
Abstract
microRNAs (miRs) regulate vascular diseases such as atherosclerosis and cancer. miR-126 is important for endothelial cell signaling and promotes angiogenesis, protects against atherosclerosis, and reduces breast cancer cell growth and metastasis. The overexpression of miR-126, therefore, may be an attractive therapeutic strategy for the treatment of cardiovascular disease or cancer. Here we report a novel strategy to deliver miR-126 to endothelial and breast cancer cells. We tested three different strategies to deliver miR-126 by linking the miR to an aptamer for the ubiquitously expressed transferrin receptor (transferrin receptor aptamer, TRA). Linking the precursor of miR-126 (pre-miR-126) to the TRA by annealing of a complementary stick led to efficient uptake and processing of miR-126, resulting in the delivery of 1.6×106±0.3×106 copies miR-126-3p per ng RNA in human endothelial cells and 7.4×105±2×105 copies miR-126-3p per ng in MCF7 breast cancer cells. The functionality of the active TRA-miR-126 chimera was further demonstrated by showing that the chimera represses the known miR-126 target VCAM-1 and improved endothelial cell sprouting in a spheroid assay. Moreover, the TRA-miR-126 chimera reduced proliferation and paracrine endothelial cell recruitment of breast cancer cells to a similar extent as miR-126-3p mimics introduced by conventional liposome-based transfection. Together, this data demonstrates that pre-miR-126 can be delivered by a non-specific aptamer to exert biological functions in two different cell models. The use of the TRA-miR-126 chimera or the combination of the delivery strategy with other endothelial or tumor specific aptamers may provide an interesting therapeutic option to treat vascular disease or cancers.
Introduction
M
Due to their multiple roles in physiological and disease processes, miRs have come into the focus of research in recent years. In the cardiovascular system, various miRs were described to modulate atherosclerosis and regulate the growth of new blood vessels during embryonic development as well as during vascular diseases [6–8]. Particularly, miR-126 was shown to be essential for vessel growth in zebrafish and its inhibition reduced neovascularization in mice [9–11]. Unlike other miRs described in the literature, both the 3p and the 5p forms of miR-126 have been described to fulfill a biological function. Since earlier publications have used a complete deletion or viral delivery of both forms, it is not always clear which of the two forms is responsible for the observed phenotypes. MiR-126-3p reduces vascular inflammation by targeting the endothelial adhesion molecule vascular cell adhesion molecule (VCAM-1) [12], and both miR-126-3p and miR-126-5p inhibit atherosclerotic lesion progression [13,14].
In addition to the beneficial effects of miR-126 in the vasculature, miR-126 has been directly and indirectly implicated in the progression of various types of cancer [15,16]. In breast cancer, miR-126 is often downregulated, and restoring its expression reduces tumor growth, tumor angiogenesis, and metastasis [17–19]. Therefore, overexpression of miR-126 might be a promising therapeutic approach to improve vascular function and reduce tumor growth and metastasis. Whereas inhibition of miRs can be easily achieved (e.g., by cholesterol-modified antagomirs or locked nucleic acid–based antimiRs) in large animal models and first clinical trials [20,21], the delivery of pre-miRs is less efficient and requires carriers (e.g., liposomes) [22]. Since miRs are known to exert different functions in individual cell types, a cell-specific delivery may provide an advantage and prevent unwanted activities of a particular microRNA therapeutic.
In recent years aptamers have emerged as tools to specifically transport molecules into cells [23,24]. Aptamers are small single stranded oligonucleotides possessing a specific three dimensional structure [25]. This allows aptamers to interact with target molecules, in a manner similar to antibodies. RNA aptamers have been developed against a range of targets, for example against vascular endothelial growth factor (VEGF) to treat age related macular degeneration [26–28]. Some aptamers bound to surface receptors can be internalized into cells, allowing their usage as agents for drug delivery [29]. During the last couple of years, several research groups have used aptamers to transport small interfering RNAs into cells [30–32] by using e.g. the general RNA internalization motif (GIRM) [33] or an aptamer for the ubiquitously expressed transferrin receptor (TRA) [34]. Recently, two groups reported that aptamers can also be used to deliver miRs into cells. A MUC1 aptamer, which preferentially binds to tumor expressed mucin 1, was linked to miR-29b and inhibited the growth of xenograft tumors [35,36]. Let-7g miR was conjugated to the aptamer GL21.T that binds to and antagonizes the oncogenic receptor tyrosine kinase Axl, and was efficiently delivered to target cells, leading to the suppression of let-7 target genes and reduction of tumor growth in a xenograft model of lung adenocarcinoma [37]. Moreover, polymers conjugated to the TD05 immunoglobulin heavy mu chain aptamer [38] were used to deliver miR-15a and miR-16 to prostate cancer cells [39].
