granules within Kupffer cells. There were no changes in liver enzymes or hepatic function (ALT, or bilirubin) observed in monkeys following 6 weeks of treatment despite the fact that liver concentrations of ISIS 481464 were higher than those observed in mice (587 μg/g at 30 mg/kg/week). The absence of ALT changes in monkeys reflects the general observation that primates are less sensitive to the proinflammatory effects of oligonucleotide than rodents (Henry et al., 2000b; Henry et al., 2001).
Immune system
Several findings in mice, including increased spleen weight, lymphohistiocytic cell infiltrates in the liver (minimal to moderate in severity), and small increases in inflammatory markers (IL-10 and MCP-1), that were observed at ≥20 mg/kg/week were collectively regarded as proinflammatory changes. There were no significant alterations in other inflammatory cytokines (IL-1β, IL-6).
Other than the presence of inflammatory cell infiltrates at the SC injection site in the 6-week study, proinflammatory effects were not observed in monkeys.
Proinflammatory effect and cytokine production, particularly in rodents, is a common effect associated with this class of compound (Zhao et al., 1997; Monteith et al., 1998). Although this is an expected observation at high doses, the degree of inflammatory change can vary widely, depending on either of the sequence or structural modifications of any particular oligonucleotides. By comparison, ISIS 481464 treatment resulted in a relatively low degree of proinflammatory effect in both species. This is attributed in large part to the shorter length of the oligonucleotides (16 nucleotides compared with the more common range of 19 to 21 nucleotides for most MOE ASO (Henry et al., 2007). Sequence also certainly plays a role in that this ASO is devoid of CpG dimmers or other sequence motifs known to be associated with inflammatory effects.
Dose–response relationships characterized in these studies and our understanding of the toxicity and pharmacokinetics of related 2′MOE ASOs indicate that there is a sufficient therapeutic margin to assure the safe clinical use of ISIS 481464 at the expected dose and regimen for the treatment of cancer. Compared with existing cytotoxic agents for cancer chemotherapy, the use of antisense oligodeoxynucleotides is a less toxic approach for treatment.
Overall, the toxicity profile of the cEt-modified antisense oligonucleotide, ISIS 481464, in mice and monkeys is very similar to 2′-MOE-modified oligonucleotide (Henry et al., 2007). Most of the class effects are related to protein binding conferred by the presence of phosphorothioate linkage (LEVIN, 1999). For Isis 481464, those class effects tended to be slightly lower that has typically been reported, presumably because of the shorter length and lower phosphorothioate content. Furthermore, class effect related to tissue exposure had similar target organ concentration relationship compared with 2′-MOE modified oligonucleotides. More importantly, ISIS 481464 treatment did not reveal new toxicity associated with cEt ASO. Additional experience with the characterization of other cEt-modified oligonucleotides will be needed before definitive conclusions can be drawn.
Footnotes
Acknowledgments
S.A. Burel, S.-R Han, H.-S. Lee, T.-W. Kim, D.A. Norris, S.-Y. Park, A.R. MacLeod, B.P. Monia, and S.P. Henry participated in research design. S.-R Han, H.-S. Lee, T. Machemer, T. Zhou, G. He, and Y. Kim conducted experiments. S.A. Burel, S.-R Han, H.-S. Lee, B.-S. Lee, T. Machemer, and D.A. Norris performed data analysis. S.A. Burel, T.K. Kim, S. Lio, D.A. Norris, and S.P. Henry wrote or contributed to the writing of the manuscript.
Author Disclosure Statement
No competing financial interests exist.