
Abstract
Gene knockdowns using oligonucleotide-based approaches are useful for studying gene function in both in vitro cell culture systems and in vivo animal models. We evaluated the efficacy of 2′,4′-bridged nucleic acids (BNA)-modified antisense oligonucleotides (AONs) for gene knockdown in zebrafish. We used the tcf7l1a gene as a model for testing the knockdown efficacy of 2′,4′-BNA AONs and examined how the target sites/affinity and RNase H induction activity of 2′,4′-BNA AONs affect knockdown efficacy. We found that tcf7l1a gene function was knocked down by 2′,4′-BNA AONs that target the start codon and induce RNase H activity. Although nonspecific p53-mediated developmental defects were observed at higher doses, the effective dose of the 2′,4′-BNA AONs for tcf7l1a is much lower than that of morpholino oligonucleotides. Our data thus show a potential application for 2′,4′-BNA AONs in the downregulation of specific genes in zebrafish.
Introduction
O
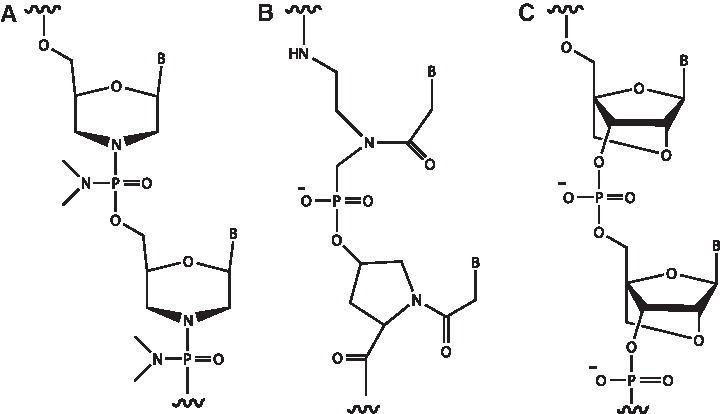
Antisense oligonucleotides.
RNase H-dependent oligonucleotides, which can induce mRNA degradation, comprise another class of antisense oligonucleotides (AONs). RNase H specifically recognizes RNA:DNA heteroduplexes composed of the A-form RNA strand and the B-form DNA and subsequently hydrolyzes the RNA component. We previously developed a 2′,4′-bridged nucleic acid (2′,4′-BNA, Fig. 1C), also called locked nucleic acid (LNA), that includes a methylene bridge between the 2′ and 4′ positions in the ribose ring (Obika et al., 1997; Obika et al., 1998; Singh et al., 1998). This modification confers resistance to nucleases. Further, 2′,4′-BNA has a high affinity for RNA. 2′,4′-BNA:RNA duplexes structurally resemble A-form RNA:RNA duplexes; thus, 2′,4′-BNA does not activate RNase H on its own (Singh et al., 1998). However, 2′,4′-BNA/DNA chimeric oligomers with a DNA stretch of at least six nucleotides can activate RNase H (Kurreck et al., 2002). Therefore, the 2′,4′-BNA/DNA chimeric oligomer can act through both steric blocking and RNase H-dependent mechanisms to efficiently knock down endogenous gene expression (Obika et al., 2001). Recently, modified BNA AONs have been developed as one of the most promising antisense oligonucleotide analogues for clinical applications (Rayburn and Zhang, 2008; Seth et al., 2009; Prakash et al., 2010; Yamamoto et al., 2011; Yamamoto et al., 2012).
Zebrafish is a useful in vivo model to understand gene function in both developmental and physiological contexts. Antisense agents, such as morpholino oligos, have been widely used for loss of gene function studies in zebrafish (SUMMERTON, 1999; Nasevicius and Ekker, 2000; Urtishak et al., 2003). However, the efficacy of 2′,4′-BNA AONs in zebrafish has not been well studied. Here, we examine whether 2′,4′-BNA/DNA chimeric oligomers are able to knock down gene function by screening various target regions and numbers and positions of 2′,4′-BNA in the oligonucleotide sequence using zebrafish tcf7l1a as a model gene. Although the dose for nonspecific toxicity is close to the effective dose, select 2′,4′-BNA AONs can knock down tcf7l1a function in zebrafish.
