
Abstract
Activated and regulatory T cells express the negative co-stimulatory molecule cytotoxic T-lymphocyte–associated antigen-4 (CTLA-4) that binds B7 on antigen-presenting cells to mediate cellular responses. Single nucleotide polymorphisms in the CTLA-4 gene have been found to affect alternative splicing and are linked to autoimmune disease susceptibility or resistance. Increased expression of a soluble splice form (sCTLA-4), lacking the transmembrane domain encoded by exon 3, has been shown to accelerate autoimmune pathology. In contrast, an exon 2–deficient form lacking the B7 ligand binding domain (liCTLA-4), expressed by diabetes resistant mouse strains has been shown to be protective when expressed as a transgene in diabetes susceptible non-obese diabetic (NOD) mice. We sought to employ an antisense-targeted splice-switching approach to independently produce these CTLA-4 splice forms in NOD mouse T cells and observe their relative impact on spontaneous autoimmune diabetes susceptibility. In vitro antisense targeting of the splice acceptor site for exon 2 produced liCTLA-4 while targeting exon 3 produced the sCTLA-4 form in NOD T cells. The liCTLA-4 expressing T cells exhibited reduced activation, proliferation and increased adhesion to intercellular adhesion molecule-1 (ICAM-1) similar to treatment with agonist α–CTLA-4. Mice treated to produce liCTLA-4 at the time of elevated blood glucose levels exhibited a significant reduction in the incidence of insulitis and diabetes, whereas a marked increase in the incidence of both was observed in animals treated to produce sCTLA-4. These findings provide further support that alternative splice forms of CTLA-4 affects diabetes susceptibility in NOD mice and demonstrates the therapeutic utility of antisense mediated splice-switching for modulating immune responses.
Introduction
E
The associated genetic links of type 1 diabetes, Grave's, Coeliac and Addison's diseases along with other autoimmune maladies is significant (Kouki et al., 2002; King et al., 2003; Ueda et al., 2003; Blomhoff et al., 2004; Gough et al., 2005). Numerous genes have been mapped with regard to their associative risk to other autoimmune diseases, including a region on the human chromosome 2q33 containing T cell regulatory genes CD28, inducible costimulator (ICOS) and cytotoxic T-lymphocyte-associated antigen-4 (CTLA-4) (Ueda et al., 2003; Greve et al., 2004). Furthermore, an equivalent region in chromosome 1 containing murine orthologues of cd28, icos, and ctla-4 is mapped to the susceptibility of spontaneous type 1 diabetes in the non-obese diabetic (NOD) mouse (Ueda et al., 2003; Greve et al., 2004; Vijayakrishnan et al., 2004; Araki et al., 2009).
Activated CD4+ and CD8+ T cells express CTLA-4 and its homologue CD28. Both bind to B7 on antigen presenting cells (APCs), though CTLA-4 does with higher affinity with each have opposing effects on T cell stimulation and activity (Krummel and Allison, 1995). CD28 provides a positive co-stimulatory signal while CTLA-4 imparts a negative signal. The regulation of T cell responses impacting their number and function activity depends largely on which co-stimulatory signal prevails (Kuhns et al., 2000). While positive costimulation through CD28 results in the rapid expansion of effector cells able to combat pathogens, negative signaling through CTLA-4 plays a critical role in maintaining T cell homeostasis and regulating peripheral tolerance to self-antigen (Perez et al., 1997) as well as the induction and function of natural and adaptive T regulatory cells (Tang et al., 2004; Vasu et al., 2004; Read et al., 2006). These regulatory functions are evident in mice lacking CTLA-4 expression (ctla-4–/–) or function, which rapidly develop lymphoproliferative disease and generalized autoimmune destruction of tissues (Tivol et al., 1995).
CTLA-4 negative co-stimulation can prevail over CD28 positive costimulation due to several structural and functional characteristics. On the T cell membrane surface the external domain of CTLA-4 can spatially interfere with the colocalization and sustained occupation of CD28 within the immune synapse (Saito et al., 2010; Yokosuka et al., 2010; Pentcheva-Hoang et al., 2004). The inhibitory function of the intracellular portion of CTLA-4 is via interactions with various signaling phosphatases and kinases within the T cell receptor (TCR) complex (Guntermann and Alexander, 2002; Schneider et al., 2002). The external and internal structurally dependent regulatory functions of CTLA-4 combine to restrain TCR signaling which results in the blockade of cell cycle progression reduced interleukin-2 (IL-2) production and ultimately diminished T cell responsiveness and effector activity (Krummel and Allison, 1995, 1996).
Both humans and rodents produce alternatively spliced forms of CTLA-4, lacking either extracellular or intracellular domains. Over-expression of one form or the other has been linked autoimmune disease susceptibility or resistance (Liu et al., 2003; Ueda et al., 2003; Simone and Saverino, 2009). A soluble form of CTLA-4 (sCTLA-4) that excludes intracellular portions of the molecule are naturally produced by humans and rodents (Oaks et al., 2000; Ueda et al., 2003) and overexpression in mice can lead to the induction and exacerbation of autoimmune diseases including experimental autoimmune encephalomyelitis, lupus and diabetes (Ueda et al., 2003; Ichinose et al., 2013; Liu et al., 2003, 2012). Additionally, diabetes susceptibility in the NOD mouse has been linked to a single-nucleotide polymorphism (SNP) that diminishes the ability to produce a splice variant called the ligand independent form (liCTLA-4) produced by the exclusion of exon 2 in the mature mRNA (Ueda et al., 2003). When liCTLA-4 was made to be expressed in ctla-4–/–t cells by recombinant retroviral vector these cells were more potently inhibited in aspects of proliferation and cytokine production than ctla-4–/– cells made to express the full-length form (flCTLA-4). Moreover, there is enhanced expression of liCTLA-4 in the memory and regulatory T cells of a diabetes-resistant congenic stain of NOD compared to susceptible NOD mice (Vijayakrishnan et al., 2004; Chikuma et al., 2005; Araki et al., 2009). However, recently it has been shown that suppression of autoimmune diabetes is dependent on the expression of both the ligand independent and full-length forms of CTLA-4 (Stumpf et al., 2013).
Taken together these data strongly suggest that expression of the alternative splice forms of CTLA-4 play a significant role in the resistance and susceptibility to T cell-mediated autoimmune disease including type 1 diabetes. In this study, we use a novel antisense approach to mask pre-mRNA splice recognition sites and redirect the splicing machinery to skip selected exons. Here we show for the first time that antisense can be used to induce the expression of alternative splice forms of CTLA-4, including liCTLA-4 found in diabetes resistant mice or sCTLA-4 found to accelerate autoimmunity (Ichinose et al., 2013), to result in the suppression or augmentation of diabetes, respectively in the NOD mouse. These findings further support the observation that expression and activity of CTLA-4 gene products contribute largely to NOD mouse diabetes susceptibility.
