
Abstract
Background:
Ischemia-reperfusion (I/R) is the main cause of acute kidney injury (AKI) in patients. We investigated renal microRNA (miRNA) expression profiles and the time course of changes in selected miRNA expressions after renal I/R to characterize the miRNA network activated during development and recovery from AKI.
Methods and Results:
One day after lethal (30 minutes) and sublethal (20 minutes) renal ischemia, AKI was verified by renal histology (tubular necrosis, regeneration), blood urea nitrogen (BUN) level, renal mRNA expression, and plasma concentration of neutrophil gelatinase-associated lipocalin (NGAL) in C57BL/6J mice. On the first day after 30-minute, lethal I/R miR-21, miR-17-5p, and miR-106a were elevated out of the 21 miRNAs successfully profiled on the Luminex multiplex assay. After 20-minute, sublethal I/R, renal miR-17-5p and miR-106a expressions were elevated on the first and second days of reperfusion, while miR-21 expression increased later and lasted longer. Renal miR-17-5p and miR-21 expressions correlated with each other. Renal function returned to normal on the fourth day after sublethal I/R.
Conclusions:
Our results demonstrate that besides miR-21, miR-17-5p, and miR-106a are additionally activated during the maintenance and recovery phases of renal I/R injury. Furthermore, a correlation between renal miR-17-5p and miR-21 expressions warrants further investigation of how they may influence each other and the outcome of renal ischemia-reperfusion injury.
Introduction
MicroRNAs (miRNAs) are small, ∼22-nucleotide-long, double-stranded RNAs that are involved in posttranscriptional gene-expression regulation through multiple mechanisms (Eulalio et al., 2008). The role of miRNAs in the I/R-induced AKI was first reported in a proximal tubular cell targeted Dicer knockout mouse model, in which miRNA depletion attenuated renal ischemic damage (Wei et al., 2010). Further research pointed to a prominent role of miR-21 in the prevention of tubular cell death after hypoxia in vitro (Godwin, Ge et al., 2010). MiR-21 contributed to the beneficial effects of the delayed renal ischemic- preconditioning (Xu et al., 2012; Jia et al., 2013). Furthermore, miR-21 was proposed not only as a possible urinary biomarker of AKI (Saikumar et al., 2012), but as a prognostic marker after cardiac surgery-induced AKI as well (Du et al., 2013). Godwin et al., reported the course of changes in the expression of miRNAs differentially expressed after I/R injury (Godwin et al., 2010); however, only the histological damage and immune infiltration were assessed without measuring kidney function in this study. Changes in miRNA expression did not overlap in the post-ischemic kidney in the 2 microarray studies performed so far (Godwin et al., 2010; Wei et al., 2010). Therefore, additional experiments are required to identify miRNAs, which are altered the most by I/R injury.
In order to further identify miRNAs regulated during reperfusion injury, we investigated the miRNA expression pattern in I/R injury-induced AKI with multiplex analysis using a novel microsphere bead and flow cytometry based system (Luminex). Furthermore, to gain more information about the possible role of the most affected renal miRNAs we have carried out a time-course analysis of miRNA expression after lethal and sublethal renal ischemia. Our results demonstrated that besides miR-21, miR-17-5p, and miR-106a expressions were also elevated during the maintenance and recovery phases of the I/R-induced AKI. Furthermore, miR-21 and miR-17-5p correlations were influenced by the stages of reperfusion—an interesting finding which should be investigated further.
Methods
Animals
Male C57BL/6J mice (Charles River) weighing 29.4±2.9 g were maintained under specific pathogen-free conditions with access to standard rodent chow (Altromin standard diet) and tap water ad libitum. Approval from the Semmelweis University Animal Research Ethical Committee has been obtained under the registration No. XIV-I-001/2103-4/2012.
Kidney ischemia-reperfusion injury
Renal ischemia-reperfusion (I/R) injury was performed as described previously (Hamar et al., 2004). Briefly, the experiments were carried out at room temperature, 24±0.5°C, using standard operating procedures. The intra-abdominal temperature was maintained using a heating pad (HK-3, DOPS). Under light ether anesthesia, the animals were narcotized with an intraperitoneal (i.p.) injection of 80 mg/kg ketamine and 4 mg/kg xylazin cocktail (CP-Pharma Handelsgesellschaft mbH). The left renal pedicle was prepared and clamped to obtain sublethal (20-minute) and lethal (30-minute) ischemia as determined in previous studies (Heemann et al., 2000; Hamar et al., 2004). The right kidney was removed. Sham-operated control mice were also prepared in the same manner as those subjected to I/R except that the renal pedicle was not clamped.
Sacrifice and blood and organ collection
At the specified time-points animals were sacrificed under ether anesthesia. Right before sacrifice, 500 U per mouse of heparin (Merckle GmbH.) was injected i.p., and blood was collected from the thoracic cavity after cross-sectioning the vena cava. After blood urea measurements blood was centrifuged at 1,500g for 8 minutes at 4°C to obtain plasma for analysis. The kidney was removed after a slow transcardiac perfusion with 20 mL physiological salt solution precooled to 4°C. Plasma and renal tissue samples were snap frozen in liquid nitrogen and kept at −80°C until use.
Plasma urea and NGAL ELISA
Renal function was evaluated by determination of blood urea nitrogen (BUN). Blood urea levels were measured from 32 μL whole blood obtained during organ collection with Reflotron® Urea test strips (Roche Diagnostics GmbH) on a Reflotron® Plus device (Roche Diagnostics GmbH) as described in the manufacturer's protocol. Urea values were divided by 2.14 to obtain BUN levels.