Here, we tested three different strategies for delivering miRs to endothelial cells by linking the 22 nt mature duplex or pre-miR-126 to TRA. We showed that all three strategies resulted in delivery of TRA and miR-126 to endothelial cells, however only the pre-miR-126 linked aptamer led to generation of functionally active miR-126. Aptamer delivered pre-miR-126 reduced its target VCAM-1 and increased angiogenesis in endothelial cells. In tumor cells, TRA-linked pre-miR-126 increased levels of miR-126-3p and miR-126-5p, which affected proliferation and inhibited the recruitment of endothelial cells.
Material and Methods
Cell culture
Human umbilical cord-derived venous endothelial cells (HUVEC) were purchased from Lonza and cultured in endothelial cell basal medium (EBM) (Lonza, Basel, Switzerland) supplemented with 10% FBS and EGM-SingleQuots (Lonza). Murine endothelial cells (MEC) were provided by E. Dejana (Department of Biomolecular Sciences and Biotechnologies, School of Sciences, University of Milan, Italy) and cultured as previously described [40]. Murine embryonic fibroblasts were grown in Dulbecco's modified Eagle's medium (DMEM) (Sigma-Aldrich, St. Louis, MO) supplemented with 20% M-199, 15% fetal calf serum (FCS), and 1% penicillin and streptomycin (Roche, Mannheim, Germany). MCF7 and SK-BR3 breast cancer lines were kindly provided by Prof. Dr. A. Rieger (LOEWE Center for Cell and Gene Therapy, Goethe-University, Frankfurt am Main, Germany) and cultured in DMEM (high glucose, no glutamine; Gibco, Carlsbad, CA) supplemented with 10% FCS, 1% insulin-transferrin-selenium solution in Earle's balanced salt solution (Gibco) and 1% penicillin and streptomycin (Roche).
Aptamer generation
The 2′-F (2′-deoxy-2′-fluoro)-modified RNA aptamers and chimera constructs were purchased from Dharmacon (Lafayette, CO; sequences, see Table 1). Sequences were designed not to interfere with the folding of the secondary structures of the aptamers and pre-miR as predicted by RNAstructure [41] (Supplementary Fig. S1A–C; Supplementary Data are available online at www.liebertpub.com/nat). The control aptamer construct was designed as a scrambled version of TRA and ordered as single stranded DNA from Sigma-Aldrich. The DNA was amplified using standard polymerase chain reaction (PCR) reagents (Sigma) then run on a 3% agarose gel. Bands of appropriate size were excised and purified using the QiaQuick gel extraction kit (Qiagen, Hilden, Germany). The purified DNA was then transcribed in vitro with 2′-F modified uridine and cytosine using the T7 Durascribe kit (Epicentre, Madison, WI). Before use, aptamers and chimeras were folded and then annealed by separately heating all RNA fragments at a concentration of 5 μM in Dulbecco's phosphate-buffered saline (DPBS) containing magnesium chloride and calcium chloride (Sigma) at 70°C for 3 minutes, followed by 10-minute incubation at room temperature and subsequent incubation at 37°C for 10 minutes to allow folding. The individual components of the chimeras were then mixed and annealed at 37°C for 10 minutes.
Bold indicates nucleotides with 2′-F modified sugars, italics indicate sequences of the stick structure, small letters indicate nucleotides of the linkers, and mature miR sequences are underlined. Fi, 5′-fluorescein label (for microscopy); idT, inverted dT sequence at the 3′ end; S-S, 5′-disulfide thiol-modifier.
Cell treatment
After blocking cells with 0.1 mg/mL yeast tRNA (Sigma) for 30 minutes to inhibit unspecific binding, cells (250,000 per well in 12-well plates) were treated with aptamers at concentrations from 50 to 500 nM in 200 μL medium for 1 hour. Then cells were either directly used for detecting early aptamer uptake at 1 hour or 2 mL medium was added for prolonged incubation.
