
Abstract
Although oligodeoxynucleotides containing CpG motifs (CpG-ODN) are potent immune stimulators, the use of natural CpG-ODN—phosphodiester-backbone CpG—has been limited due to its instability by nuclease in vivo. The aim of this study is to investigate the anticancer efficiency of CpG-ODN capsulated using liposome, which enhances the stability of CpG-ODN. We formulated lipoplex, encapsulated natural CpG-ODN from Mycobacterium bovis with liposome, and tested its immune stimulatory activity in vitro and in vivo. The lipoplex induced a systemic innate immune response in vivo and stimulated dendritic cells, but not macrophages, to stimulate proinflammatory cytokines such as tumor necrosis factor alpha and interleukin-6 in vitro. As expected, the lipoplex effectively mediated the prolonged cancer-therapeutic activity against B16 melanoma, which was dependent on natural killer and CD8+ T cells. The therapeutic activity was observed after only intratumoral administration of lipoplex among several treatment routes. Intratumoral treatment of lipoplex significantly increased the populations of natural killer and CD8+ T cells and reduced regulatory CD4+ T cell recruitment, which was correlated with expression profiles of chemokines (CCL1, CCL3, CXCL1, CXCL10, and CCL22). The antitumor therapeutic effect of lipoplex was dependent on the altered lymphocyte population that might be developed by the profile of intratumoral chemokine expression.
Introduction
T
Unmethylated oligodeoxynucleotides (ODNs) containing CpG motifs elicit innate immune responses and adaptive immune responses through binding to TLR9 [4]. In contrast to currently licensed adjuvants, CpG-ODN without antigen co-delivery can promote DC maturation, antigen presentation, and production of proinflammatory cytokines and chemokines. DCs maturated by CpG induce the production of interleukin (IL)-12, IL-18, and interferon (IFN)-γ and lead to enhanced T cell-, B cell-, and natural killer (NK) cell–mediated anti-tumor immune responses [5]. Indeed, CpG-ODN treatment without antigens has induced antitumor activity in clinical trials [6]. Although CpG-ODNs have strong immune stimulatory activities, CpG-ODN is not available for clinical use because the natural phosphodiester bond in CpG-ODN is too unstable to effectively stimulate the required immune response [7]. In addition, phosphorothionate-modified CpG-ODN (PS-ODN) is very stable but induces several side effects, including transient lymphoadenopathy, lymphoid follicle destruction, arthritis, and PS-ODN–specific immunoglobulin M ([8,9] in PS-ODN–treated mice in a CG sequence–dependent and backbone modification–dependent manner [10,11]. In a previous study, we constructed synthetic MB-ODN 4531, which originated from Mycobacterium bovis genomic DNA sequences, encapsulated with liposomes (lipoplex) to induce the optimal immune response without side effects and being degraded [12,13].
In this study, the immune stimulatory function and anti-cancer therapeutic effect of lipoplex were examined in vivo and in vitro. Lipoplex can promote a Th1-type immune response and strong inflammatory activity in DCs, but not in macrophages. Intratumoral (i.t.) administration of lipoplex into B16 melanoma-bearing mice induced anti-tumor activity by infiltration of T cells and NK cells and reduced the recruitment of regulatory T cells (Tregs). This antitumor activity was induced by stimulation of several inflammatory chemokines, which thereby altered the tumor microenvironment.