Materials and Methods
Fish maintenance
The zebrafish AB strain was raised and maintained under standard conditions approved by the Institutional Animal Care and Use Committee at Chiba University.
Antisense oligonucleotides
2′,4′-BNA/DNA- and 2′,4′-BNA/RNA-chimeric oligomers were synthesized by Gene Design, Inc. The RNA melting temperature (Tm) of the BNA/DNA oligomers was estimated using the Exiqon Tm prediction program (www.exiqon.com/ls/Pages/ExiqonTMPredictionTool.aspx). All morpholino oligonucleotides were purchased from Gene tools, LLC. The morpholino oligonucleotides were previously described as follows: tcf7l1a MO (Dorsky et al., 2003) and p53 MO (Robu et al., 2007).
Zebrafish embryo injections
Fertilized eggs were collected, and 0.2–1 nL of oligonucleotides was injected into one- to two-cell-stage embryos. After injection, the embryos were incubated at 28.5°C and raised until 24 hours post fertilization (hpf ). Bright-field images were taken using a Zeiss Axiocam HR mounted on a Leica stereoscopic microscope.
RNA isolation and quantitative real-time polymerase chain reaction
Embryos were injected with 2.4 fmol of 2′,4′-BNA oligos. At tail-bud stage (around 10 hpf ), 30–50 embryos were collected and total RNA was purified using standard TRIzol and propan-2-ol precipitation. Complementary DNA synthesis was performed with the ReverTra Ace. Transcript levels of tcf7l1a and Rpl13α were quantified by real-time polymerase chain reaction (PCR) with Power SYBR Mix (Applied Biosystems) on a 7300 Real-Time PCR detection system (Applied Biosystems). Results are expressed relative to the level of the housekeeping gene Rpl13α (Tang et al., 2007). Primer sequences are as follows: tcf7l1a (up), 5′-CAAGGCCAGCATATGTACTCC-3′; tcf7l1a (low), 5′- AACGGCTGGAGACCAGACTA-3′; Rpl13α (up), 5′-TCTGGAGGACTGTAAGAGGTATGC-3′; and Rpl13α (low), 5′-AGACGCACAATCTTGAGAGCAG-3′.
Results and Discussion
Injection of an antisense morpholino against tcf7l1a causes the loss of the anterior head in zebrafish
Antisense MOs have been used to knock down endogenous genes in zebrafish (Nasevicius and Ekker, 2000). To analyze the efficacy of gene knockdown by 2′,4′-BNA antisense oligonucleotides (AON), we used the zebrafish tcf7l1a gene as a case study. As previously published, tcf7l1a functions as a repressor of Wnt signaling (Kim et al., 2000). Wnt signaling plays an important role in anterior–posterior neural patterning, and exaggerated Wnt signaling leads to the loss of the rostral brain region and the expansion of the more caudal brain region (Ciani and Salinas, 2005). The headless (hdl) mutant, in which the tcf7l1a gene is inactivated and thus has excessive Wnt signaling, lacks eyes and a forebrain region but exhibits a rostral expansion of the midbrain–hindbrain boundary (Kim et al., 2000). It has been shown that injection of an antisense MO against tcf7l1a causes similar head defects to those of hdl mutants (Dorsky et al., 2003; Thorpe and Moon, 2004). To confirm this finding, we injected 240 or 600 fmol of the tcf7l1a MO into fertilized zebrafish eggs. Injected embryos showed head defects at 24 hpf (70%–90% of the embryos had no eyes, Fig. 2A, B). The morphological head defect phenotype was enhanced by injection of a higher dose of the tcf7l1a MO, but this treatment also caused additional neuronal death, possibly due to off-target effects of the MO (Robu et al., 2007).