Aptamer visualization
Fluorescin-conjugated GIRM was purchased from Dharmacon whereas the TRA aptamer was labelled with Cy3 using the LabelIT™ siRNA Tracker Kit (Mirus, Madison, WI). Cells were grown to confluence on 8-chamber glass slides and then treated with 150 μL 500 nM aptamer as described above and fixed 1 hour after treatment using 4% formaldehyde in PBS. Nuclei were stained with 4′,6′-diamidino-2-phenylindole (DAPI) and cells were mounted with coverslips. Images were taken using an AxioObserver microscope (Zeiss, Oberkochen, Germany).
microRNA overexpression
Cells were transfected with 10 nM pre-miR™ microRNA mimics (Ambion/Life Technologies, Carlsbad, CA) using Lipofectamine RNAiMAX (Life Technologies) in OptiMEM (Gibco) according to the manufacturer's protocol.
RNA isolation and analysis
Cellular RNA was isolated by adding Trizol (Qiagen) followed by extraction with the miRNAeasy kit (Qiagen) according to the manufacturer's protocol. To isolate only internal RNA, cells were detached from culture dishes using trypsin. It should be noted that trypsin also digests most membrane bound proteins and releases any RNA bound to these proteins in the process. Then, cells were suspended in 100 μL PBS and subjected to digestion with 10 units of Riboshredder RNase cocktail (EpiBio, Madison, WI) for 15 minutes at 37°C (Gibco) prior to addition of trizol. Aptamer levels were analyzed using custom-made primers (sequences see Supplementary Table S1) and SYBR green reagents (Life Technologies), both on a StepOne cycler (Applied Biosystems, Carlsbad CA). Copy numbers were calculated based on a standard curve using recombinant miR-126-3p.
Western blot
Forty-eight hours after miR-126 overexpression or treatment with the chimera, cells were lysed in ice-cold RIPA buffer (Sigma) containing protease inhibitor cocktail (Roche). Thirty micrograms of protein per sample were run on a 10% polyacrylamide gel (AppliChem, Darmstadt, Germany), blotted to a methycellulose membrane, and stained with primary antibodies for VCAM-1 (Abcam, Cambridge, UK) or tubulin (Neomarkers, Fremont, CA) overnight. After washing, secondary anti-rabbit or anti-mouse-antibodies (GE Healthcare, Buckinghamshire, UK) were applied for 2 hours. The blots were then soaked in Immobilon Western Substrate (Millipore, Billerica, MA) and imaged using a Fluorochem analyzer (Biozym, Oldendorf, Germany). Relative amounts were calculated using Fiji software [42].
Spheroid assay
HUVEC were transfected with miR-126 or control RNA or treated with aptamers/chimeras as described above. After 24 hours, HUVEC were harvested and 400 cells per well were seeded in 96-well plates in endothelial cell basal medium (Gibco) containing methylcellulose (Sigma) for 24 hours. Resulting spheroids were harvested and divided in two wells of a 24-well plate and seeded in a mixture of collagen solution and medium containing methylcellulose. Half of the spheroids were left under basal condition, and half were treated with 50 ng/mL VEGF-A (Peprotech, Rocky Hill, CO). After 24 hours, cells were fixed in 4% formaldehyde in DPBS and images were taken using an AxioObserver microscope (Zeiss, Jena, Germany). Sprout length was measured manually using Fiji software [42].
Cck8 proliferation assay
MCF7 and SK-BR3 breast cancer cells were seeded as triplicates in a 96-well plate, incubated overnight, and treated with miR-126 or the chimeras as described above the next day. Seventy-two hours after the treatment, medium was replaced and medium containing cck8 assay reagent (Dojindo, Kumamoto, Japan) was incubated according to the manufacturer's protocol for 3 hours. Then, the absorption was measured at a wavelength of 450 nm using a Synergy HT plate reader (BioTek, Winooski, VT).