Neutrophil gelatinase associated lipocalin (NGAL, Lipocalin-2) has been demonstrated to be a sensitive marker of tubular epithelial damage (Devarajan et al., 2003). The plasma NGAL level was determined with a mouse Lipocalin-2/NGAL DuoSet ELISA Development kit (R&D Systems) as described by the manufacturer. Shortly, the 96-well plates (Nunc™ GmbH and Co. KG) were coated with the capture antibody, and the nonspecific binding sites were blocked with reagent diluent [1% bovine serum albumin (BSA) in phosphate-buffered saline, pH 7.2–7.4]. Adequately diluted samples (103- to 105-fold) were incubated on the plate in duplicates for 2 hours, and then the detection antibody was added. Next, streptavidin–horseradish peroxidase was linked to the biotinylated detection antibody, followed by a short incubation with TMB substrate (Sigma-Aldrich Chemie GmbH). A washing session (5 times with 300 μL of washing buffer) was performed after each step until the addition of the substrate solution. The enzymatic reaction was terminated by stop solution containing H2SO4. The optical density was measured with Victor3™ 1420 Multilabel Counter (PerkinElmer, WALLAC Oy) at 450 nm with wavelength correction set to 544 nm. The NGAL concentrations were calculated with WorkOut software (Dazdaq Ltd.), using a four-parameter logistic curve-fit.
Histology of renal tissue injury and NGAL immunohistochemistry in tissue microarray slides
Renal tissue samples fixed in 4% buffered formaldehyde were dehydrated and embedded in paraffin wax (FFPE) for histology and immunohistochemistry. Experiments were evaluated morphologically by tissue microarray (TMA) as described previously (Krenacs et al., 2010). Briefly, blocks of 70-sample TMAs contained duplicates of 2-mm diameter cylinders cut by the computer-controlled puncher of the TMA Master Device (3DHISTECH Kft) from each FFPE kidney. For morphology and immunohistochemistry, 4-μm-thick sections were cut from the TMA blocks. Renal tubular necrosis and regeneration were evaluated in Periodic acid-Schiff stained TMA sections. A histological score of 0 to 4 was given by a pathologist blinded to the origin of the tissue as follows: 0, no lesion; 1, minimal or focal changes affecting less than 20% of the field; 2, mild changes or the extension of the lesion/regeneration to approximately 25% of the field; 3, moderate changes or the extension of the lesion/regeneration to less than 50% of the field; 4, severe changes or the extension of the lesion/regeneration to more than 50% of the field.
Renal tubular cell damage was evaluated by NGAL immunostaining. Dewaxed and rehydrated TMA sections were cooked at 100°C for 25 minutes in a 0.01M Tris-HCl and 0.1 EDTA buffer (TBS; pH 9.0) for antigen retrieval. The immunostaining involved consecutive incubations of TMA sections in 1% BSA in TBS (pH 7.4) for 15 minutes, rabbit anti-human NGAL immunoglobin-G (IgG) (1:100; R&D Systems) for 16 hours and in goat anti-rabbit IgG EnVision-peroxidase polymer kit (Dako) for 40 minutes, all at room temperature. Tissue-bond peroxidase activity was developed with a DAB/H2O2 chromogen/substrate kit (Dako). Immunostained TMA slides were digitalized using a Pannoramic Scan instrument, and the results were analyzed with the Pannoramic Viewer software (3DHISTECH).
RNA preparation
Total RNA was extracted from the upper third of the kidney with TRIzol® Reagent (Invitrogen™) according to the protocol provided by the manufacturer (CHOMCZYNSKI, 1993). In brief, the frozen renal tissues were homogenized by an IKA® DI 18 basic grinder (IKA® Works do Brasil Ltd. Chloroform (Sigma-Aldrich, Inc.) was added to each sample and mixed by vortex. The aqueous phase was separated from the organic phase by centrifugation. RNA was precipitated from the transferred aqueous phase with an equal quantity of isopropyl alcohol by incubation for 30 minutes at room temperature. The RNA pellet was washed twice with 75% ethyl alcohol and dissolved in 100 μL RNase-free water. The RNA pellet was treated with DNase 1, RNase-free (Fermentas) to eliminate possible DNA contamination. The DNase was inactivated by phenol/chloroform extraction (Fluka, Sigma-Aldrich). The RNA concentration and purity was inspected with NanoDrop 2000c spectrophotometer (Thermo Fisher). All RNA samples had an absorbance ratio (260 nm/280 nm) above 1.8. To investigate RNA integrity, samples were electrophoresed on 1% agarose gel with BioRad iCycler™ Thermal Cycler (Bio-Rad Laboratories, Inc.), and the 28S and 18S ribosomal RNA fraction integrity was examined. The RNA solutions were kept at −80°C until further procedures.
Multiplex analysis of the microRNA profile
The miRNA expression pattern of the kidney samples was analyzed after 24 hours of reperfusion following 30 minutes I/R (n=9) or sham operation (n=8). Based on a review of available literature about miRNA networks in renal diseases (Kaucsar et al., 2010), the largest, most suitable, commercially available Luminex miRNA panel was selected. The expression of 46 microRNAs (Table 1) was determined with the Vantage™ microRNA Detection Kit (Marligen Biosciences, Inc.) on the Luminex® 200™ System (Luminex Corporation) as described in the protocol. First, 1.5 μg of the extracted RNA was poly(A)-tailed and biotinylated, and then sample RNA hybridization with the bead mix and microRNA detection was performed. All samples were tested in duplicates and the background median fluorescence intensity (MFI) was subtracted before further calculations. MicroRNAs with low MFI (<100, ≈3 times the background MFI) were excluded from the statistical analysis.
Quantitative real-time polymerase chain reaction analysis of miRNAs and NGAL expression in renal tissue
MicroRNA expressions were evaluated with TaqMan probes (Chen et al., 2011). First, complementary DNA (cDNA) was reverse-transcribed (RT) from 5-ng RNA sample using a miRNA-specific, stem-loop RT primer (for miR-21, miR-17-5p, miR-106a, and U6 snRNA) from the TaqMan® Small RNA Assays and reagents from the TaqMan® MicroRNA Reverse Transcription Kit (Applied Biosystems™), as described in the manufacturer's protocol. Second, in the real-time polymerase chain reaction (PCR) step, PCR products were amplified from the cDNA samples using the TaqMan® Small RNA Assay together with the TaqMan® Universal PCR Master Mix 2. All measurements were done in duplicates, and the miRNA expressions were normalized to the U6 small nuclear RNA (snRNA) applied as an endogenous reference (Liu et al., 2011; Nie et al., 2011). The miRNA expressions were calculated with the relative quantification (ΔΔCq) method, and the efficiency of the quantitative PCR reaction was verified with standard curves.