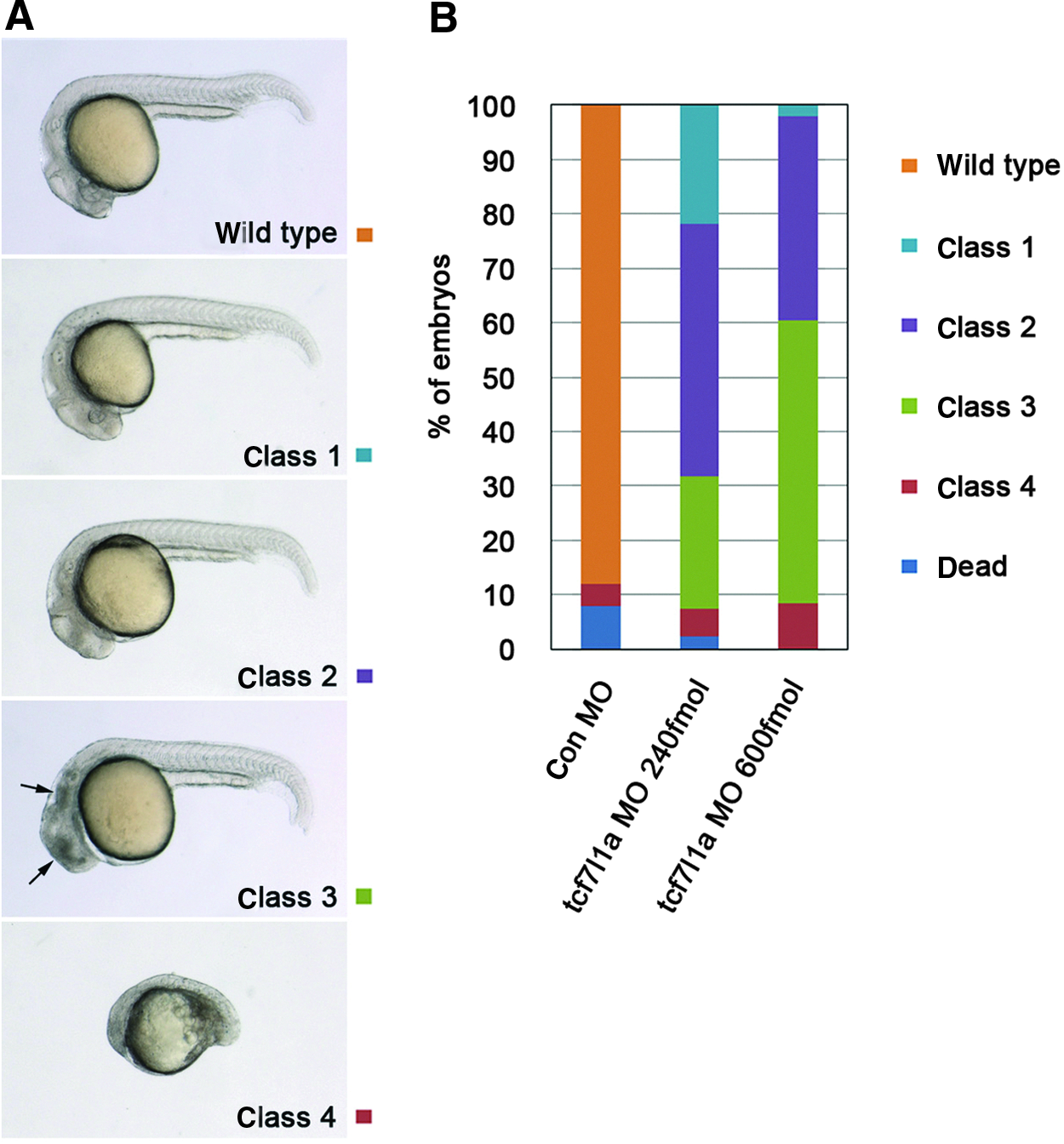
Representative phenotypes obtained after injection of control or tcf7l1a MO. At 24 hours post fertilization (hpf ), phenotypes were assessed by morphological criteria, according to severity.