HUVEC recruitment assay
Five hundred thousand MCF7 breast cancer cells were seeded in a 24-well plate and incubated overnight before treatment with TRA or chimeras for 72 hours. To eliminate the effects of serum in the medium on recruitment, medium was replaced by DMEM containing 0.5% bovine serum albumin 16 hours prior to the analysis. Next, Boyden chambers with transwell membranes (pore size of 8 μm; Millipore) containing 50,000 HUVEC in EBM without FCS or aliquots were placed on top of the breast cancer cells and were co-cultured for 5 hours. The HUVEC on the transwell membranes were fixed in 4% formaldehyde, and then the HUVEC adherent to the upper part of the membrane were scratched off using cotton swabs. The HUVEC on the bottom were stained with DAPI, pictures were taken using an AxioObserver microscope (Zeiss), and the number of migrated cells were automatically counted using Fiji software [42].
Statistics
Statistical analysis was done using Prism 5 software (GraphPad, La Jolla, CA) using Student's t-test. All bar graphs are represented as the means±the standard error of the mean.
Results
Aptamer uptake in endothelial cells
First, we determined the efficiency of uptake of the two well characterized TRA and GIRM aptamers in HUVEC and MEC compared with fibroblasts. Since we wanted to discern between aptamers bound to the cell surface and those taken up by the cells, total and intracellular RNA was isolated after 1 hour and 24 hours of incubation with the aptamers, and TRA and GIRM levels were detected by PCR in different cell types (Fig. 1A, B). TRA was detected in total RNA isolated from all three cell types after incubation for 1 hour or 24 hours (Fig. 1A). After 1 hour of incubation, intracellular TRA was similar in all three cell types, whereas after 24 hours, internalization was optimal (about 50%) in MECs (Fig. 1A, right panel). Incubation with the GIRM aptamer showed similar results (Fig. 1B, right panel). Although GIRM levels in total RNA tended to be higher in HUVEC, the internalization ratio was best in MECs (Fig. 1B). The uptake of the aptamers was further confirmed using fluorescently labelled TRA and GIRM. Intracellular staining was detected in both TRA- and GIRM-treated MEC and fibroblasts, but not in cells treated with the control aptamer (CA), which is a scrambled version of the TRA (Fig. 1C), suggesting that both aptamers can be taken up and can be used to target endothelial cells. Since both aptamers were taken up efficiently, we chose to use TRA for the subsequent experiments since the TRA target—the transferrin receptor—is well characterized and expressed in human and mouse endothelial cells as well as tumor cells (data not shown).

Aptamer internalization in different cell types. The leftmost panel shows schematic illustrations of the secondary structure of the transferrin recepter aptamer (TRA)
Design of the aptamer-microRNA chimeras
Next we designed three chimeras linking TRA to miR-126. In the first chimera (Ch1), the mature miR-126-5p strand was covalently linked to TRA and the active miR-126-3p was annealed (Fig. 2A; Supplementary Fig. S1A). For the second chimera (Ch2), the TRA was synthesized attached to a stick structure [43], which is complementary to a stick sequence linked to the mature miR-126-5p (Fig. 2B; Supplementary Fig. S1B). Equimolar concentrations of the TRA-stick and stick-miR-126-5p sequence were then annealed, followed by the annealing of miR-126-3p with the miR-126-5p sequence (Fig. 2B). The third chimera consists of a similar covalently linked aptamer stick sequence, which was bound to a complementary stick sequence covalently linked to the pre-miR-126 (Fig. 2C; Supplementary Fig. S1C). All chimeras contain 2′-F modified sugars on all cytidine and uridine nucleotides to increase the stability of the construct. Only miR-126-3p in chimera 1 and 2 was not modified with the hypothesis that this may allow a better incorporation into the RISC complex. All three chimeras were stable for up to 24 hours, as shown by the detection of the aptamer and miR-126-3p and miR-126-5p levels (Supplementary Fig. S1D–F).