NGAL mRNA levels were measured by double-stranded DNA dye–based real-time PCR. Reverse transcription into cDNA was carried out by the High-Capacity cDNA Archive Kit (Applied Biosystems™) according to the manufacturer's protocol. In brief, 1 μg of total renal RNA was denaturated at 70°C for 5 minutes. After the annealing of the random hexamer primers on the RNA template at 25°C for 10 minutes, cDNA was synthesized at 37°C for 2 hours. The reaction was terminated by heat inactivation (85°C for 2 minutes). The NGAL (Lcn2) gene expression from kidney tissue homogenates was evaluated on the Bio-Rad C1000™ Thermal Cycler with CFX96™ Optics Module real-time PCR system (Bio-Rad Laboratories, Inc.). The PCR reaction was performed with Maxima™ SYBR Green qPCR Master Mix (Fermentas), according to the manufacturer's protocol. Primers for NGAL were designed by NCBI/Primer-BLAST online software (forward: ACg gAC TAC AAC CAg TTC gC; reverse: AAT gCA TTg gTC ggT ggg g) and synthesized by Integrated DNA Technologies. The endogenous reference gene was GAPDH (forward primer: CCA gAA TgA ggA TCC CAg AA; reverse primer: ACC ACC TgA AAC ATg CAA CA). Primer annealing was set to 58°C, and the melting curve was analyzed to detect any abnormality of the PCR product. All samples were measured in duplicates and expressions were calculated using the relative quantification (ΔΔCq) method. The efficiency of the qPCR reaction was also verified with standard curves.
Statistical analysis
Results are presented as mean±standard error of the mean unless otherwise indicated. Continuous variables were compared using unpaired T-test or one-way analysis of variance, followed by the Dunnett's multiple comparison post hoc test versus the (sham) control group or Tukey's multiple comparisons test. Linear correlation was assessed with Pearson product-moment correlation coefficient. To compare 2 regression lines with each other, the online version of StatTools.net was used (Armitage, 1980). The null hypothesis was rejected if the p value reached statistical significance (*p<0.05, **p<0.01, ***p<0.001).
Results
Lethal renal ischemia-reperfusion injury markers
In the first series of experiments, C57BL/6J mice were subjected to 30-minute unilateral renal ischemia or sham operation followed by removal of the right kidney and 24 hours of reperfusion. The main histological lesions were found in the outer stripe of the outer medulla, and to a lesser degree in the cortical region (Fig. 1). In the renal cortex, tubular cell cytoplasmic vacuolization, brush border loss, pyknosis, tubular dilatation, and flattening of epithelial cells were present (Fig. 1C). In the outer stripe, massive tubular cell necrosis, nuclear loss, tubular cast formation, and immune cell infiltration were observed (Fig. 1G). Quantitative histologic analysis confirmed massive tubular cell necrosis after 30-minute (lethal) ischemia (Fig. 2D).