Materials and Methods
Mice
Female NOD mice 7 weeks of age were obtained from Jackson laboratories and housed in microisolator cages three to five per cage during acclimatization and treatment periods in the laboratory animal resource center at Oregon State University. Animals were exposed to 12-hour light/dark cycle in temperature- and humidity-controlled environment and allowed access to commercially available pre-autoclaved sterilized rodent diet and autoclaved sterilized water. Temperature controls were set to maintain temperatures at 18°C to 26°C. Autoclaved caging, water bottles, and storage containers were used to avoid exposure of animals to any contaminated materials. The animal studies were used in accordance with the guidelines established in the Guide for the Care and the Use of Laboratory Animals of National Research Council and monitored by the institutional animal care and use committee, Oregon State University. Animals were treated by intraperitoneal injection (i.p.) 150–200 μg per injection] with peptide-conjugated phosphorodiamidate morpholino oligomer (PPMO) [1.0 μg/μL] suspended in sterile phosphate-buffered saline (PBS). Blood glucose levels were measured using 5–10 μL of blood obtained via needle stick to the saphenous vein and an ACCU-CHEK Advantage monitor and Comfort Curve strips (Roche Diagnostics). The monitor was calibrated weekly. Animals were terminated after two consecutive blood glucose readings above 250 mg/dL.
Antisense reagents
Murine CTLA-4 antisense targeting sequences were determined from the genomic sequence in GenBank accession number AF142145 and were synthesized by Sarepta Therapeutics, Inc. chemistry group as previously described (Nelson et al., 2005). A cell-penetrating peptide composed of (RXR)4XB, where R=L-arginine, X=6-aminohexanoic acid, and B=beta-alanine, was conjugated on the 5′ end of each PMO, respectively. The PPMO sequences specific for the murine ctla-4 mRNA are as follows where AUG indicates sequence encompassing the start codon for translational initiation, SA indicates splice acceptor sequence flanking the exon side of an intron–exon boundary, and SD indicates the splice donor sequence flanking the exon side of an exon–intron boundary for the numerical exon designation:
Control: 5′ CCT CTT ACC TCA GTT ACA 3′′ CTLA-4 AUG: 5′ CCA AGA CAA GCC ATG GCT GG 3′ CTLA-4 SA3: 5′ CCG GGC ATG GTT CTG GAT C 3′ CTLA-4 SA2: 5′ GGT TGG GTC ACC TGT ATG G 3′ CTLA-4 SD2: 5′ GTA AGG CGG TGG GTA CAT G 3′ CTLA-4 SD3: 5′ CAT CTT GCT CAA AGA AAC AG 3′
Splenocyte culturing and in vitro antisense treatment
Splenocyte cell suspensions in Dulbecco's modified Eagle's (DME)/high glucose and 1% fetal bovine serum (FBS) media were made by passing macerated spleens though cell strainers. Suspensions were washed twice, and cultured in mouse complete media [MCM=RPMI plus 10% FBS (Hyclone®), 1% antibiotic-antimycotic stock (Cellgro®) (10,000 IU penicillin, 10,000, μg/mL streptomycin and 25 mg/mL Amphotericin B), 2 mM
Western blotting
Splenocytes (5×106 cells) were stimulated with 5 μM Con A in the presence or absence of 5 μM CTLA-4-SA2 for 48 hours and lysed in total protein extraction buffer (Millipore) as per the manufacturer's instructions. Polyvinyl difluoride membranes were blocked with 5% milk in 0.1% Tween-PBS. Ctla-4 was detected using an anti-Ctla-4 antibody (C-19, SantaCruz) at 1:200 in 1.5% block buffer and donkey-anti-goat immunoglobin G–horseradish peroxidase (IgG-HRP) (SantaCruz) in 5% block buffer. Actin was probed using anti-actin (Sigma) in 5% block buffer and goat-anti-rabbit IgG-HRP (SantaCruz).
RNA extraction and reverse transcription polymerase chain reaction
Splenocyte or pancreatic lymph node RNA was extracted using Qiagen's RNeasy Mini Kit (Qiagen USA) as per manufacturer's protocol. SuperScript™ 3 One-Step RT-PCR System with Platinum® Taq DNA Polymerase kit (Invitrogen) was used according to the manufacture's protocol to convert isolated RNA to cDNA and amplify. Targeting the flctla-4 transcript, the following primers were designed from GenBank accession number NM_009843: F 5′-ACA CAT ATG TAG CAC GTA CCT TGG A-3′and R 5′-GGA ATT TTG CAT CCA GCT TTC TAT-3′. To amplify a portion of the ICOS transcript, the primers F 5′-AAG CCG TAC TTC TGC CGT-3′ and R 5′-CCA CAA CGA AAG CTG CAC-3′ were designed from GenBank accession number BC034852. Primers targeting the CD28 transcript, F 5′-ATG ACA CTC AGG CTG CTG-3′ and R 5′-GCA AGC CAT AAC AAA ACA G-3′, were designed from GenBank accession number BC034852. Real-time quantitative polymerase chain reaction (PCR) to determine induced levels of liCTLA-4 in vivo was conducted using BioRad iScript one-step real-time PCR kit for probes and primer set F 5′- GCCTTTTGTAGCCCTGCTCA-3, R 5′- TCAGAATCCGGGCATGGTT-3′ and probe FAM 5′-TTCTTTTCATCCCAGTCTTCTCTGAAGATCCA-3′ as previously described (Vijayakrishnan et al., 2004). All reverse-transcription and PCR runs were completed on BioRad iCycler. The reverse transcriptase PCR was ran to determine the compound activity according to the following protocol: reverse-transcription reaction at 55°C for 30 minutes, Taq polymerase activation at 94°C for 2 minutes, 35 cycles of 94°C denaturation for 30 seconds, annealing for 45 seconds at 56°C, and extension for 1 minute at 68°C, and a final extension for 10 minutes at 68°C. The amplicons were analyzed by either agarose gel, by the Lab Chip GX (Caliper Life Sciences, Inc.) using HT 1Kb DNA LabChip Dual protocol DNA Analysis and Quantitation Reagent Kit (Caliper Life Sciences Inc.) as per manufacturer's protocol.