Materials and Methods
Lipoplex preparation
MB-ODN 4531 and its control, MB-ODN 4531GC, were obtained from Samchully Pharm and GenoTech, respectively. The MB-ODN 4531 originated from Mycobacterium tuberculosis and consists of 20 bases and 3 CpG motifs (underlined): AGCAG
Cell culture and mice
Female C57BL/6 mice aged 8 weeks were obtained from Koatech. All mice were kept at 23°C±1°C with a 12 h light/dark cycle. They had free access to water and diet. All procedures were reviewed and approved by the Animal Ethical Committee of Gyeongsang National University. Murine melanoma B16 cell line was grown in Dulbecco's modified eagle media supplemented with 10% fetal bovine serum (FBS; Sigma), 100 U/mL penicillin, and 100 μg/mL streptomycin (LONZA). Elicited macrophages of the C57BL/6 mice were obtained after intraperitoneal (i.p.) injection of a 4% thioglycollate broth (1 mL; Sigma) [15]. Three days later, peritoneal exudate cells were isolated from the peritoneal cavity. Then, the cells were cultured for 2 h in culture medium, and the adherent cells were used as peritoneal macrophages. Bone marrow-derived dendritic cells (BMDCs) from the C57BL/6 mice were prepared from the tibia and the femur [16]. After red blood cell lysis, the bone marrow cells were counted and seeded at a density of 3×106 cells in petri dishes containing complete medium supplemented with granulocyte macrophage colony-stimulating factor (PeproTech) for 10 days. Fresh medium was added at day 4 and day 7, and the cells were harvested, washed, and plated again. Finally, the BMDCs were assessed using CD11c staining (>95%). The macrophages and DCs were cultured in RPMI1640 supplemented with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin. The cells were gown at 37°C in a humidified atmosphere of 5% CO2.
Measurement of cytokines
The mice received i.p. injections of CpG 4531 or lipoplex (50 μg/200 μL for each) and the blood samples were taken from the retro-orbital sinus at 12 h. The macrophages and DCs were stimulated with 5 μg/mL of GC 4531, GC 4531 encapsulated with liposome (lipoplex GC 4531), CpG 4531, or lipoplex for 24 h. The cytokines from the blood sample or culture medium were analyzed using a cytometric bead array mouse inflammation kit or Mouse Th1/Th2 Cytokine kit (BD Biosciences).
Stimulation of OT-II mouse splenocytes
The spleen cells from the OT-II mouse were prepared by enzymatic digestion with collagenase (1 mg/mL; Sigma). The cell suspension was filtered and plated on tissue culture plates. After 2 h, the cells were stimulated with liposome, 5 μg/mL GC 4531, lipoplex GC 4531, CpG 4531, or lipoplex with or without 20 ng/mL of the OT-II OVA peptide (chicken OVA peptide 323-339 ISQAVHAAHAEINEAGR) and incubated for 48 h.
Tumor treatment and monitoring
The B16 tumor cells (1×106) were subcutaneously implanted into the mice. Subsequently, the mice received phosphate buffered saline (PBS) (50 μL), CpG 4531 (12.5 μg/ 50 μL), or lipoplex (12. 5 μg/50 μL) by i.t., intravenous (i.v.), intramuscular (i.m.), or i.p. injection on days 12, 15, and 18 after the initial implantation. The tumor volumes were estimated on days 12, 15, 18, 24, and 28 after the injection.
Lymphocyte depletion
The monoclonal anti-NK1.1 antibody (Ab; PK136) was used for NK cell depletion, the anti-CD4 Ab (GK 1.5) was used for CD4+ T cell depletion, and the anti-CD8 Ab (Yis 169.4.2) was used for CD8+ T cell depletion. The depletion was confirmed using flow cytometry. The mice were i.p. administrated with 250 μg of each Ab 3 times, at 12, 15, and 18 days after tumor injection. Lipoplex was administrated i.t. three times, on day 14, day 17, and day 20 after tumor injection. The tumor volumes were estimated on days 14, 17, and 20 after the injection.
Analysis of tumor-infiltrated lymphocytes
The B16 tumor-bearing mice were killed at 24 h after the final injection of CpG 4531 or lipoplex, and the tumors were resected. The chapped tissues were incubated with 5 mL of collagenase (1 mg/mL; Sigma) under shaking conditions for 20 min and filtered using a cell strainer (BD). The prepared samples were washed and incubated with red blood cell lysis buffer for 5 min. After the incubation, the cells were washed and stained with anti-CD4, anti-CD8, anti-CD45, and anti-NK1.1 Abs (BD) for 30 min. If necessary, the cells were washed and fixed with the Foxp3 staining kit (eBioscience) and stained with anti-Foxp3 Ab for 30 min.