2′,4′-BNA/DNA chimeric oligomers that activate RNase H against tcf7l1a affect anterior head formation
The tcf7l1a MO is a 25-bp sequence of oligonucleotides that targets a region that includes the start codon (Region 1, Fig. 3). We first chose the same region (Region 1 in Fig. 3, Table 1) to study the effects of 2′,4′-BNA in zebrafish embryos. MOs function solely by a steric block mechanism to inhibit translation and RNA splicing (SUMMERTON, 1999; SUMMERTON, 2007). In contrast, 2′,4′-BNA/DNA-mixed AONs either exploit RNase H to cleave their target RNAs and/or interfere sterically with RNA to block its function. Previous studies have shown that a DNA stretch of six nucleotides in the 2′,4′-BNA/DNA chimeric oligomer is required to activate RNase H (Kurreck et al., 2002; Elmen et al., 2004). We therefore designed various 2′,4′-BNA AONs that would either activate RNase H or not (Table 1). The injection of 60 fmol of tcf7l1a 2′,4′-BNA/DNA chimeric oligomers (tcf7l1a-BNA1 through tcf7l1a-BNA5) resulted in a lethal phenotype (i.e., no injected embryos survived to 24 hpf ). When the injected dose of 2′,4′-BNA/DNA chimeric oligomers was reduced to 2.4 fmol per embryo, more than half of the embryos survived. Among the tcf7l1a-BNA1-5 AONs, tcf7l1a-BNA2 yielded the highest proportion of head defects (51% of embryos had no eyes; 34% of embryos had small eyes; n=87, Fig. 4A, B). However, 8% of