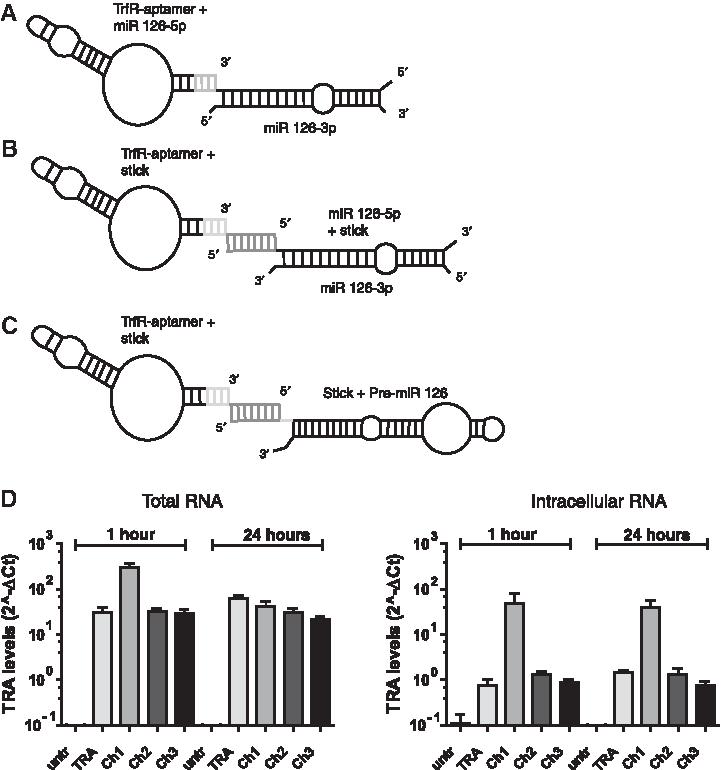
Internalization and miR-126 delivery by aptamer-miRNA chimeras
After incubation of HUVEC with 50 to 500 nM of the different chimeras for 1 hour and 24 hours, the expression of TRA was detected by quantitative PCR. TRA was detected at similar levels in TRA aptamer and Ch2- and Ch3-treated HUVEC, whereas higher levels of intracellular TRA were observed when cells were incubated with Ch1 (Fig. 2D). Next, we determined whether mature miR-126-3p and miR-126-5p are delivered into chimera-treated cells. Indeed, miR-126-3p was significantly increased in HUVEC treated with all three chimeras at 24 hours (Fig. 3A), but the highest levels of intracellular miR-126-3p were detected in Ch2-treated cells (Fig. 3B, right panel). miR-126-5p levels also were most efficiently increased by Ch2 treatment in total and intracellular RNA (Fig. 3B). Mouse endothelial cells showed a slightly different response. Whereas TRA was detected under all conditions (Supplementary Fig. S2A, B), only a modest increase of miR-125-3p levels was detected in chimera-treated cells (Supplementary Fig. S2C).

Chimeras increase levels of miR-126.
Biological functions of the chimera
After demonstrating that all chimera constructs are internalized and can increase mature miR-126 levels in HUVEC, we performed further studies to test the functionality of the chimera. Since miR-126-3p is required for angiogenesis and its overexpression increases angiogenesis (Fig. 4A, B), we determined the effect of the chimeras on endothelial sprouting using spheroid assays. Ch3 significantly increased VEGF-stimulated angiogenesis in a spheroid sprouting assay compared with TRA alone, whereas Ch1 and Ch2 did not affect angiogenesis (Fig. 4A, B). In addition, Ch3, but not Ch1 and Ch2, significantly reduced tumor necrosis factor alpha–stimulated expression of the miR-126 target VCAM-1 to a similar extent as miR-126-3p mimic transfection (Fig. 4C, D). Thus, only Ch3 showed a biological efficacy in both of these assays.