Chemokine mRNA expression
The B16 tumor-bearing mice were killed at 24 h after the final injection of CpG 4531 or lipoplex, and the tumors were resected and weighed. Total RNA was isolated from tissues using a total RNA isolation solution (Riboex, GeneAll), and the cDNA was synthesized by using oligo-dT primers and reverse transcriptase (Fermentas). Real-time polymerase chain reaction (PCR) was performed using the SSoFast EvaGreen Supermix and CFX96 real-time detection system (Bio-Rad Laboratories). The following primers were used for amplification: CCL1, 5′-GGA TGT TGA CAG CAA GAG C-3′ and 5′-CAG GGG TTC ACC TTC TTC A-3′; CCl3, 5′-ATC ATG AAG GTC TCC ACC AC-3′ and 5′-TCT CAG GCA TTC AGT TCC AG-3′; CXCL1, 5′-CGC TTC TCT GTG CAG CGC-3′ and 5′-AAG CCT CGC GAC CAT TCT T-3′; CXCL10, 5′-GGA AGC CTC CCC ATC AGC AC-3′ and 5′-AGA GAG GCT CTC TGC TGT C-3′; CCL22, 5′-ATG TGA GGC CAA ATA GAC GAA-3′ and 5′-CTG GCA CTG TCA ATC CCT G-3′; GAPDH, 5′-CCA TCA CCA TCT TCC AGG AG-3′ and 5′-ACA GTC TTC TGG GTG GCA GT-3′.
Statistical analysis
Student's t-test was used. Error bars represent the standard error of the mean. A p value<0.05 was considered statistically significant.
Results
Lipoplex induced immune response through DCs
To test whether lipoplex effectively induces an immune response, the mice were administrated a single i.p. injection of naked CpG 4531, lipoplex, naked GC 4531, or lipoplex GC 4531. Elevated tumor necrosis factor alpha (TNF-α) production was observed in the lipoplex-treated mice, whereas CpG 4531, GC 4531, and lipoplex GC did not induce TNF-α production (Fig. 1A). To investigate which cells were activated by lipoplex, the peritoneal macrophages and BMDCs were treated with liposome, CpG 4531, or lipoplex to examine which innate cells are targeted by lipoplex. The production of TNF-α and IL-6 was significantly increased in the DCs treated with lipoplex, but not in the macrophages (Fig. 1B). These results might be due to the high lipoplex uptake efficiency of DCs compared with macrophages (Fig. 1C). Next, we examined the uptake efficacy of DCs to CpG 4531 and lipoplex. Lipoplex was taken up by the DCs within a short time, but the naked CpG 4531 was not (Fig. 1D).
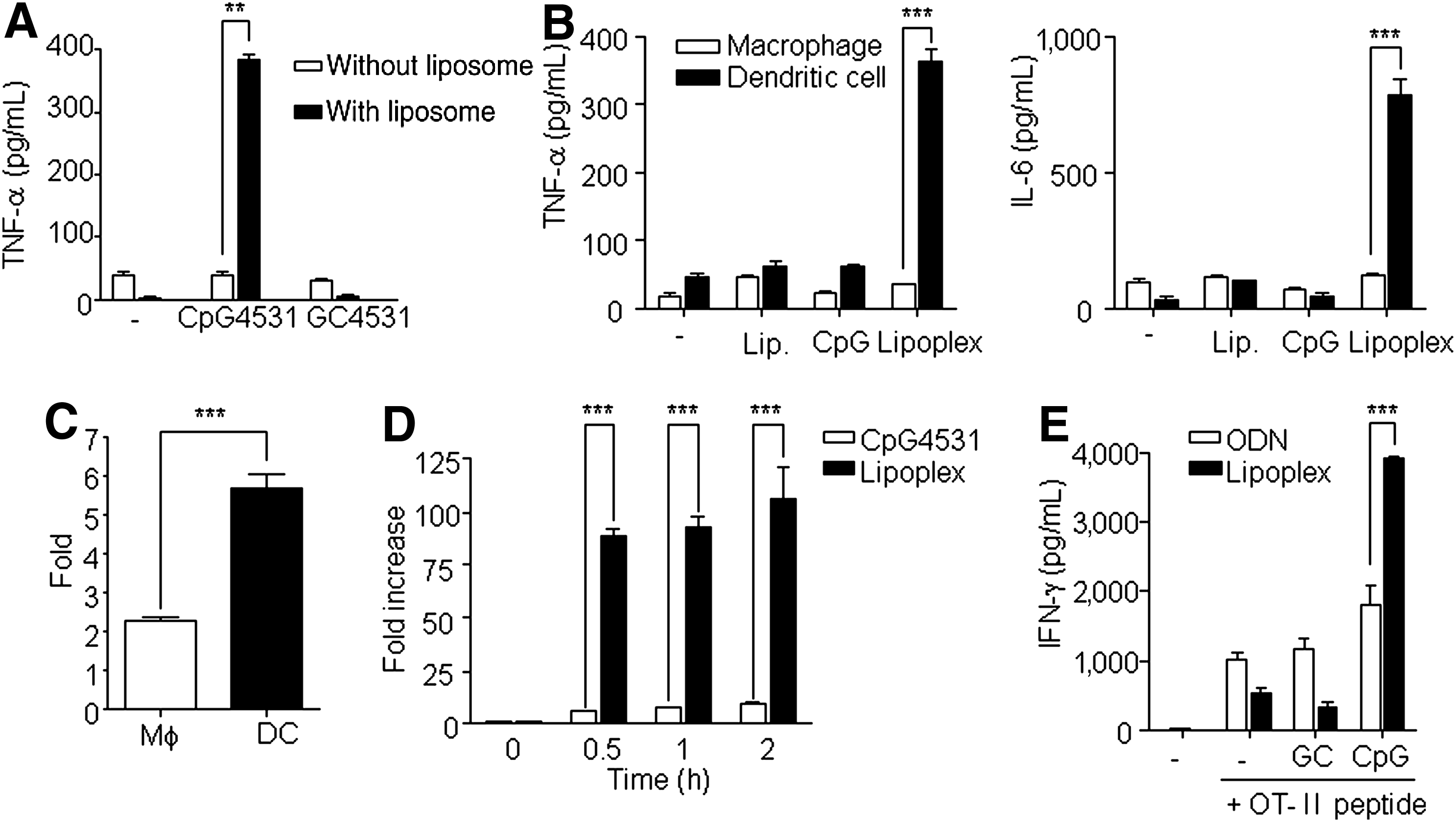