Flow cytometry
Magnetic bead-purified T cells (Miltenyi Biotec) were labeled with carboxyfluorescein-succinimidyl-ester (CFSE) [2 μM] and stimulated with plate bound -CD3 antibody [5 μg/mL]. Cells were cultured for 48 hours with either PPMO (5 μM) or α-CTLA-4 antibody [5 μg/mL] or isotype. Cells were stained with CD69 fluorescein isothiocyanate (Caltag). Surface staining and CFSE dilution were examined by flow cytometry gating on live cells using 7-aminoactinomycin D (7-AAD) (Calbiochem). Intracellular cytokine staining (ICS) was conducted as previously described (Marshall et al., 2007). Data were acquired using Beckman Coulter FC-500 and analyzed using Winlist (Verity) or FlowJo (Tree Star). Functional uptake of the PPMO was analyzed in resting or plate bound α-CD3 activated T cell using splenocytes isolated from the enhanced green fluorescence protein (EGFP)-654 transgenic mouse that reports antisense mediated exon-skipping by blocking of an cryptic site that mediates aberrant splicing and interruption of the EGFP open reading frame (Sazani et al., 2002).
Adhesion and blocking assays
To examine the adhesion activity, purified splenic derived T cells (Mitenyi) were pretreated with PPMO (5 μM) for 16 hours or not and plated in triplicate into a 96-well plate pre-coated with ICAM-1 (12.5 μg/mL) and then stimulated with α-CD3 with or without α-CD28 or α-CTLA-4 (CD152) (10 μg/mL) costimulation for 1 hour. Loose cells were removed by inverting the plate remaining cells were removed and enumerated using a hemocytometer.
Soluble CTLA-4 activity in pre-filtered and five-fold concentrated (50 kDa and 10 kDa centricons, respectively) culture supernatants (SN) derived from (0.5–5.0 μM) SA3 PPMO treated NOD splenocytes stimulated with concanavilin A (4 μg/mL) for 48 hours was assessed by competitive blockade of CD86 staining. Freshly isolated B6 splenocytes activated with lipopolysaccharide [500 ng/mL] for 24 hours were incubated with either PPMO treated culture SN or CTLA-4 Fc chimera [0.125 – 0.5 μg] in 500 μL FACs buffer for 3 hours. The cells were fixed (4% paraformaldehyde), washed, Fc blocked, stained with anti-CD86-PECy5 (eBioscience) and examined by flow cytometry for mean fluorescent intensity (MFI) of CD86. Inhibition was determined as a percent reduction in the MFI compared to B6 splenocytes incubated with SN from PBS control treated cells.
Histological assessment of islet insulitis
Pancreata were frozen in optimal cutting temperature compound. A minimum of five, 5-μm sections, each 200 μm apart, were cut for each tissue block. Sections were fixed in formalin, and stained with H&E. Each islet was scored as: no insulitis or intra-insulitis (>50% infiltration).
Statistics
One-way analysis of variance (ANOVA) with Tukey's multiple comparison and the student's test t-test were performed using Prism Software (GraphPad). A value of P<0.05 was considered statistically significant.
Results
Targeted splice switching of CTLA-4
The genomic architecture of ctla-4 is shared by mouse and humans, both contain 4 exons encoding distinct functional domains within the receptor: exon 1, a leader sequence for protein trafficking in the cell; exon 2, an IgV-like domain that binds B7-1 (CD80) and B7-2 (CD86) ligand on APCs; exon 3, a transmembrane domain; and exon 4, a cytoplasmic tail that interacts with intracellular signaling molecules (Fig. 1A).

Redirected splicing via antisense-induced splice switching produces different splice forms of cytotoxic t-lymphocyte antigen 4 (ctla-4).
Initially we designed antisense reagents to target exon splice donor (SD) or splice acceptor (SA) sites within the murine ctla-4 gene sequence derived from GenBank accession number AF142145. PMO chemistry was chosen as the antisense modality in these studies because an uncharged backbone affords oligomer nuclease resistance and does not induce RNAse H activity that would cause degradation of the targeted mRNA, and thus destroy expression of any splice-altered gene products (Stein et al., 1997). In addition, a cell penetrating peptide was conjugated to each of the oligomers (PPMOs). The conjugated-peptide readily delivers PMO into primary murine dendritic, macrophage, and T cells and effectively induces splice alterations in vitro with limited impact on cell viability (Marshall et al., 2007). Antisense oligomers designations are given as SD or SA and then the exon targeted (e.g., SA2 targets the splice acceptor site of exon 2). The induction of alternate CTLA-4 splice forms was examined by RT-PCR using total cellular RNA isolated from NOD splenocytes after stimulation and treatment with PPMO in culture for 24 hours. Alterations to the size of products were analyzed by agarose gel or capillary and it was observed that targeting of splice acceptor sites were more efficient at altering splicing compared to targeting splice donor sites (data not shown).
We further sought to determine if the splice alterations were as predicted, and thus would maintain fidelity of the open reading frame (ORF) to produce a functional protein. PCR amplicons were gel isolated, cloned, and cDNA sequenced and translation of the ORF made in silico. The predicted sequence after antisense-induced excision of exon 2 was obtained using both the SD2 and SA2 PPMO. Sequence analysis showed precise excision of exon 2 producing liCTLA-4 with the IgV region removed and ORF reestablished at exon 3 (Fig. 1C, upper alignment). Targeting of exon 3, some clones (3/12) obtained from cells treated with SD3 or SA3 PPMO did exhibit the predicted splice alteration. However, the dominant sequence (9/12) was similar to a natural occurring splice form of CTLA-4 form expressed by rats (accession number U90271) where a cryptic acceptor site in exon 4 is used and an alternative ORF established, suggesting that this might be a soluble form of CTLA-4 conserved in rodent evolution, but not naturally or highly expressed by NOD mice (Fig. 1C, lower alignment).
Verification of protein production was conducted using western blot analysis with anti-CTLA-4. A protein with size similar to that predicted for the liCTLA-4 was detected in Splenocyte preparations after in vitro treatment with SA2 (Fig. 1D). Western blot detection of the soluble form of CTLA-4 proved to be less sensitive, so we employed a competitive blockade assay to determine if the antisense induced CTLA-4 was capable of binding to CD86. Soluble CTLA-4 activity in culture supernatant derived from SA3 PPMO treated and stimulated splenocytes was assessed by competitive blockade of CD86 staining. Splenocytes served as target binding cells for SA PPMO treated culture supernatant or CTLA-4 Fc chimera antibody. The target cells were fixed and examined for competitive blockade of anti-CD86-PECy5 staining by flow cytometry (Fig. 1E). The percent inhibition of CD86 staining compared to control supernatant exhibited a dose-dependent response to the treatment of SA3 suggesting that an increase in the production of soluble CTLA-4 was due to increased targeted alternative splicing activity.