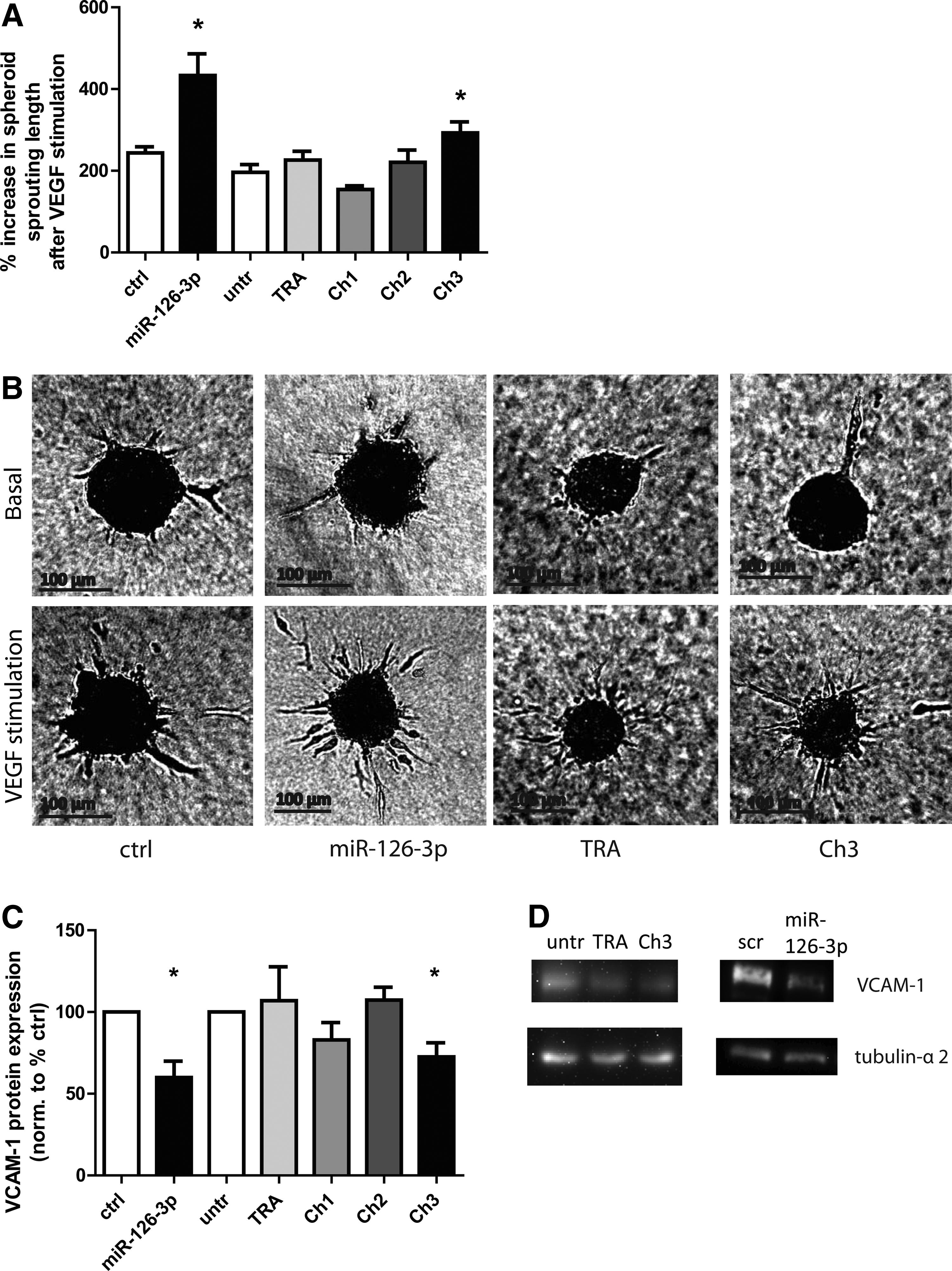
Effects of chimeras on angiogenesis and target repression in HUVEC.
Since miR-126 additionally affects tumor cell proliferation and metastasis, we determined whether Ch3 might also be used to deliver miR-126 to breast cancer cells to inhibit tumor growth. miR-126-3p expression was significantly and profoundly augmented in different breast cancer cell lines (Supplementary Fig. S3A). Since the baseline expression of miR-126-3p is rather low in MCF7 and SK-BR3 cells, we analyzed the increase in copy numbers of miR-126-3p in these cells and compared it to endothelial cells. Ch3 treatment resulted in the generation of 1.6×105±0.35×105 copies in SK-BR3 cells and 7.4×105±2×105 intracellular copies of the mature miR-126-3p per ng RNA in MCF7 cells (Fig. 5A). Ch3 treatment also led to a significant increase of miR-126-5p expression in the breast cancer cell lines (Fig. 5B).

miR-126 delivery to breast cancer cells by Chimera 3 affects proliferation and endothelial cell recruitment.