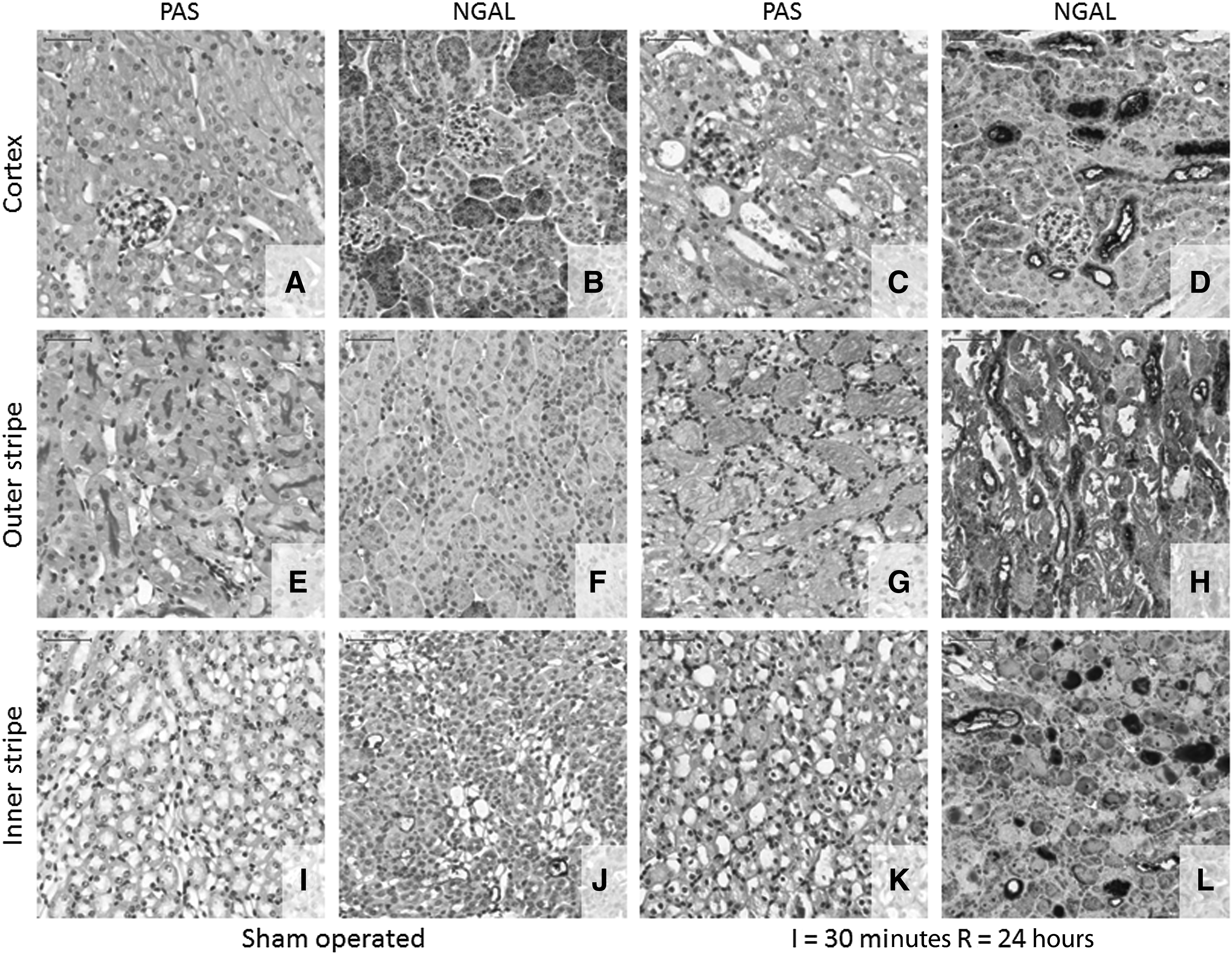
Histopathology and neutrophil gelatinase-associated lipocalin (NGAL) immunohistochemistry (IHC) after a 30-minute renal ischemia-reperfusion (I/R) injury. Representative images scanned at 30× magnification from the cortical
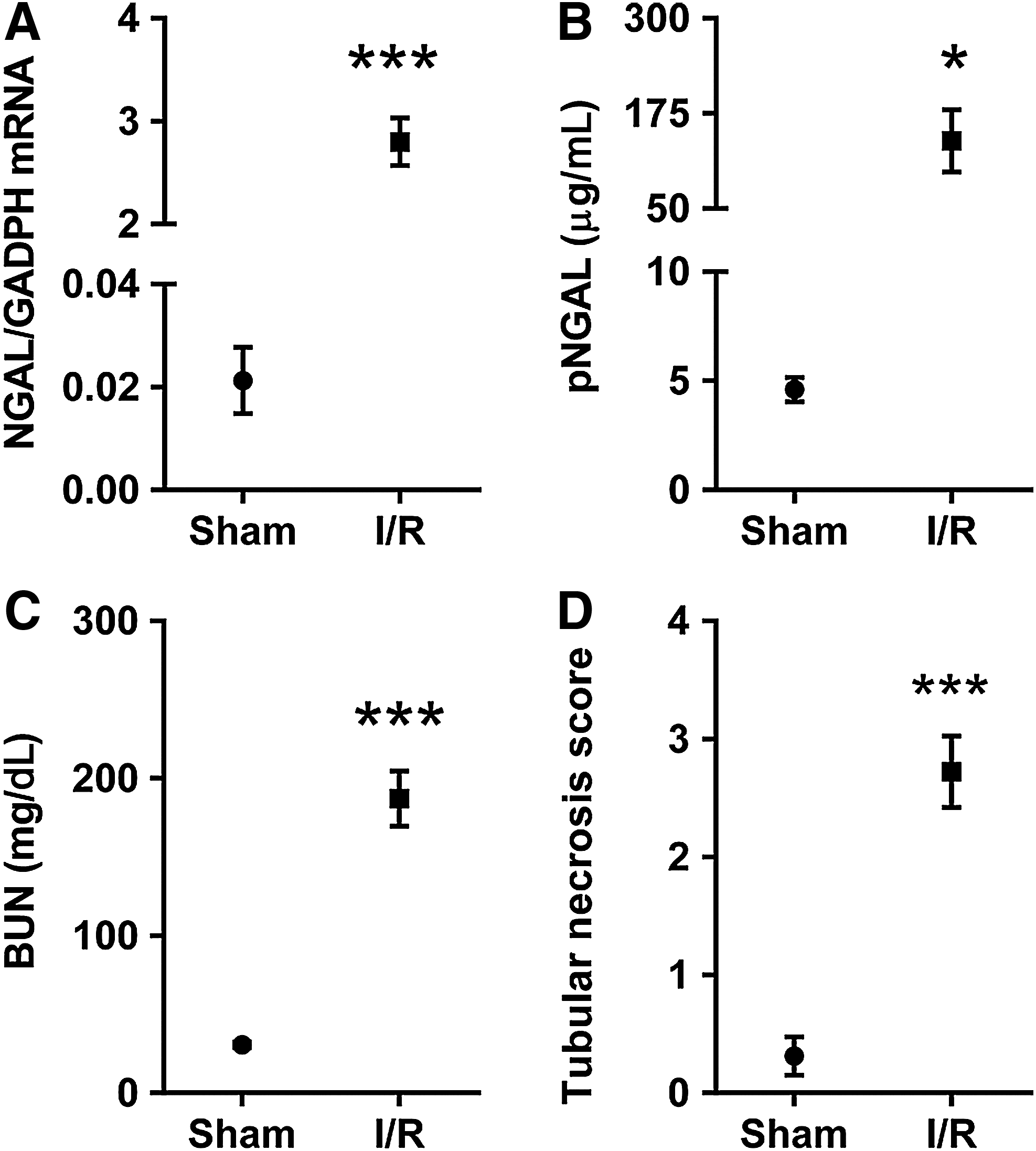
Kidney damage markers after 30-minute I/R injury.
Immunohistochemistry of the renal tubular damage marker NGAL demonstrated a low level of staining in the proximal tubule, and mild staining in the medulla in the kidneys of sham-operated mice (Fig. 1B, F, J). However, there was a strong NGAL specific tubular staining after I/R injury in the outer stripe (Fig. 1H) and cortex (Fig. 1D). Furthermore, whole kidney NGAL mRNA increased 149-fold (p<0.001) (Fig. 2A), and plasma NGAL level increased 30-fold (p=0.05) (Fig. 2B) compared with the sham operated group. In concordance with the morphological results, BUN increased 6-fold 24 hours after 30-minute ischemia compared with the sham-operated group (p<0.001) (Fig. 2C).
Kinetics of sublethal renal ischemia-reperfusion injury markers
To measure the time-course of markers after 20-minute (sublethal) I/R injury, mice were subjected to 20-minute, unilateral renal ischemia or sham operation with various times of reperfusion. After 24 hours of reperfusion, all kidney damage markers increased significantly. Renal NGAL mRNA expression (Fig. 3A) and plasma NGAL protein levels (Fig. 3B) were elevated more than 100-fold (p<0.001) and 16-fold (p<0.001), respectively. Renal function also deteriorated, as reflected by the 4-fold increase (p<0.05) in BUN levels compared with the sham-operated group (Fig. 3C). Renal tubular necrosis score was also significantly elevated 24 hours after 20-minute (sublethal) I/R injury (Fig. 3D). Thereafter, histologic signs of tubular regeneration appeared (Fig. 3E) and renal damage markers started to decrease. The increase in renal NGAL mRNA expression diminished to 66-fold (p<0.05) on day 3, and became nonsignificant after 4 days of reperfusion. Plasma NGAL decreased to the control level already on day 3. On the fourth day of reperfusion, all damage markers returned close to sham values.