Analysis of splice recognitions site surrounding exon 2
Targeting of sequences surrounding the intron/exon boundary was conducted to determine if sequence position for inducing liCTLA-4 was optimized. PPMOs flanking both sides of the intro-exon boundary as well sequences inside the intron and exon were tested (Fig. 2A). The initial SA2 sequences positioned at the exon side of the intron–exon boundary were most effective at inducing liCTLA-4. Surprisingly, however, an oligomer sequence positioned 67 bp into the exon was nearly as potent as PPMO positioned at the boundary, suggesting a putative splice signal sequence for the splicing machinery resides inside the exon (Fig. 2B).
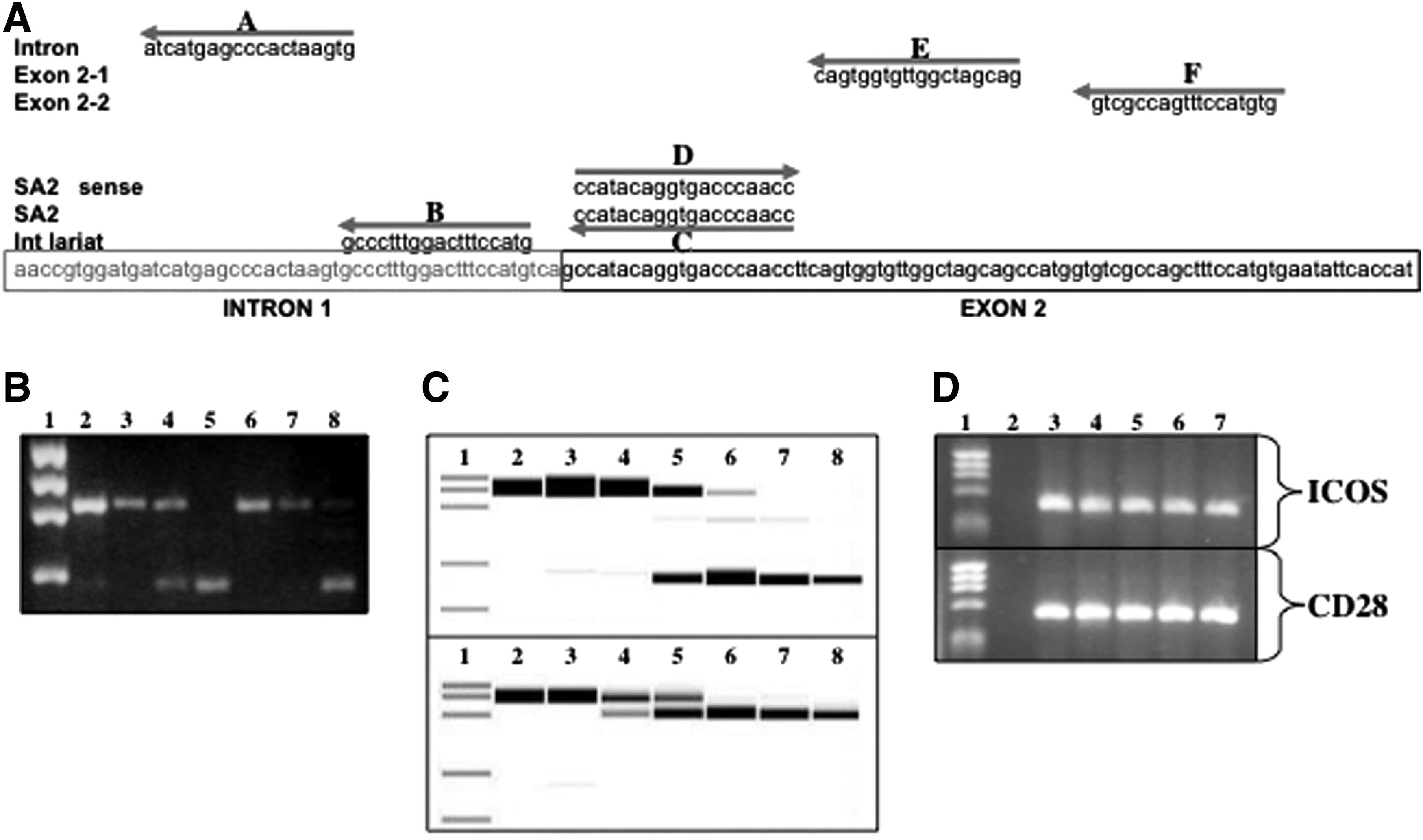
Antisense induced skipping of Exon 2 is position and concentration dependent and target specific. PPMO targeting different regions surrounding the intron 1 exon 2 pre-messenger (m)RNA boundary were used to optimize induction of exon 2 skipping. Cultured NOD splenocytes were stimulated and treated with the various PPMO [5 μm] for 24 hours and RT-PCR conducted with isolated mRNA.
Specificity and dose dependent targeting of CTLA-4 pre-mRNA
The splice-altering activity of the SA2 and SA3 PPMO was examined to determine if induction of the CTLA-4 alternative splice forms behave in a dose-dependent manner. We have observed detectable splicing in cells treated with as little as 0.1 μM PPMO producing ∼10%–20 % of the targeted form and ∼100 % altered product was observed at concentrations between 2 and 5 μM (Fig. 2C, SA2 upper panel and SA3 lower panel). However, using quantitative RT-PCR the percent conversion of splicing to the liCTLA-4 form was observed to be only ∼70% at a treatment dose as high as 10 μM (Fig. 2C, lower panel).
Splicing profiles of other costimulatory molecules structurally, sequentially, and functionally related to CTLA-4 were examined to determine if they were affected by treatment with sequence-specific CTLA-4 PPMO. RNA isolated from cultured NOD splenocytes after treatment with PPMO and examined by RT-PCR using primer pairs specific for CD28 or ICOS to determine if the splice patterns of these mRNA remained normal (Fig. 2D). No alterations to these molecules were detected using PPMO SA2 or SA3 (data not shown), demonstrating the specificity of these reagents.