Since miR-126 was reported to inhibit tumor cell proliferation and additionally interferes with the recruitment of endothelial cells by interfering with the paracrine activity of tumor cells [18], we determined the effects of Ch3 on two different breast cancer cell lines. As shown in Figure 5, we observed striking differences between the two cell lines. As expected, Ch3 significantly repressed the viability measured in SK-BR3 cells (64.5±7.7%), as did liposomal transfection with miR-126-3p (74±3.9%) (Fig. 5C, left panel). However, Ch3 did not inhibit and even increased the viability of MCF7 cells (150.1±9.7%) (Fig. 5C, right panel). Similar effects on viability were observed when miR-126-5p was overexpressed in MCF7 cells (152.1±28.5%) or when cells were treated with TRA (122.4±9.4%) (Fig. 5C, right panel). Of note, miR-126-3p did not affect the viability of MCF7 cells. Together, these data demonstrate that Ch3 inhibits SK-BR3 proliferation, but increases viability of MCF7 cells. However, migration of HUVEC toward MCF7 cells was significantly decreased after treatment with miR-126-3p (75.5±8.3%) and Ch3 (80.5±7.5%), but no effect was observed in SK-BK3 cells (Fig. 5D, E). The biological properties of Ch3 mirrored the effects seen after overexpression of miR-126-3p (Fig. 5C–E), suggesting that the differences between the cell types is most likely due to a different biological response to miR-126. Of note, overexpression of miR-126-3p as well as Ch3 did not affect the expression of the miR-126-3p target gene Spred-1 in MCF7 cells but reduced Spred-1 levels in SK-BR3 cells (Supplementary Fig. S3B), suggesting that a different modulation of target genes underlies the different responses of the cell types.
The design of Ch3 can be used to deliver other miRNAs
The noncovalent binding of the pre-miR to the aptamer might provide the advantage to enable its use for the design of other miR-chimeras. To test whether the design of Ch3 can be used to deliver other pre-miRs, we linked the TRA to pre-miR-1 via the two complementary sticks (Ch4). In humans and mice, there are two precursors for miR-1, but both encode the same mature 3p and 5p forms; the better-described pre-miR-1a-2 was used for the chimera. For Ch4, we again used 2′-F modifications of the sugars of cytidine and uridine nucleotides, but this time we did not modify the nucleotides located within the mature miR sequences (Supplementary Fig. S4A, B). Twenty-four hours after incubation of the TRA-miR-1 chimera (Ch4), intracellular miR-1 levels were measured in murine fibroblasts. Treatment with Ch4 significantly increased the intracellular levels of mature miR-1a-2-3p (Supplementary Fig. S4C), demonstrating that the aptamer design can generally be used to deliver pre-miRs and that the constructs are stable in medium containing serum although the mature miR sequences were not modified.
Discussion
This study demonstrates that aptamers can be used to deliver functionally active miRNAs such as miR-126 into endothelial and tumor cells. Aptamer delivered pre-miR-126 (Ch3) was endogenously processed and inhibited the expression of the known miR-126-3p target VCAM-1. The pre-miR-126 aptamer chimera increased endothelial cell sprouting and affected tumor cell proliferation and endothelial cell recruitment.
To identify the best strategy for aptamer-mediated miR delivery, we generated several constructs using both the mature and precursor miR-126 sequences. Our data demonstrates that annealing the aptamer to the precursor molecule using a complementary stick sequence resulted in an increase in expression and biological activity of miR-126-3p. This strategy allowed the uptake of the aptamer and the processing into the mature miR. While linking mature forms of miR-126 in Ch1 and Ch2 also led to a significant increase in intracellular levels of miR-126-3p and -5p in HUVEC, the generated miR-126 appears to be nonfunctional. A possible explanation might be that Ch3 is recognized by the RISC complex and is processed by the cells comparable to endogenous pre-miR-126, which would also explain the preferential increase in miR-126-3p instead of miR-126-5p levels observed 24 hours after treatment with Ch3, which is consistent with the previously described natural processing of the isoforms in HUVEC cultured without shear stress [14]. This finding also emphasizes the importance of functional analysis when using synthetic tools to deliver microRNAs, since PCR-based detection of miRNAs is not distinguishing between processed, active miR-126 and the recombinant miR-126 that is part of synthetic chimeras. Moreover, effects on the intercellular localization cannot be detected by PCR.