the embryos exhibited short body axes and cell death (class 3 in Fig. 4A, B). These abnormalities were not observed in hdl mutant embryos, so they are likely due to off-target effects of tcf7l1a-BNA2. As for the other AONs, tcf7l1a-BNA4 caused head defects but also developmental abnormalities and embryonic lethality (9% with no eyes; 14% with small eyes; 7% with short body axes; 22% malformed; 44% dead; n=100); the others, including tcf7l1a-BNA3, had no or very little effect (77% normal; 6% with small eyes; n=48, Fig. 4A, B, data not shown for tcf7l1a-BNA1 and tcf7l1a-BNA5). To evaluate the RNase H activity of 2′,4′-BNA/DNA chimeric oligomers in vivo, tcf7l1a expression was examined in injected embryos by quantitative real-time PCR. tcf7l1a-BNA2 and tcf7l1a-BNA4 reduced tcf7l1a expression but tcf7l1a-BNA3 did not have a significant effect (Fig. 4C).
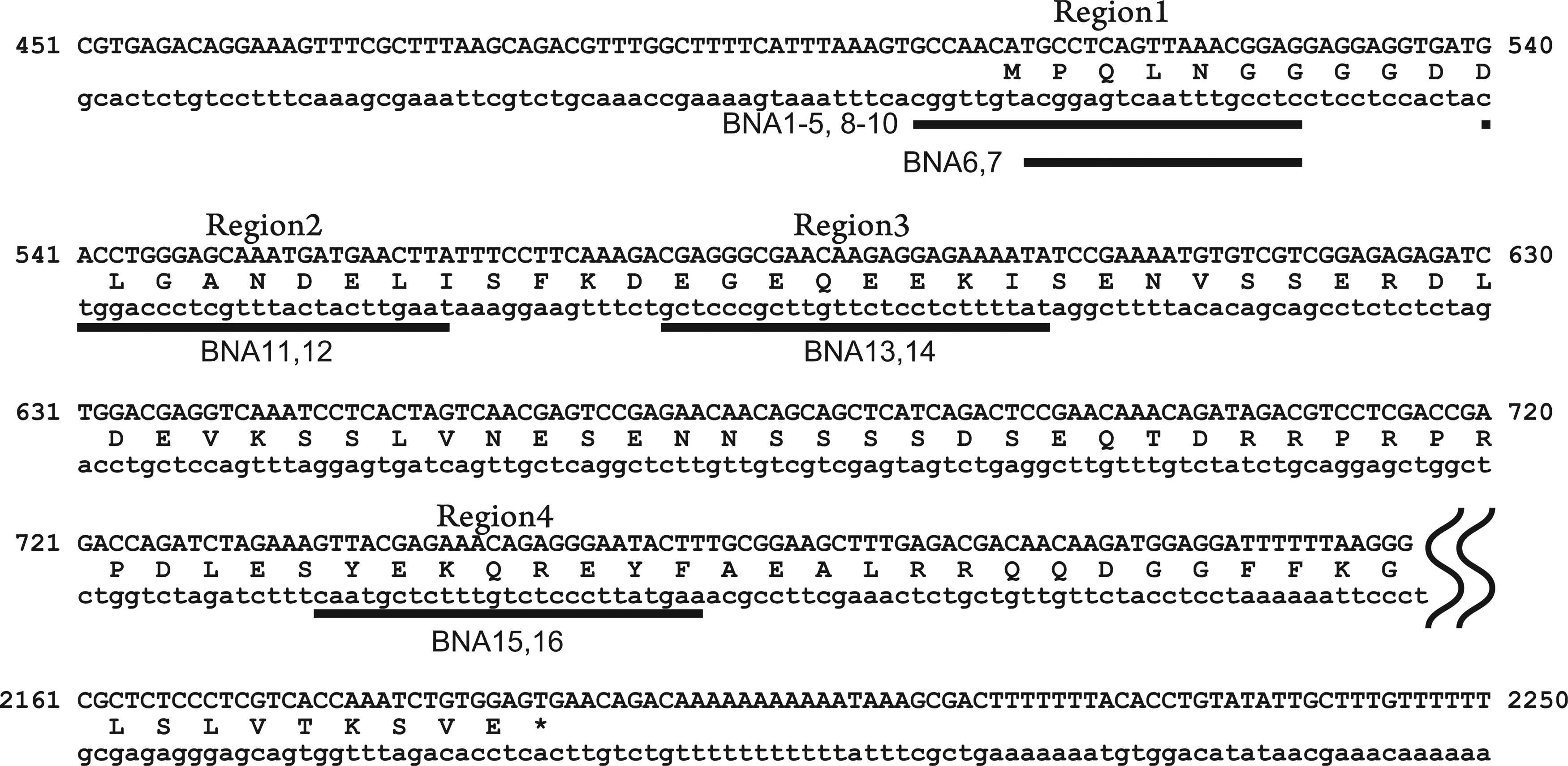
Target positions of 2′,4′-BNA/DNA chimeric oligomers in tcf7l1a. Uppercase and lowercase letters in the sequences represent the sense and antisense strands, respectively. The numbers at the left and right refer to the nucleotide positions in the tcf7l1a gene target sequences of the 2′,4′-BNA/DNA AONs are underlined.
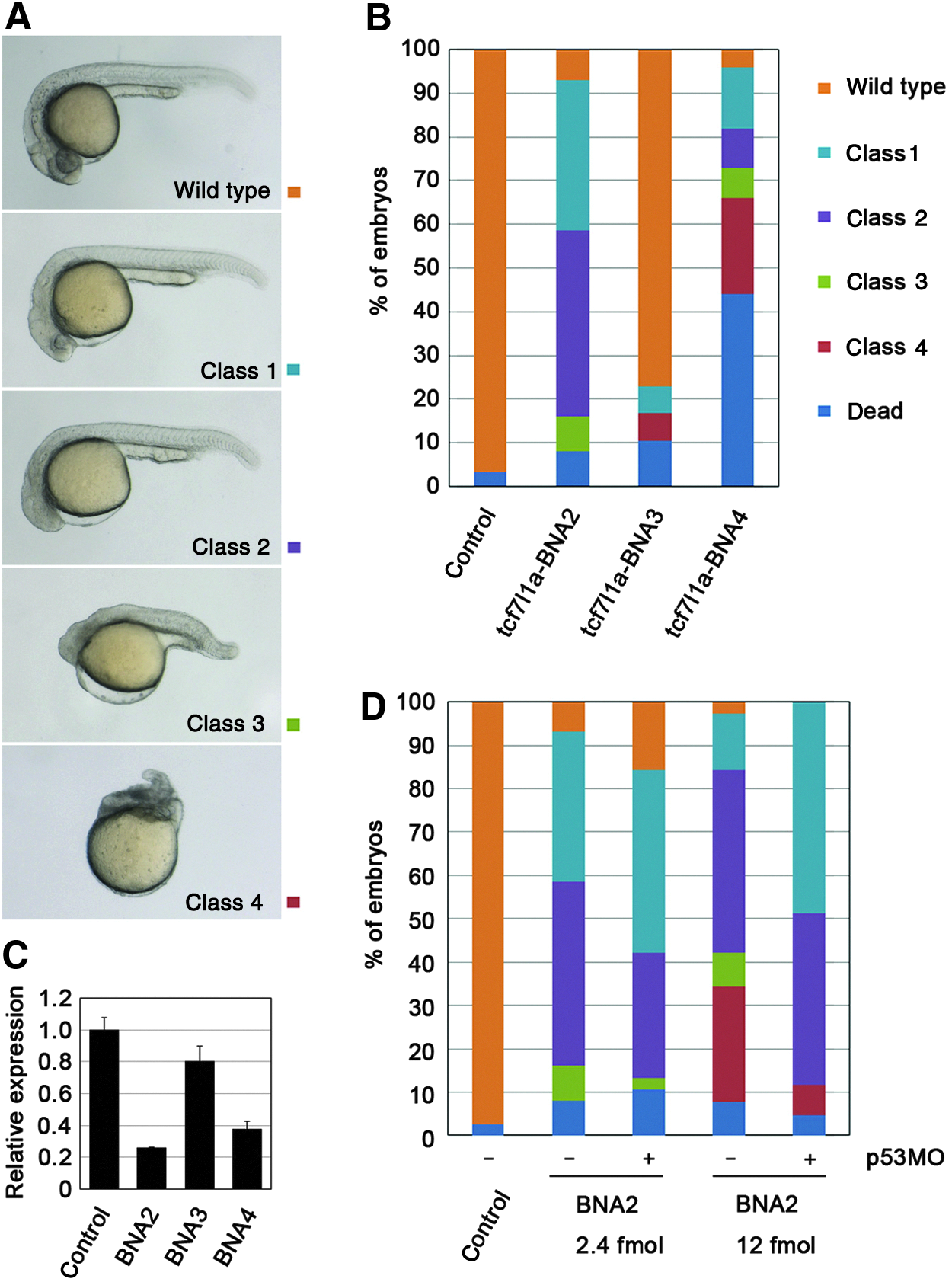
2′,4′-BNA/DNA chimeric oligomers that activate RNase H against tcf7l1a affect anterior head formation, and off-target effects are mediated by p53 activation.