Induction of altered forms of CTLA-4 affects T cell activity in vitro
After establishing the molecular requirements for targeting the pre-mRNA, we next addressed the biological effects that splice alterations might have on T cell activity in vitro. It has been shown that triggering through CTLA-4 influences proliferation and adhesion of activated T cells (Krummel and Allison, 1996; Schneider et al., 2005). Two separate assay systems were employed to determine if induction of liCTLA-4 would cause T cells to behave similar to activated T cells triggered through CTLA-4 via an agonist antibody (Fig. 3). Proliferation in response to α-CD3 and α-CD28 stimulation of purified CD4+ T cells labeled with CFSE after treatment with PPMO SA2 or SA3, α-CTLA-4 agonist or isotype antibody was examined by flow cytometry (Fig. 3A). Control PPMO, SA3, and isotype treated cells all exhibited similar proliferation. It has been demonstrated in previous studies, α-CTLA-4 inhibited T cell proliferation in our assay. We observed that induction of liCTLA-4 by antisense exon-skipping also inhibited proliferation. Increased adhesion to ICAM-1 is another effect on T cell function recently described for triggering CTLA-4 (Schneider et al., 2005). Cells were plated in wells previously coated with ICAM-1 protein and stimulated with α-CD3 in combination with SA2, SA3, α-CTLA-4 antibody, α-CD28 antibody, or SA2 alone. Nonadherent cells were removed by plate inversion and remaining cells enumerated by hemocytometer count. Again, treatment with SA2 recapitulated α-CTLA-4 effects on T cell activity (Fig. 3B). Interestingly, treatment with SA3 also influenced cellular adhesion, but in this case, it was decreased.

PPMO targeting altered CTLA-4 splice forms produce different T cell activities in vitro.
We examined other effects described for α-CTLA-4 triggering including the expression of activation markers CD25 (data not shown) and CD69 (Fig. 3C). In both experiments, treatment with SA2 decreased protein expression on the surface of stimulated T cells similar to treatment with α-CTLA-4. Effects on cytokine production were examined by ICS. PPMO treatment with SA3 or SA2 produced opposite and significant results with regards to production of TNF-α by stimulated CD8+ T cells compared to control and each treatment. Examination of other cytokines (e.g., IL-2 and γ-IFN exhibited similar trends in cytokine production but not significantly different levels; data not shown).
Effects of PPMO treatment on diabetes in NOD mice
Having confirmed that antisense targeted induced splice-switching influences T cell behavior in vitro, we postulated that using the antisense reagents in vivo to alter the expression profile of liCTLA-4 or sCTLA-4 in the NOD mouse would affect the outcome of diabetes. Female mice were treated with splice altering PPMO SA2 or SA3 or a PPMO targeting translational initiation site of the CTLA-4 protein (AUG). Animals (n=12 per group) were treated by (i.p.) injection with PPMO (a previously determined maximum feasible dose of 150 μg (data not shown) in 200 μL saline for 2 weeks starting at age 8 weeks and blood glucose levels (b.g.l.) monitored twice weekly. Animals exhibiting a b.g.l. greater than 250 mg/dL on two consecutive measurements were considered diabetic. Although animals receiving SA2 did develop diabetes to the same extent as saline and AUG treated animals, a delay in the time to onset of disease was observed (1.8 weeks later). Surprisingly, the animals treated with SA3 developed disease at a point approximately 2 weeks earlier than controls. Moreover, a greater percentage of animals developed disease when treated with SA3 (83 % compared to an average of 64% in the other treatment groups) indicating that induction of soluble CTLA-4 in the NOD mouse exacerbates disease (Fig. 4A). These results demonstrate that alteration to CTLA-4 splicing in the NOD mouse can affect the course of spontaneously occurring diabetes. However, because the onset of diabetes in the female NOD does occur within a fairly wide age window (∼12–27 weeks for saline controls in this experiment) we concluded that the 2-week treatment from age 8 to 10 weeks may have missed the therapeutic window for many of the animals. Statistical analysis using a cut off of a b.g.l. of 180 mg/dL did show a significant difference in the treatment groups compared with control (p=0.04).
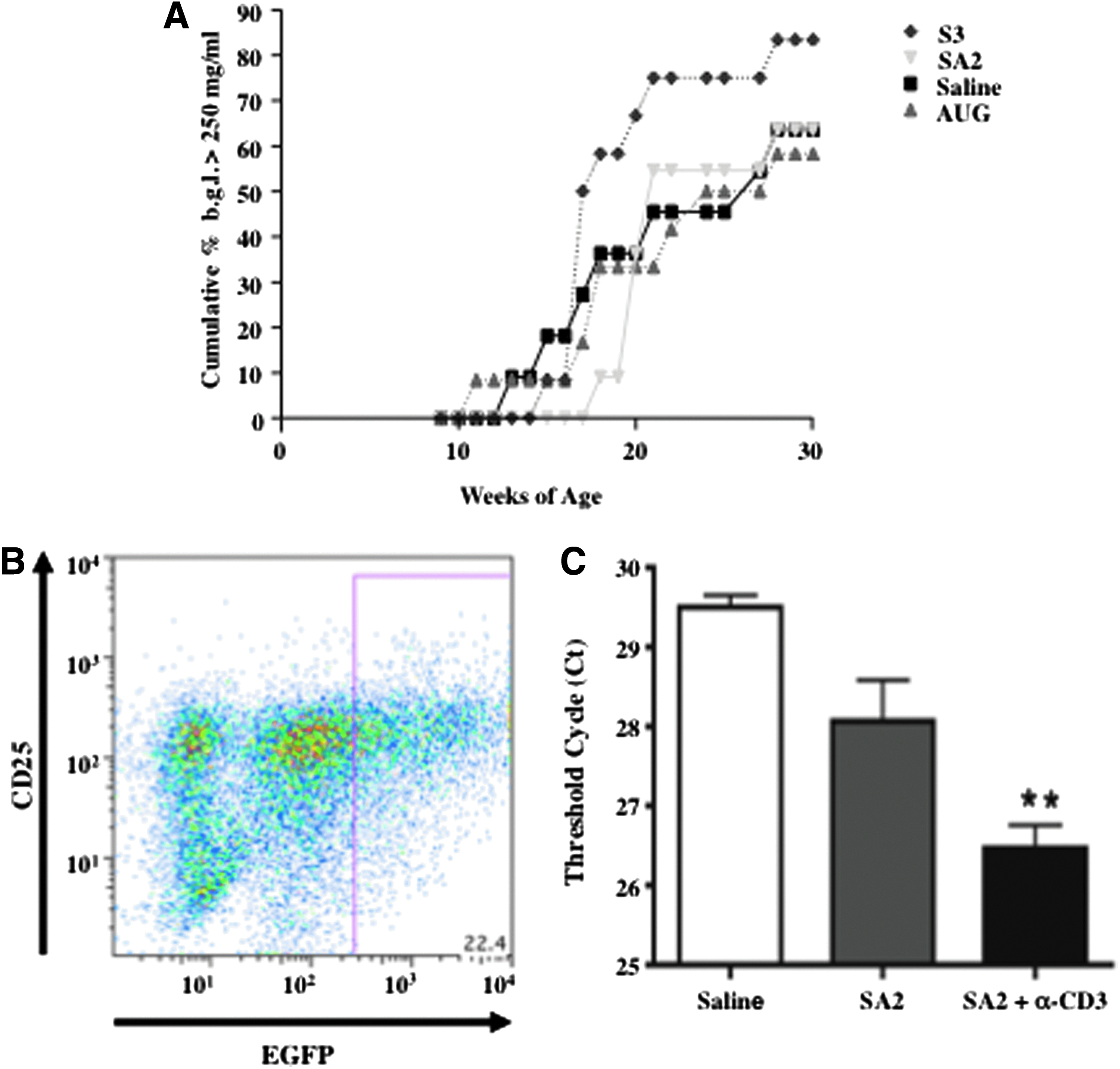
Occurrence of diabetes in NOD mice treated with antisense CTLA-4 targeted splice-switching PPMO and in vitro and in vivo activation dependent enhanced antisense activity.