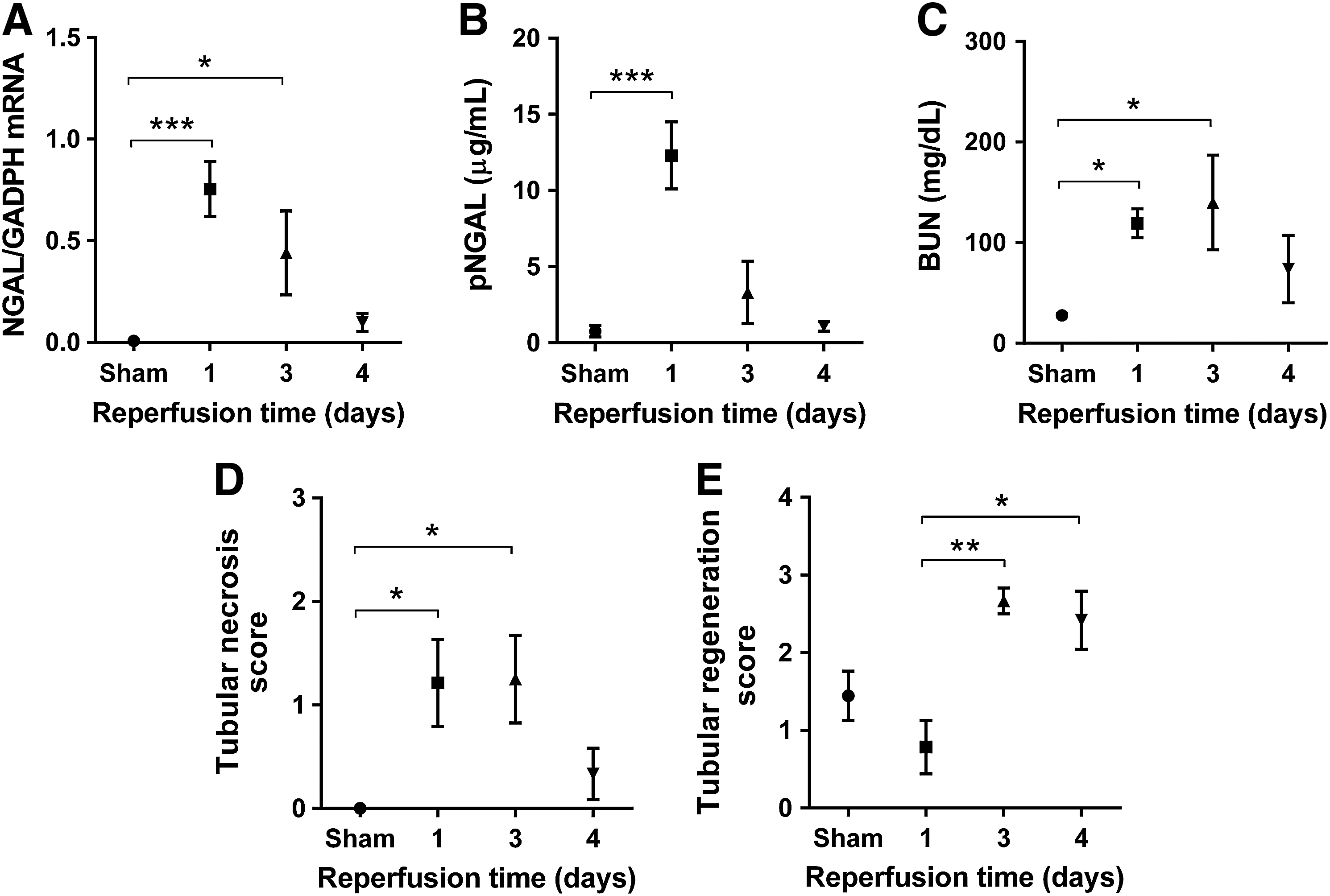
Time-course of kidney damage markers after a 20-minute I/R with various times of reperfusion.
MicroRNA expression changes and time-course of renal miR-17-5p, miR-106a, and miR-21 expressions after renal ischemia-reperfusion injury
The miRNA profile was assessed by the Luminex platform, a novel microbead-based technology. After the exclusion of miRNAs with low fluorescent signal (<100 MFI), 22 miRNAs had been included in the analysis. Five (miR-21, miR-17-5p, miR-106a, let-7c, and miR-99a) out of 22 miRNAs had a significantly different expression relative to the sham-operated group. However, only miR-21, miR-17-5p, and miR-106a changed more than 30% after I/R injury (Fig. 4). These three miRNAs were further analyzed and validated with TaqMan MicroRNA Assays. The qPCR analysis confirmed the significance of the results regarding miR-17, miR-21, and miR-106a (Table 2).