Immune stimulatory activity of lipoplex.
To test whether lipoplex could induce antigen-specific Th1 immune response, splenocytes of the OT-II Tg mouse were stimulated under the indicated conditions for 48 h (Fig. 1E). The results analyzing the antigen specific CD4+ T cell activation revealed that lipoplex induced a Th1-mediated immune response, which induced IFN-γ. The amounts of IFN-γ, which was mainly produced by the Th1 cells, were increased in the presence of lipoplex and the OT-II peptide complex. Comparatively, IL-4, which is produced by the Th2 cells, was not detected in the control or in the presence of lipoplex or the OT-II peptide complex (data not shown). Therefore, these data suggest that capsulation of CpG with liposome increased its ability to be taken up by DCs, which effectively induced immunostimulatory activity involving both innate and adaptive immunity.
Lipoplex induced anti-tumor activity
The aforementioned data showed that lipoplex induced the Th1 immune response, which promotes the cell-mediated immune response and cytotoxic T lymphocyte (CTL) response [17]. Because activated CTLs are critical components in the treatment of tumors, we examined the antitumor activity of lipoplex using B16 melanoma as a tumor model. Mice were s.c. implanted with B16 tumor cells. On days 12, 15, and 18 after implantation, PBS, CpG 4531, or lipoplex were i.t. injected. As expected, treatment with lipoplex attenuated the tumor growth (Fig. 2A). To test whether a specific lymphocyte population is necessary for lipoplex mediated-tumor therapy, anti-CD4 mAb, anti-CD8 mAb, and anti-NK mAb were used for the depletion of each lymphocyte in vivo. Treatment with each antibody caused a loss of 98% of each lymphocyte in peripheral blood cells (data not shown). Our results also revealed that NK and CD8+ T cells are essential for lipoplex-mediated tumor therapy (Fig. 2B). CpG-ODN induced different effects with respect to tumor growth inhibition depending on the route of administration [18]. Therefore, we explored the role of the administration route (i.p., i.t., i.m., and i.v.) on the anti-tumor effect of lipoplex. As shown in Fig. 2C, only i.t. injection of lipoplex induced an antitumor growth effect.