Activation dependency of PPMO antisense activity of in T cells in vitro and in vivo
While testing the uptake properties of various arginine-rich PPMO, we had made the prior observation that delivery was enhanced in T cells following activation stimulus (Marshall et al., 2007). However, those previous data were generated utilizing a fluorescent conjugated PPMO. Therefore, we sought to determine if the same observation of enhanced PPMO uptake could be extended to an increase in functional antisense activity also dependent on the activation state of the T cells. Employing an in vitro assay system with splenocytes isolated from the enhanced green fluorescence protein (EGFP)-654 transgenic mouse that reports antisense mediated splice-switching to restore the EGFP transgene open reading frame (Sazani et al., 2002) we examined this question. Splenocytes were cultured for 48 hours in wells previously coated with α-CD3 or PBS with or without treatment of PPMO targeting the 654 cryptic splice site, and then stained for CD25 and examined by flow for EGFP expression. Figure 4B shows that the majority of EGFP expressing splenocytes co-stain for the T cells activation marker CD25, confirming the results that activation enhances PPMO uptake and functional antisense activity.
To confirm the in vivo alternative splice-altering capability of the liCTLA-4 PPMO, female NOD mice (n=6 per group) were injected via i.p. route with SA2 or saline control once a day for 10 days. On day 10, three mice from each of the treatment groups were injected via intravenous route of administration with α-CD3 antibody as this has been shown to induce activation of tumor specific T cells in vivo (Ellenhorn et al., 1988). Four hours after antibody treatment, the pancreatic lymph node was harvested from each animal and processed for isolation of total mRNA. RT-PCR analysis was used to confirm if antisense treatment resulted in an increased expression of liCTLA-4 (Fig. 4). The resulting cDNA was examined by agarose gel electrophoresis to determine if a band of the predicted size of liCTLA-4 was observable (data not shown). All animals receiving the SA2 treatments with or without the addition of α-CD3 exhibited cDNA bands of the approximate predicted size for both flCTLA-4 and liCTLA-4, while no observable liCTLA-4 band was present in any of the saline treated animals (data not shown). Both upper and lower bands were later verified by sequencing to be the CTLA-4 splice forms. Analysis by quantitative PCR with a FAM probe specific for the liCTLA-4 was performed to measure level of expression for the various treatment groups (Fig. 4C). Treatment with PPMO alone produced a marked increase in expression of liCTLA-4 over the saline treatment; however, there was only a significant increase in liCTLA-4 expression (p=0.01, ANOVA) in animals receiving α-CD3 in combination with the PPMO. We verified that the α-CD3 treatment alone does not induce liCTLA-4 expression in NOD mice by RT-PCR (data not shown), thus confirming that PPMO is responsible for in vivo expression of the isoform and that activated cells are more potently effected by the splice-altering PPMO when activated.
We therefore concluded, to produce a therapeutic benefit from the antisense induced liCTLA-4, the PPMO need to be administered during the early stages of diabetogenic autoimmune T cell activation and prior to onset of fully developed diabetes. A window that is not precisely age related for each individual NOD mouse, so in order to target this therapeutic window we conducted a subsequent study where animals were monitored for b.g.l. twice per week beginning at age 8 weeks. Animals, regardless of their age at the time of presenting a b.g.l. between 150 and 180 mg/dL, were randomlyz placed into treatment groups (n=10–17) and administered either saline as a control or PPMO—SA2 to induce the therapeutically beneficial liCTLA-4 splice form to inhibit onset of autoimmune disease, SA3 to induce the autoimmune disease accelerating splice form, or AUG as a control—for 10 consecutive days. B.g.l. was monitored again until animals reached 30 weeks of age. Animals that maintained a b.g.l. <250 mg/dL on all successive readings to the age of 30 weeks were considered protected. The results of this experiment are shown in Fig. 5a. Under this treatment regimen we observed a dramatic influence on the incidence of diabetes in mice treated with SA2 where ∼65% (11 out of 17) of the animals were protected from disease out to 30 weeks of age. Moreover, we again observed that animals treated with the SA3 exhibit an increased sensitivity to diabetes compared with controls and SA2 treatment. All animals in this group developed disease compared to 73% and 80% in the saline and AUG groups, respectively (Fig. 5A). The influence of the two forms of CTLA-4 on development insulitis was also examined in a separate study treating young (4-week-old) NOD mice and then followed by histological examination of frozen sections taken from the pancreata. Insulitis scoring showed a significant difference between treatment groups and control animals, both in the percentage of islets exhibiting no infiltration (Fig. 5B) and those exhibiting >50% infiltration or insulitis (Fig. 5C). Mice receiving the SA3 treatment were also examined for indications of other autoimmune disease, however no overt signs were observed prior to the onset of diabetes (data not shown).