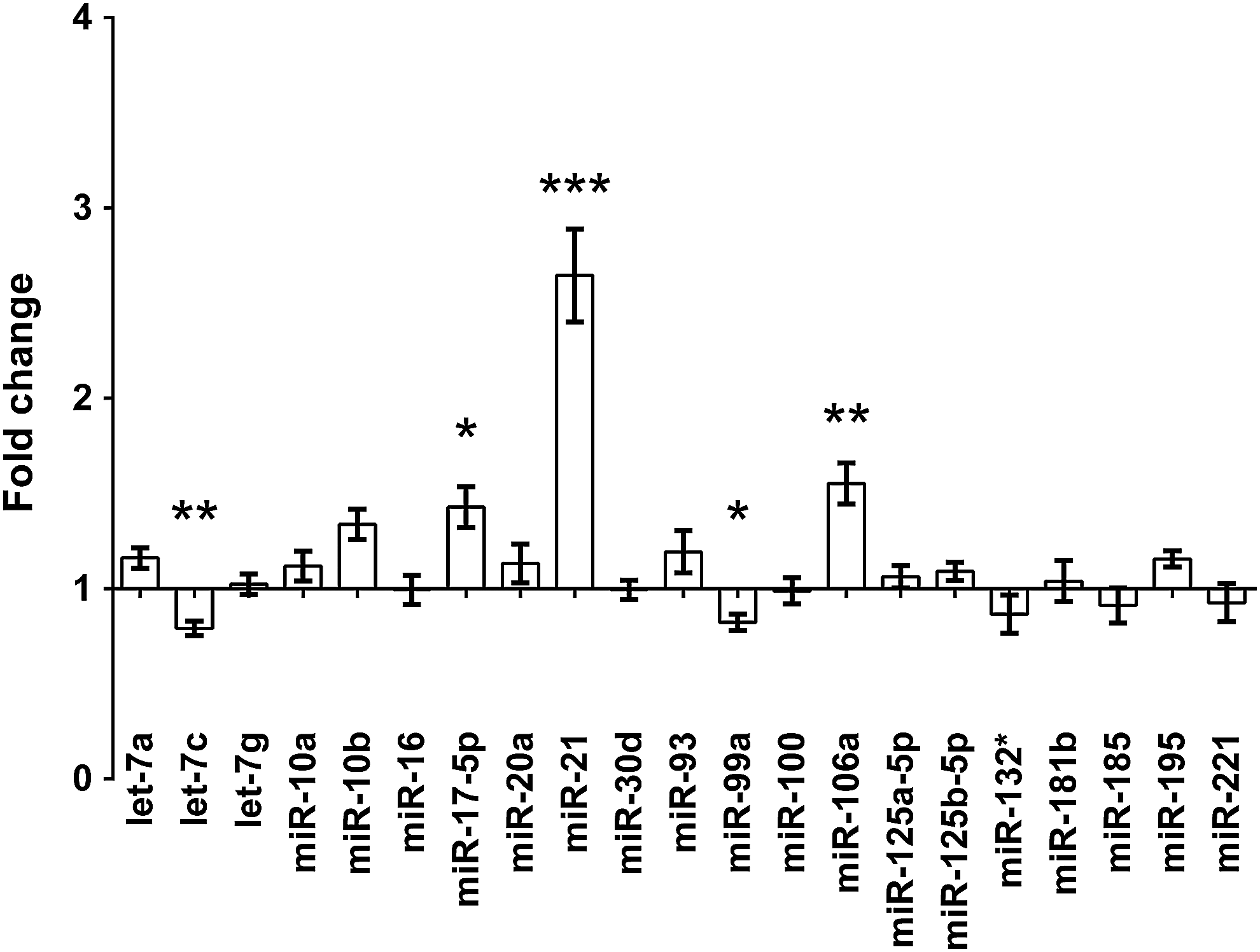
The microRNA (miRNA) expression profile of renal I/R injury, measured on the Luminex multiplex platform. Fold changes were observed after 24 hours of reperfusion following 30-minute ischemia, compared with the sham-operated group. *p<0.05, **p<0.01, ***p<0.001.
p<0.05, **p<0.01 and ***p<0.001.
miRNA, microRNA; qPCR, quantitative polymerase chain reaction; SEM, standard error of the mean.
To determine if the identified miRNAs are upregulated during early or late reperfusion, their expression time-course was evaluated. MiR-21, miR-17-5p, and miR-106a levels increased before any other damage markers, 24 hours after 30-minute (lethal) ischemia (Fig. 5). MiR-17-5p and miR-106a have a similar nucleotide sequence and thus belong to the same miR17-family. As miR-17-5p and miR-106a also had similar expression patterns, we studied miR-17-5p in detail, because its expression was higher than that of miR-106a (Fig. 5A, B). Twenty-minute (sublethal) ischemia was performed to identify the phases of acute kidney injury in which the expressions of miR-17-5p and miR-21 increase. First, miR-17-5p increased significantly (1.6-fold, p<0.05) after one day of reperfusion, and remained significantly elevated (1.7-fold, p<0.01) until the third day (Fig. 6A). On the other hand, renal miR-21 expression was first elevated (3-fold, p<0.001) after 3 days of reperfusion, and remained upregulated (2-fold, p<0.05) on the fourth day (Fig. 6B), when miR-17-5p expression already returned to the sham-operated level.

Time-course of
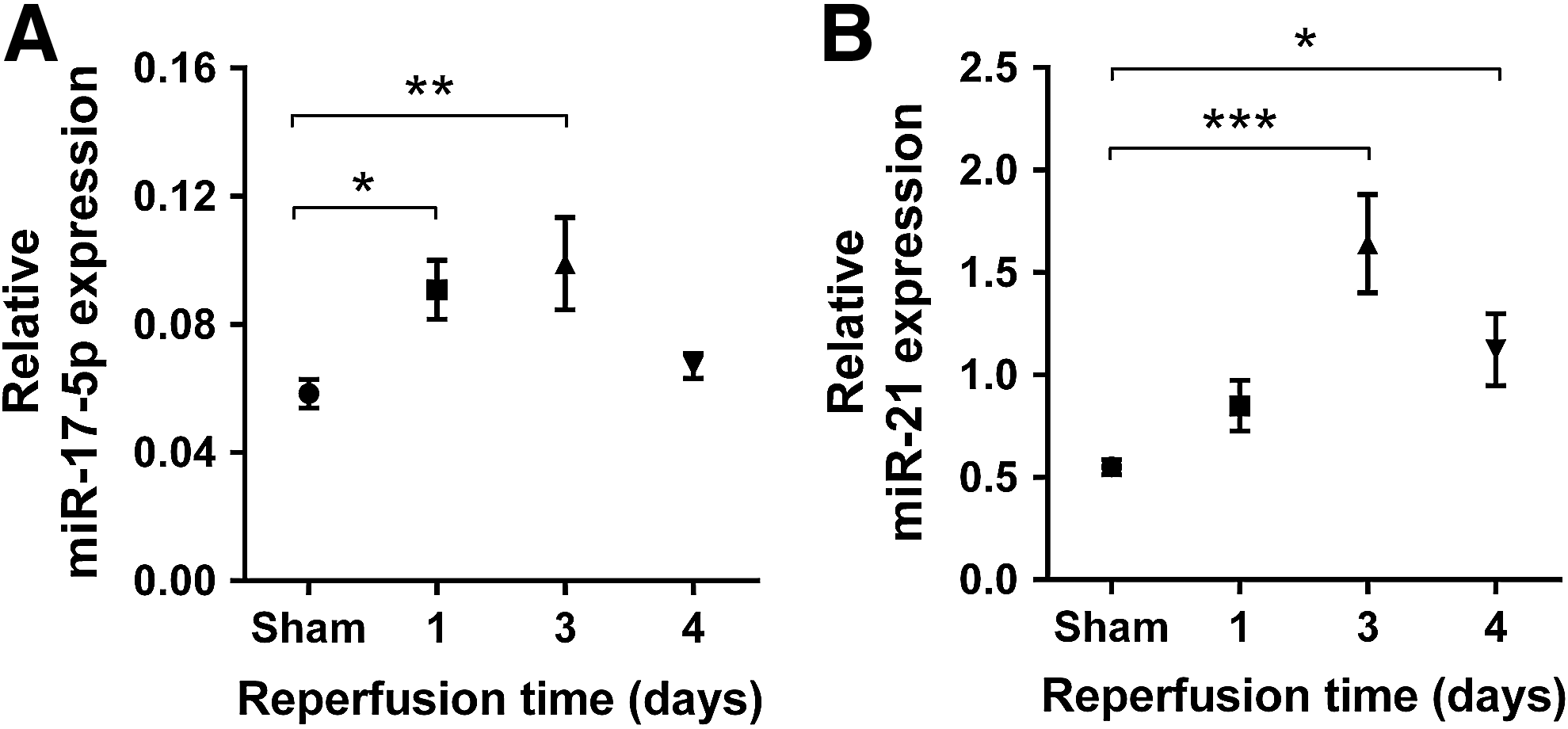
Time-course of
Correlation between the two renal miRNA expressions and other markers of renal I/R injury
We detected significant correlations between renal miR-17-5p and miR-21 expressions at every studied time-point in both the ischemia-reperfusion and sham-operated groups (naive: r2=0.89, p<0.0001, n=20; sham: r2=0.56, p=0.012, n=9; R24h: r2=0.83, p=0.004, n=7; R72h: r2=0.90, p=0.003, n=6; and R96h: r2=0.76, p=0.024, n=6; Fig. 7). However, the slopes of the regression lines were significantly different between the ischemia-reperfusion and sham-operated groups on the third and fourth days of reperfusion (p=0.019 and p=0.001). Moreover, the relationship between the expression of the two miRNAs changed with the duration of reperfusion: the slopes became steeper as the reperfusion time increased (sham: 6.3, p=0.011; 24 hours: 12.2, p=0.004; 72 hours: 15.8, p=0.003; 96 hours: 39.4, p=0.024; Fig. 7).

Correlation of miR-17-5p with miR-21 in the naive, sham–operated, and I/R groups after reperfusion times of 1 day (24 hours), 3 days (72 hours), and 4 days (96 hours).
Discussion
This study provides further evidence for a functional role of miRNAs in the recovery from renal I/R injury. We found for the first time that miR-17-5p and 106a expression increased in the I/R-induced acute kidney injury similarly to that of the previously described miR-21 (Godwin et al., 2010; Saikumar et al., 2012; Xu et al., 2012). Furthermore, elevation of miR-17-5p and miR-106a expression preceded that of miR-21. Based on the correlation between miR-21 and miR-17-5p we speculate that these miRNAs may be part of a regulatory network that can contribute to determine the outcome of I/R injury.