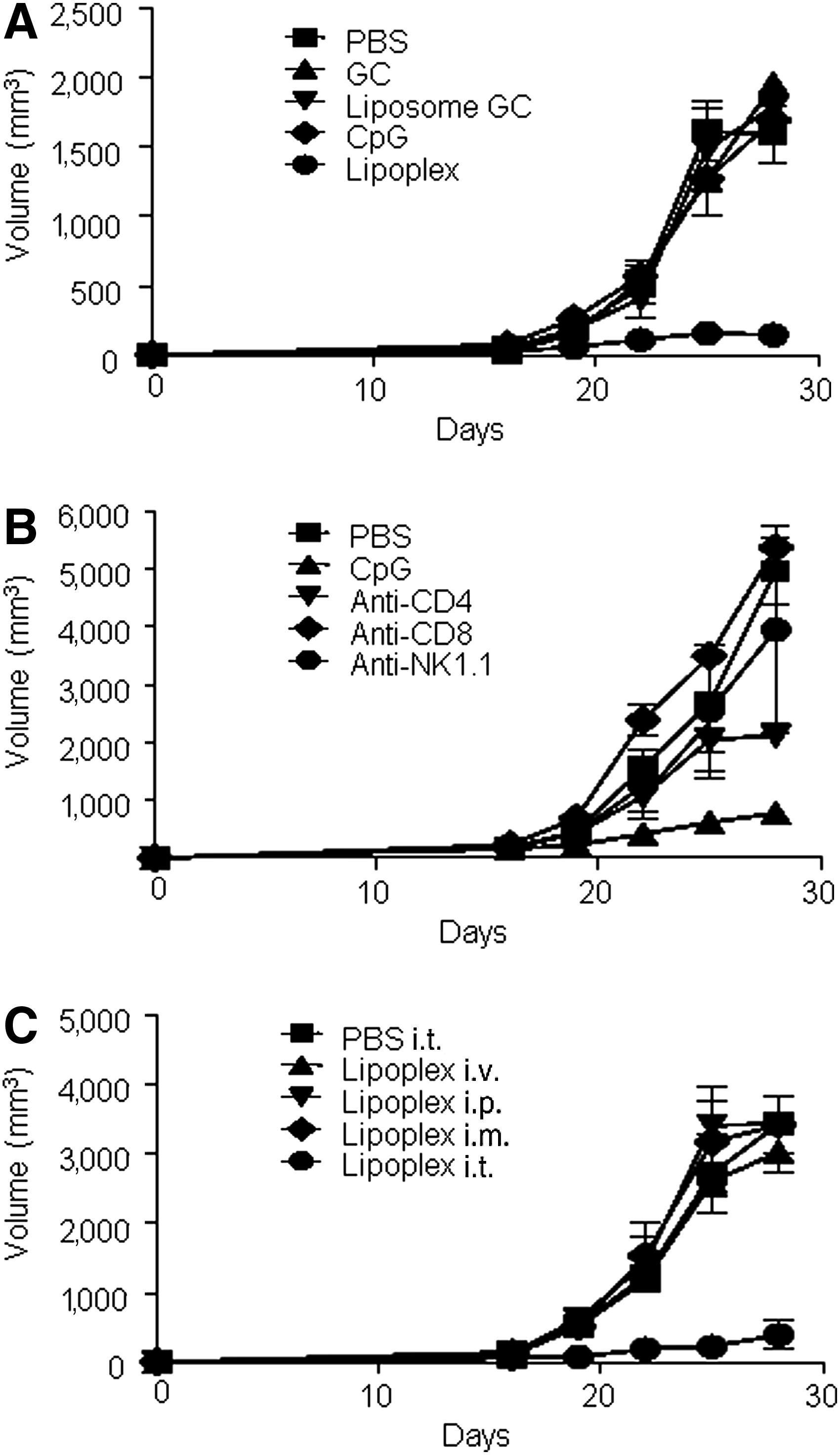
Antitumor activity of lipoplex.