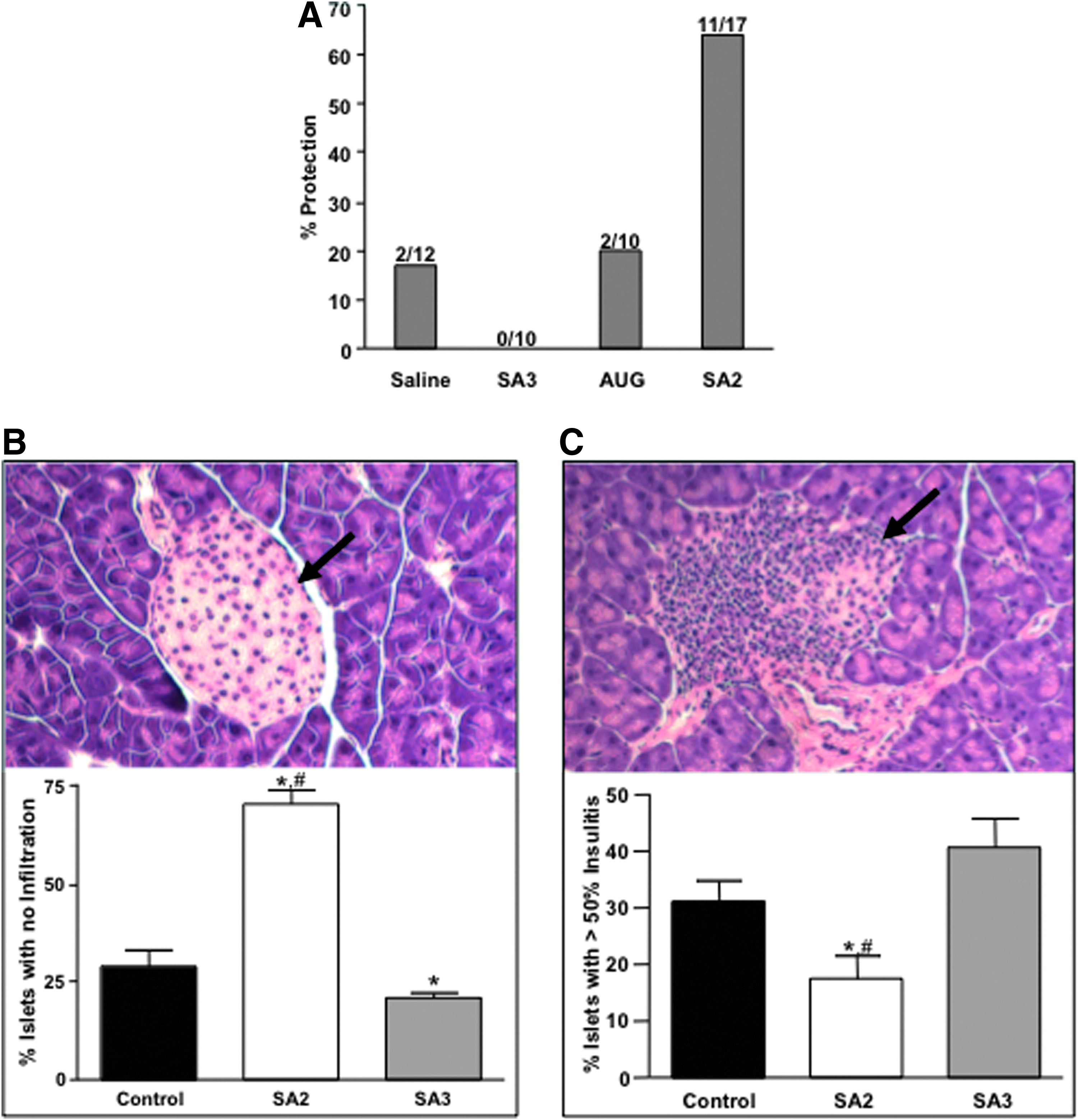
Antisense induced splice switching to produce either liCTLA-4 or sCTLA-4 protects or exacerbates diabetes onset and insulitis, respectively, in PPMO treated NOD mice.
Discussion
The role of liCTLA-4 expression in the NOD mouse model for spontaneous autoimmune diabetes has been examined in studies using transgenics (Ichinose et al., 2013; Ueda et al., 2003). Additionally, recent studies by using haplotype analysis, congenic and transgenic strains of mice on the NOD background that express levels of liCTLA-4 comparable to diabetes resistant strains of mice found that liCTLA-4 expression can alone account for protection from disease (Araki et al., 2009). Interestingly, the level of protection for inbred NOD mice strains expressing the B10 Idd 5.1 region containing the corrected SNP in exon 2 and transgenic NOD mice that express both full-length and liCTLA-4 forms together exhibit a similar level of protection (60%–65%) we observed in the NOD mice receiving the liCTLA-4 inducing SA2 PPMO. Furthermore, we have shown that NOD mice T cells induced to express liCTLA-4 by PPMO SA2 exhibit decreased proliferation and activation in addition to increased adhesion to ICAM in a similar fashion to T cells treated with α-CTLA-4, suggesting that antisense induction confirms the activity of this splice form both in vitro and in vivo.
A role in the regulation of murine T cell responses for the CTLA-4 splice forms lacking intracellular domains has only been recently examined in detail (Liu et al., 2003, 2012; Ueda et al., 2003; Ichinose et al., 2013). This is likely due to the relatively low level of expression in mice for these forms compared to those expressed in human T cells (Oaks et al., 2000; Ueda et al., 2003). However, there is some evidence to support its role in NOD type 1 diabetes observed in mice generated to inhibit the expression of sCTLA-4 and not full-length and liCTLA-4 via RNAi, where these mice exhibited reduced potency of T regulatory cells (Kissler et al., 2009). CD4+ CD25+ FoxP3+regulatory T cells (T regs) have been described in the NOD mouse to be defective in numbers and suppressive function and may play a role in the onset of autoimmune disease and CTLA-4 is critical to T reg function (Read et al., 2006). We did examine the population of CD4+ CD25+ cells expressing FoxP3 in the nodes of young NOD mice after PPMO treatment and found an increased percentage of positive cells in the SA2 treated group compared to control and SA3 (data not shown). It is, however, unlikely that the therapeutic benefit produced by the liCTLA-4 SA2 PPMO treatment was due to promoting expression in T regs, as others have shown by adoptive transfer of liCTLA-4 expressing T regs that they do not provide protection from disease onset in NOD mice. Taken together these results suggest that antisense mediated splice-switching technology can be used to examine what influence alternative splice forms have on the immune response and a therapeutic approach can be employed to restore expression of beneficial splice forms.