Upper- and lowercase letters in the sequences represent 2′,4′-bridged nucleic acids (BNA) and DNA, respectively.
Lowercase and underlined letters in the sequence represent RNA.
T, G, C: 2′,4′-BNA-T, G, 5-methy-C.
To validate the effects of the 2′,4′-BNA modification, we injected tcf7l1a-DNA, which is composed of DNA only but has the same sequence as tcf7l1a-BNA2. Sixty-four percent of the embryos injected with 10 fmol of tcf7l1a-DNA survived with no brain defects (n=41, data not shown). Increasing the dose to 50 fmol per embryo did not elicit the aforementioned head defects but did reduce the survival rate (17% survival, n=44). Next, all of the DNA nucleotides within the tcf7l1a-BNA2 sequence were replaced with RNA nucleotides, so that RNase H could not be activated (designated tcf7l1a-BNA/RNA2). The survival rate at higher dosages was improved by this modification, but the survivors showed a weak phenotype (at 60 fmol injection per embryo, 93% survived; 0% had no eyes; 41% had small eyes; n=41). Increasing the dose of tcf7l1a-BNA/RNA2 (600 fmol per embryo) slightly enhanced the gene knockdown effects but reduced the survival rate (0% with no eyes; 100% with small eyes; 11% survival; n=28). Therefore, these results suggest that 2′,4′-BNA/DNA chimeric oligomers with RNase H induction activity have a higher ability to inhibit endogenous tcf7l1a gene function.