Lipoplex enhanced the intratumoral infiltration of NK and CD8+ T cells
The previous results suggested that the local immune environment and NK and CD8+ T cells played important roles in the antitumor activity of lipoplex. Therefore, we analyzed the infiltration of lymphocytes into tumor tissue using flow cytometry. As expected, treatment with lipoplex, but not CpG 4531, induced the infiltration of CD45+ lymphocytes (Fig. 3A), CD8+ T (Fig. 3B), and NK cells (Fig. 3C). The recruitment of NK and CD8+ T cells into tumor tissues by lipoplex indicated that the expression of lymphocyte-attracting chemokines might be elevated in the tumor microenvironment. Therefore, we explored the effect of CpG4531 and lipoplex treatments on the mRNA expression profiles of various chemokines (CCL1, CCL3, CXCL1, and CXCL10) in the tumor tissue. These proinflammatory chemokines regulate intratumoral trafficking of leukocytes, including T cells and NK cells, and inhibit tumor growth [19–22]. As shown in Fig. 3D, the administration of lipoplex enhanced the expression of inflammatory chemokines.

Accumulation of CD8+ T and NK cells at the tumor site. B16 melanoma-bearing mice were administered i.t. CpG 4531 or lipoplex at days 12, 15, and 18, and the tumor was resected and sized on day 20. The tumor tissues were homogenized, treated with collagenase, filtered, and stained with CD45, CD8, and NK for flow cytometry analysis. The percentages of tumor-infiltrating CD45+
Lipoplex inhibited Treg trafficking into tumor tissue
We further tested the ability of lipoplex to regulate the recruitment of Foxp3+ Tregs. As shown in Fig. 4A, the population of Foxp3+ Tregs in the tumor microenvironment was reduced by treatment of lipoplex, whereas there were no observed differences in the draining lymph node following treatment with PBS, CpG 4531, or lipoplex. These results suggest that lipoplex attenuated the recruitment of Foxp3+ Treg into tumor tissues. To determine whether the decreased Foxp3+ Treg accumulation at the tumor site was due to decreased levels of CCL22, which accumulates in Foxp3+ Treg cells [23], we performed real-time PCR for this chemokine. As shown in Fig. 4B, CCL22 expression was downregulated by lipoplex treatment.
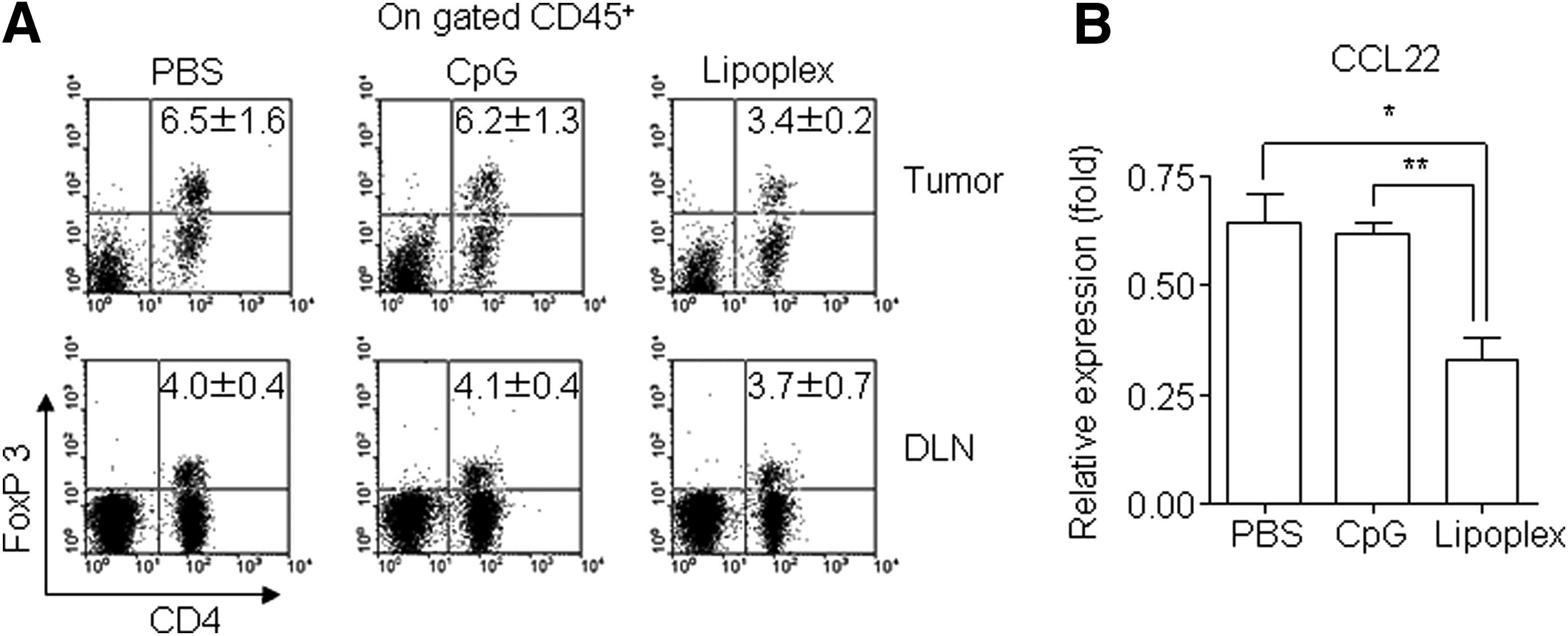
Regulatory T cell (Treg) infiltration in tumor tissues. Tumor tissues were harvested from the B16 tumor–bearing mice at 2 days after the final injection of lipoplex.
Discussion