High BUN levels indicated that 30-minute ischemia caused severe renal dysfunction one day after ischemia in C57BL/6 mice similarly to the results of Burne et al. (Burne et al., 2000). Histology indicated severe tubular damage mainly in the outer stripe of the outer medulla, where the S3 segment of the proximal tubule and the medullary thick ascending limb are located, which are both very sensitive to ischemia (Lieberthal and Nigam, 1998). In accordance with previous reports (Schmidt-Ott et al., 2006), tubular origin of renal dysfunction was also verified by NGAL, a sensitive marker of tubular epithelial damage (Mishra et al., 2003), as whole kidney NGAL mRNA and plasma NGAL levels both increased by orders of magnitudes 1 day after ischemia. Similarly to previous studies by us (Heemann et al., 2000; Hamar et al., 2004) and others (Linkermann et al., 2012), 30-minute renal ischemia was lethal in C57BL/6 mice (Zhang et al., 2011). To follow the time-course of miRNA expression during recovery from the ischemia-induced AKI, mice were subjected to 20-minute ischemia leading to reversible injury (Heemann et al., 2000; Hamar et al., 2004).
The sublethal, 20-minute ischemia-induced acute renal failure was also severe. BUN, renal NGAL mRNA expression, and plasma NGAL level significantly increased on the first day after ischemia though the changes were not as great as after 30-minute of ischemia. Furthermore, massive necrosis was also present in the outer stripe of the outer medulla. The changes in the above kidney damage markers became non-significant after 4 days of reperfusion, a period that covered the extension, maintenance, and recovery phases of AKI. Post-ischemic AKI has been divided into three phases: extension, maintenance, and regeneration. The extension phase is characterized by continued ischemic injury. During the maintenance phase, both tubular cell death and regeneration occur simultaneously, while renal function is still at its nadir. As the kidney enters the recovery phase, cell recovery and proliferation balance shifts from cell death to regeneration. Regeneration is associated with improvement of renal function (MOLITORIS, 2003; DEVARAJAN, 2006). Our data support the above stages of AKI time-course. Histologically, tubular regeneration was observed from day 3. Less tubular necrosis, improvement of BUN, and plasma NGAL demonstrated functional and histological recovery on day 4 after ischemia similarly to previous studies (Prachasilchai et al., 2008; Jiang et al., 2012). Therefore, in our sublethal AKI model, the extension and maintenance phases took place during the first 3 days and the recovery phase was initiated on the third day of reperfusion (Regner et al., 2011).