We next investigated the potency of the shorter 2′,4′-BNA/DNA AONs, tcf7l1a-BNA6 and 7. The AON tcf7l1a-BNA6, which was expected to have a similar RNase H induction activity to that of tcf7l1a-BNA2, did not cause head defects, nor did tcf7l1a-BNA7. Thus, to ensure high knockdown efficacy in zebrafish, 2′,4′-BNA/DNA chimeric oligomers should have optimal binding affinity to the target sequence. We next examined the effects of 2′,4′-BNA/DNA chimeric oligomers with longer consecutive DNA nucleotides (tcf7l1a-BNA8-10, Table 1). Kurreck et al. reported that 2′,4′-BNA/DNA chimeric oligomers with a stretch of six DNA bases stimulated RNase H, while those with longer DNA stretches activated RNase H more effectively (Kurreck et al., 2002). Upon injection of tcf7l1a-BNA8-10 at 10 fmol per embryo, only tcf7l1a-BNA10 showed very weak activity (4% of embryos had small eyes; 92% survival, n=25), while the others elicited no head defects at all (tcf7l1a-BNA8: 23% survival, n=23; tcf7l1a-BNA9: 65% survival, n=23). Because the Tms of tcf7l1a-BNA8-10 are not as high as those of tcf7l1a-BNA2 and 4, and because higher RNase H activity cannot compensate for the lower Tms, adequate Tm and RNase H activity are required for effective 2′,4′-BNA/DNA chimeric oligomer function.
Target-dependent action of 2′,4′-BNA/DNA chimeric oligomers
We next attempted to identify effective 2′,4′-BNA/DNA chimeric oligomers for tcf7l1a in other regions of the tcf7l1a gene. Six 2′,4′-BNA/DNA chimeric oligomers targeting three regions downstream of the start codon were examined (tcf7l1a-BNA11-16, Fig. 3). Because their target regions are at a distance from the ATG codon, they cannot function to inhibit translation. As a result, none of the tcf7l1a-BNA11-16 AONs showed any head defects at doses of 3 or 10 fmol per embryo (data not shown). Taken together, both protein translation blocking and RNase H-mediated mRNA degradation may play important roles for effective gene knockdown by 2′,4′-BNA/DNA chimeric oligomers in zebrafish.
Nonspecific toxicity of 2′,4′-BNA/DNA chimeric oligomers in zebrafish embryos is mediated by p53 activation
We found that tcf7l1a-BNA2 was highly efficient at blocking tcf7l1a function but also that some embryos showed nonspecific abnormalities and cell death, which were also observed with the other 2′,4′-BNA/DNA chimeric oligomers. Some morpholino AONs and small interfering RNAs have been reported to induce p53-mediated apoptosis nonspecifically in zebrafish and in cultured cells (Scacheri et al. 2004; Robu et al. 2007). Therefore, we examined whether 2′,4′-BNA/DNA AON-induced off-target toxicity is mediated by p53. Embryos were initially injected with a control MO or p53 MO, and then tcf7l1a-BNA2 was injected at 2.4 or 12 fmol per embryo. The percentage of embryos with short body axes was reduced by p53 knockdown at both dose levels (class 3, Fig. 4D). Furthermore, when coinjected with 12 fmol of tcf7l1a-BNA2, the p53 MO reduced the percentage of malformed embryos (class 4) and reciprocally increased the percentage of embryos with small eyes (class 1, Fig. 4D). Thus, p53 knockdown significantly alleviated the off-target developmental abnormalities induced by 2′,4′-BNA/DNA AONs in zebrafish. These data suggest that p53 activation is a major cause of toxicity by 2′,4′-BNA/DNA AONs in zebrafish.
Conclusions
In conclusion, we have shown that 2′,4′-BNA/DNA chimeric oligomers are able to knock down gene function in zebrafish embryos. One advantage of 2′,4′-BNA/DNA chimeric oligomers is that the effective dose is much lower (∼one-hundredth) than that of morpholino oligos. However, the effective dose is close to the toxic dose that triggers nonspecific developmental defects. Thus, the fine adjustment of target sequences, the number and position of the 2′,4′-BNA, and the RNase H induction activity are required to effectively achieve knockdown using 2′,4′-BNA/DNA AONs. Future studies are required to further improve the effectiveness of 2′,4′-BNA/DNA AONs in zebrafish.
Footnotes
Acknowledgments
We thank the Zebrafish National BioResource Project in Japan for providing the zebrafish strains and H. Matsuo and R. Inaba for their technical assistance. This research was supported by the Daiichi Sankyo Foundation of Life Science and the Sumitomo Foundation (MI) as well as by Grants-in-Aid for Scientific Research programs in Japan (MI, TI, SO).
Author Disclosure Statement
No competing financial interests exist.