It has been well established that CpG motifs are recognized by the innate immune system via interaction with the PRR TLR9 [24,25]. The CpG motif promotes an immune response that activates immune cells, such as DCs, macrophages, NK cells, B cells, and T cells, and induces secretion of various cytokines and chemokines [26]. Although CpG-ODN has strong immunostimulatory activities, the natural phosphodiester bond CpG-ODN is not viable in vivo because it is rapidly degraded by nucleases [27,28]. To solve this problem, we constructed lipoplex that induces an optimal immune response with slower degradation rates compared to CpG-ODN [14]. In this study, we have shown that lipoplex induces the production of proinflammatory cytokines to a greater extent than CpG 4531. It is well established that encapsulating CpG-ODN in liposomes facilitates their uptake by macrophages and DCs and increases their immunostimulatory activity [29,30]. Our study suggests that DCs are the main target cells of lipoplex and that DCs stimulated by lipoplex produce greater amounts of cytokines such as TNF-α and IL-6 than macrophages. The liposomes used in this study were a mixture of the pH-sensitive liposomes DOPE and CHEMS [31]. Therefore, the DOPE:CHEMS formulations were stable at neutral and basic pH, but not at acidic pH [32]. Phagosomal pH in the DCs is >7, whereas the pH in macrophages is <6 [33]. Consequently, lipoplex might be unstable in macrophages and therefore unable to effectively stimulate macrophages compared with DCs.
Upon activation, DCs rapidly produce type 1 IFNs, which in turn activates NK cells, B cells, and T cells, thereby leading to the induction of innate and adaptive immune responses [34]. CD4+ T cells provoked with antigen-specific stimuli proliferate and differentiate into either the Th1 or Th2 subset. Systemic administration of CpG-ODN induces the Th1 cell immune response that inhibits the development of the Th2 type responses such as allergic asthma [35]. In our study, we revealed that lipoplex induced a greater Th1-mediated immune response with respect to antigen specific CD4+ T cell activation compared to CpG 4531. The Th1 immune response would be required for immunological memory of tumor-specific CTLs [36]. CTLs recognize tumor-associated antigens from tumor cells through uptake and processing by DCs and they subsequently induce tumor-specific immunity [37]. Indeed, we showed that lipoplex has the potential to inhibit tumor growth and that the anti-tumoral activity of lipoplex is associated with the presence of NK and CD8+ T cells. Although the lipoplex had a similar immune stimulatory effect to PS-ODN, usage of lipoplex would be available due to low side effect compared with that of PS-ODN [8–14].