Although a correlation has been observed in humans between the risk for developing autoimmune disease, including type 1 diabetes, and expression of sCTLA-4 (Liu et al., 2003; Ueda et al., 2003; Simone and Saverino, 2009) as well as soluble forms of other costimulatory molecules (Wan et al., 2006; Kakoulidou et al., 2007; Nielsen et al., 2007) the direct regulatory effect remains unclear and somewhat controversial. In our study, we have observed that induction of sCTLA-4 in NOD mouse T cells results in decreased ICAM-1 adhesion activity. Moreover, treatment with antisense to promote sCTLA-4 in the NOD mouse markedly increases both the rate and frequency of diabetes. This finding is currently under study to determine the immunological consequences of over expression of sCTLA-4 in the mouse immune system. Observations made in this study and confirmed by others suggest that expression of the alternative spliced forms of CTLA-4, whether through genetic variation or manipulation, do contribute to the resistance or susceptibility to autoimmune disease.
The antisense concept is simple and is based on Watson-Crick hybridization mechanisms, (CROOKE, 1999). Antisense oligonucleotides are classically designed to inhibit the translation of RNA specifically by binding to complementary RNA target-sequence with high affinity and specificity. Typically, antisense reagents consist of oligonucleotides that have been modified from the standard phosphodiester oligonucleotides that make up “plain DNA.” Some modification is required in order to develop resistance to the nuclease-mediated degradation of DNA and RNA as it exists within the in vivo environment. Various approaches have been used to achieve nuclease resistance while retaining the requisite binding affinity of the antisense reagent to the RNA target. In addition, all modifications, while retaining antisense activity, must occur in the absence of associated toxicity in vivo. The modifications that have been employed typically involve sugar modifications at the 2′ position (e.g., methoxy substitution), base modification (e.g., 5-methyl cytosine), or backbone modifications ranging from a sulfur substitution of one of the non-bridging phosphoryl oxygens (formation of phosphorothioate antisense) to replacement of the deoxy sugar (e.g., morpholino based antisense) (Summerton and Weller, 1997).
Antisense reagents can exert their influence at the RNA level by two mechanisms. The phosphorothioate antisense molecules, or S-DNAs, and siRNAs bind to their RNA target sequence and in turn trigger pathways that degrade mRNA and thus inhibit translational efficiency. Other antisense reagents do not induce RNase H activity but function by a steric blocking mechanism. Antisense reagents that induce RNase H activity are not useful for splice switching, because of subsequent destruction of targeted pre-mRNA resulting in complete loss of protein as opposed to the needed restoration of an alternative function. For this application what is needed are antisense molecules that provide adequate steric inhibition of spliceosome recognition of targeted splice-sites while remaining inert to inducible nuclease activity and innate recognition and response systems that can alter adaptive responses and confound interpretation of splice variant function.
Mechanisms regulating alternative splicing are complicated, vary greatly between genes and involve different binding sequences for numerous ancillary splice-enhancer or -suppressor components of the spliceosome (BLACK, 2003). Antisense mediated redirection of splicing has been used to block cryptic splice sites (Vacek et al., 2003). However, targeted specific splice switching involves some trial and error to optimize antisense sequences that induce the removal of a desired exon in the larger percentage of the mRNA target such is in the case with CTLA-4 presented here or when targeting mutations in the dystrophin gene (Alter et al., 2006). Although splice acceptor targeting was the most effective strategy for inducing the expression of alternative forms of CTLA-4 and other pre-mRNAs we have targeted (Marshall et al., 2007), this is apparently not a rule that can be generalized to all target genes. For example, when targeting the dystrophin gene in animal models for muscular dystrophy, splice donor sequences were found to be more effective than targeting acceptor sequences (Wilton et al., 1999). Thus, an optimal targeting strategy to induce the desired splicing event has to be determined empirically by conducting a survey with antisense molecules surrounding intron and exon boundaries of the specific exon to be targeted to find the most potent sites. However, we have observed that targeting acceptor junction sites are predictably and potently active when targeting exons in genes that are naturally skipped to produce alternative splice forms as is the case with CTLA-4.
Aberrant splicing leading to loss or altered function of a protein is the cause of numerous human diseases (Faustino and Cooper, 2003). The precise splice-site and/or control sequence mutations that cause many of these genetically acquired diseases are often known, nevertheless, little can be done to therapeutically treat the lesions in the genome or the pathogenic effects they cause. The results presented here as well as in other disease models such as with β-thalassemia, a hereditary blood disorder and Duchenne muscular dystrophy a degenerative disease primarily affecting muscles, including the heart have employed splice-switching technology with promising outcomes (Vacek et al., 2003; Fletcher et al., 2007). Thus, this approach serves as feasible way to correct genetic abnormalities due to incorrect or under expressed splice forms and offers significant promise for continued development as clinical therapeutic.
There are studies that demonstrate the usefulness of agonists of CTLA-4 in autoimmune models and some that have been shown to be effective for the treatment of diabetes in the NOD mouse (Fife et al., 2006). However, due to the complex biology of CTLA-4 and other related co-stimulatory receptors (Salomon and Bluestone, 2001), the antibody or chimeric ligand agonist approach has lead to conflicting results, as well as exacerbating autoimmunity and type 1 diabetes (Vanderlugt et al., 1997; Luhder et al., 1998). Many of these problems could be overcome utilizing the exquisite specificity that antisense can offer for targeting expression of immune regulatory molecules. The combined use of the uncharged PMO chemistry and peptide-assisted delivery we were able to effectively deliver antisense molecules into T cells influencing in vitro and in vivo activity and demonstrating the utility of this approach in regulating immune responses for the first time.
Although further experimentation is required to determine a minimal effective dose for inhibiting or significantly delaying the onset of diabetes in the NOD mouse model beyond the proof of concept utilization of a maximal feasible, these results demonstrate that antisense can alter the splicing of immune modulating molecules. Moreover, this can be conducted in a manner to produce biologically relevant alternative splice forms of CTLA-4 such as those found to be expresses in strains of mice resistant to developing spontaneous autoimmune diabetes and thus affect T cell activity to alter the course of autoimmune responses in the NOD mouse.
Footnotes
Acknowledgments
We wish to thank the chemistry group at Sarepta Therapeutics, Inc. for synthesis, purification, and peptide conjugation of the PPMO used in this study. In addition, we would like to recognize the Oregon State University Center for Genome Research and Biocomputing Laboratories for their help with DNA sequencing and the veterinarian diagnostic labs for histological processing and analysis.
Author Disclosure Statement
DVM, FJS, SLC, PLI have received employment compensation in the form of wages and benefits from Sarepta Therapeutics, Inc. All work conducted and reported herein was funded by Sarepta Therapeutics, Inc.