Slightly different strategies have been used by other groups before: Dai et al. covalently linked the precursor to the MUC1 aptamer [35,36]. Another group used a polyethylene glycol spacer and conjugated a polyamidoamine-based miR to the aptamer [39]. Recently, Esposito et al. reported that the direct fusion of the let-7g passenger strand to the 3′ end of the GL21.T aptamer and subsequent annealing of the complementary guide strand of the miR allowed an effective delivery of miR to target cells in vitro and in vivo [37]. The strategy is similar to the one we used for Ch1 and it is unclear why this design did not result in a significant biologically relevant intracellular delivery in our study. However, it is difficult to compare our results to the other studies, since the copy numbers of delivered miRNAs are not shown in the published studies, and other cells and aptamers were used, but the extent of target gene repression and the functional effects shown in our study are at least comparable to the previous reports. Interestingly, despite the delivery of the complete pre-miR-126, we only detected a significant increase of miR-126-3p but observed only a trend for miR-126-5p. The reason underlying the selective increase is unclear. It might be related to the interference of the attached stick with processing into miR-126-5p. Alternatively, the selective processing might be due to the specific regulation of miR-126 processing in endothelial cells. Thus, the generation of mature miR-126-5p was specifically promoted by laminar flow or the flow-induced transcription factor KLF2, which post-transcriptionally regulated the formation of mature miR-126-5p versus miR-126-3p [14,44]. Since our experiments were performed under static conditions this may have resulted in the favorable processing of miR-126-3p from the precursor.
The present study for the first time addresses whether endothelial cells can be targeted by aptamer-linked miRs. Functionally active endothelial cells play a key role in the vasculature to protect against the formation of atherosclerotic lesions and are required for the formation of new blood vessels after ischemia [45]. MiR-126 has been implicated in both processes and thereby might be an interesting therapeutic target [11,44,46]. On the other hand, stimulating endothelial cell functions may also promote tumor growth [47], which may limit the use of miR-126-3p as pro-angiogenic, endothelial-protective strategy. However, in tumor cells miR-126 inhibits proliferation and by changing the paracrine activity of tumor cells interferes with the recruitment of endothelial cells and metastasis [16].
This study suggests that an enhanced tumor angiogenesis might be counteracted by direct effects on tumor cells and therefore may not be considered as risk of systemic miR-126 treatment. Of note, our data showed striking differences in the response of tumor cell lines. The different responses (inhibited proliferation of SK-BR3 cells but increased proliferation in MCF7 cells) are likely not related to the aptamer but to the response of the tumor cells to miR-126, since overexpression of pre-miR-126 induced similar differential responses. It is well known that target gene repression by miRNAs may be controlled in a cell-type specific manner (e.g. by the presence of RNA binding proteins, especially in tumor cells) [48,49]. Indeed, the expression of the miR-126 target Spred1 was not affected by miR-126 overexpression or Ch3-mediated miR-126 delivery in MCF7 cells but was changed in SK-BR3 cells. Therefore, the present study warrants a careful analysis of tumor cell responses using primary human cells. In the context of cancer, one might additionally consider using aptamers that allow a selective delivery of pre-miR-126 to specific subsets of tumor cells. Such selective aptamers have been reported and, for example, Her2-targeting aptamers might be used to selectively delivery miR-126 to Her2 overexpressing tumor cells [50].
Although our in vitro data suggest that pre-miR-126 might be an interesting candidate to treat cardiovascular diseases or cancer, further studies are necessary to determine the aptamer-mediated delivery of miRs in an in vivo setting.
Footnotes
Acknowledgments
We would like to thank Y. Manavski, A. Gatsiou, P. Schuster, M. Muhly-Reinholz, and A. Fischer for technical support, as well as Y. Feuermann for his aid with breast cancer cell cultures.
This work was carried out at the Institute for Cardiovascular Regeneration, Center of Molecular Medicine, Goethe University Frankfurt, Theodor-Stern-Kai 7, 60590 Frankfurt, Germany. The study was supported by the Landesoffensive für wissenschaftliche Exellenz (LOEWE; initiative to increase research excellence in the state of Hessen, Germany) as part of the Center for Cell and Gene Therapy; by the Deutsche Forschungsgemeinschaft (German Research Foundation) as part of the Sonderforschungsbereich (Collaborative Research Center) 902; and by the Deutsches Zentrum für Herz-Kreislauf-Forschung e. V. (German Center for Cardiovascular Research).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.