The anti-tumor therapeutic effect of CpG-ODN is known to be dependent on injection route [18] because the absorption and accumulation rate varies with the administration site [38,39], and the cytokine production in tumor cells varies depending on the route used to inject liposome-capsulated CpGs [40]. In this context, we analyzed the role of the administration route in the anti-tumor activity of lipoplex. We revealed that only i.t. injection of lipoplex inhibited tumor growth, not i.p., i.v., or i.m. These results suggest that lipoplex activates a local immune response in the tumor environment. Indeed, we found that i.t. injection of lipoplex induced the recruitment of NK and CD8+ T cells at the tumor site, which was crucial for the anti-tumor activity of lipoplex.
Recently, chemokines have been utilized in tumor immunotherapy as a means of recruiting effector immune cells [41]. Tumor infiltrating lymphocytes produce chemokines, such as CCL1, CCL3, CXCL1, and CXCL10, which promote the migration of NK and CD8+ T cells to the tumor site [19–22,42]. These findings are in accordance with our observation that lipoplex induced infiltration of NK and CD8+ T cells into the tumor, thereby resulting in antitumor activity. A recent study showed that i.t injection of CpG-ODN induces differentiation and reduces the immunosuppressive activity of myeloid-derived suppressor cells [43]. In addition to these functions of CPG-ODN, our data showed another important contribution of lipoplex in antitumor activity; we demonstrated the ability of lipoplex to downregulate CCL22 expression and reduce the amounts of Foxp3+ Tregs accumulated at the tumor site. CD4+CD25+ Tregs can potently inhibit CD8+ T and NK cell responses, which leads to tumor-specific CTL activity [44–46]. Therefore, the ability of lipoplex to inhibit the recruitment of Foxp3+ Tregs to the tumor site might contribute to tumor regression.
The selective delivery of anticancer drugs to tumor sites remains an important aim of cancer therapy. Many studies demonstrating the benefits of lipid-based delivery, including protection from the nucleic acid payload, improved pharmacokinetic characteristics, enhanced intracellular uptake, and enhanced delivery to target antigen-presenting cells of CpG-ODN [4]. Additionally our results further our understanding of lipid-based delivery systems by demonstrating that i.t. injection of lipoplex might be the optimal route of delivery for cancer therapy.
In conclusion, our data suggests that administration of lipoplex inhibits tumor growth and induces infiltration of NK and CD8+ T cells to the tumor site by inducing proinflammatory chemokine expression, which alters the anti-tumor activity. Furthermore, i.t. injection of lipoplex inhibits the recruitment of Foxp3+ Tregs to the tumor site, and therefore, i.t. administration is the most efficient method for the application of lipoplex in tumor regression therapies.
Conclusions
CpG-ODN capsulated with liposome (lipolex) was a strong immune stimulator that activated dendritic cell in vitro and induced TNF-α in vivo. The anticancer activity of lipoplex was limited to i.t. administration route only, not other routes (i.p., i.v., i.m.). The activity was dependent on lymphocytes, NK and CD8+ T cells, and the lymphocytes were highly infiltrated in tumor tissue. Additionally, Treg infiltration was reduced by lipoplex administration. Chemokine expression patterns were correlated with intratumoral lymphocyte infiltration. Taken together, the antitumor therapeutic effect of lipoplex might be dependent on alterations to the chemokine environment in tumor tissue as well as the induction of innate and acquired immune responses.
Footnotes
Acknowledgment
This study is supported by next-generation biogreen21 (SSAC, PJ01107002), Rural Development Administration, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.