Though miR-17-5p is encoded by the miR-17∼92 cluster on the mouse chromosome 14, and miR-106a is encoded by the miR-106a∼363 cluster on the X chromosome, they share the same seed sequence. Therefore, they are both members of the miRNA-17 family (Xiao and Rajewsky, 2009). Since both miR-17-5p and 106a had a similar expression pattern after lethal ischemia, and miR-17-5p levels were higher, we focused on miR-17-5p. However, miR-106a also has functional relevance in I/R injury as miR-106a was upregulated in an experimental model of ischemic retinopathy in mice (Shen et al., 2008). One day after reperfusion, renal miR-17-5p was similarly overexpressed both after lethal and sublethal ischemia. The high renal miR-17-5p expression was present until the third day of reperfusion (i.e., miR-17-5p expression was upregulated during the maintenance phase). During the recovery phase on day 4, the expression of miR-17-5p positively correlated with the renal expression and plasma levels of NGAL, a tubular epithelial damage marker. However, a causative relationship between miR-17-5p and NGAL expression could be established only by further studies.
Recently, the overexpression miR-17∼92 cluster was associated with renal cyst formation and its inactivation lead to improved kidney function and survival in a mouse model of polycystic kidney disease (Patel et al., 2013). Expression of miR-17 family members may be activated by proinflammatory cytokines (Dews et al., 2006; Petrocca et al., 2008; Brock et al., 2009). MiR-17 influences immune cell function (Venturini et al., 2007) and regulates lymphocyte (Xiao et al., 2008); monocyte (Fontana et al., 2007; Schmeier et al., 2009); and B-cell development (Ventura et al., 2008). These findings point out that the miR-17 family could be involved in the regulation of immune processes during I/R injury. Overexpression of miR-17-5p inhibited hypoxia-induced apoptosis (Taguchi et al., 2008) in both tumors and renal tubules (Yan et al., 2009), and in the heart (Zhou et al., 2013). On the other hand, inhibition of miR-17-5p with antisense oligonucleotides induced apoptosis in lung cancer cells (Matsubara et al., 2007). A microarray study has demonstrated that cardiac miR-17 family members were over-expressed on the second post-ischemic day in a mouse model of myocardial infarction (Roy et al., 2009). These studies suggest that miR-17-5p can be involved in the regulation of recovery not only from renal, but also from myocardial I/R injury.
We observed high miR-21 expression after both 20- and 30-minute renal ischemia, although after sublethal (20-minute) ischemia, miR-21 expression started to increase later. The duration and severity of the maintenance phase is determined by the balance between cell survival and death (DEVARAJAN, 2006). After sublethal I/R miR-21 overexpression was demonstrated mainly in the late maintenance and recovery phases; therefore, miR-21 could have a role in determining the outcome of cell survival/death balance. Similarly to our results, significant up-regulation of miR-21 was found in post-ischemic kidneys in mice in vivo, and in proliferating renal tubular epithelial cells, in vitro (Godwin et al., 2010). However, cell death following ischemia could not be prevented by miR-21 transfection in the latter study.
The correlation between miR-21 and miR-17-5p is an intriguing finding of this study. MiR-17-5p and miR-21 were differentially expressed over the course of functional recovery from the renal ischemic insult in our study. There were significant linear correlations between their renal expression on all days of reperfusion also in the sham-operated mice. Co-overexpression of these 2 miRNAs was also reported in malignant diseases (Kutay et al., 2006; Connolly et al., 2008; Koga et al., 2010; Fernando et al., 2012) and in the aging heart (Zhang et al., 2012), though the correlation between them was not analyzed. We found not only that miR-21 and miR-17-5p expressions strongly correlated with each other after I/R and in the sham control group, but also that the slopes of correlations increased from time-to-time in the maintenance phase of AKI. These results suggest that renal miR-21 and miR-17-5p expression may influence each other.
Research aimed to determine the changes in the expression of miRNAs in response to I/R injury can lead to the development of new diagnostic, prognostic or therapeutic tools. Both the miR-21 and miR-17 families are well conserved among species (Lagos-Quintana et al., 2001; Olive et al., 2010); therefore, it is reasonable to surmise that similar mechanisms operate in humans as well. There are already studies which propose miR-21 as a biomarker in AKI, as its serum and urine levels were altered by renal I/R (Saikumar et al., 2012). The miRNAs identified in our study (miR-17 family and miR-21) were upregulated only in the later phases of AKI, suggesting that they might be connected with the clinical outcome of AKI. Indeed, there are publications to suggest that urine and plasma levels of miR-21 are potential risk markers for AKI progression associated with poor prognosis after cardiac surgery (Du et al., 2013). Regarding the therapeutic possibilities, two recent papers demonstrated that miR-21 could be involved in the renoprotective effect of preconditioning (Xu et al., 2012; Jia et al., 2013). Furthermore, chemically synthesized miR-21 oligonucleotide reduced infarct size, in vivo, in a murine (Yin et al., 2008) and rat (Cheng et al., 2010) model of cardiac I/R, suggesting that miR-21 modulation may have therapeutic potential both in the heart and the kidney. Further studies are however indispensable to investigate the possible clinical implications of miR-17 family in ischemic AKI.
In conclusion, our study demonstrated that besides miR-21, miR-17-5p, and miR-106a are involved in the pathophysiologic processes of the I/R-induced AKI. The miR-17 upregulation occurred during the maintenance phase and was followed later by miR-21 upregulation, which in turn lasted until the recovery phase. The expression of these miRNAs correlated with each other, which finding should be further investigated to get a deeper understanding of a possible relationship between miR-21 and miR-17. The timing of miRNA upregulations suggests that miR-17 and miR-21 could play a role in the recovery phase of the I/R-induced AKI. Considering the progresses made in miRNA research in the kidney (Kaucsar et al., 2010) and miRNA based therapeutic approaches (THUM, 2012) miR-17-5p, miR-106a, and miR-21 could represent novel targets in the treatment of the ischemia-induced AKI.
Footnotes
Acknowledgments
The authors are grateful to Mária Godó and Biomedica Hungary, Ltd. for providing the reagents for the Luminex analysis. Further thanks to Ágnes Cser for the administrative assistance and for taking care of the animal facility. This work was supported by U.S. National Institutes of Health Research grant #R03 TW07069 funded by the Fogarty International Center and the National Institute of Diabetes and Digestive and Kidney Diseases. Further support was provided by the Hungarian Scientific Research Fund (OTKA: K81972, NF69278) and the Health Science Council (ETT: 011-07/2009).
Author Disclosure Statement
No competing financial interests exist.