
Abstract
Nucleoside analogues are the most promising drugs for the treatment of pancreatic cancer to date. However, their use is often limited due to toxic side effects. Aptamer-mediated targeted delivery of these drugs to cancer cells could maximize their effectiveness and concomitantly minimize the toxic side effects by reducing uptake into normal cells. Previously, we identified a pancreatic cancer-specific, nuclease-resistant RNA aptamer, SQ2, which binds to alkaline phosphatase placental-like 2 (ALPPL2), a putative biomarker for pancreatic cancer. In this study, we demonstrate that the aptamer can be internalized into pancreatic cancer cells and can thus be used for the targeted delivery of therapeutics. Using the aptamer as a ligand, we established that glycophosphatidylinositol-anchored ALPPL2 is internalized by the cells through clathrin-independent and caveolae-dependent or dynamin-mediated cell-type-dependent pathways. Finally, we show that SQ2 can deliver nucleoside drug 5-fluoro-2′-deoxyuridine specifically to ALPPL2-expressing pancreatic cancer cells, inhibiting cell proliferation.
Introduction
P
5FU has been used as a component of standard therapy for pancreatic cancer for more than 40 years. Metabolites of 5FU affect cell proliferation and viability through three different mechanisms: inhibition of RNA maturation [7,8], DNA cleavage [9], and inhibition of thymidylate synthase [10], the key enzyme for the de novo biosynthesis of thymidine. The latter mode of action has the most significant contribution to the activity of 5FU and occurs by the conversion to FdUMP [11]. However, 5FU has limited use in pancreatic cancer due to its toxic side effects. To circumvent these side effects, 5-fluoro-2′-deoxyuridine (5FdU), a less toxic analogue of 5FU [12] and also the unphosphorylated precursor of FdUMP [13], can be administered. In addition, the targeted delivery of 5FdU could further overcome toxicity to nontargeted cells.
Previously, we reported the Cell-SELEX-based identification of a 2′F-modified RNA aptamer, SQ2, which binds specifically to pancreatic cancer cells [14]. The target protein of SQ2 was found to be a glycophosphatidylinositol-anchored protein (GPI-AP), namely alkaline phosphatase placental-like 2 (ALPPL2). The expression of ALPPL2 protein was then confirmed in a panel of pancreatic cancer cells, while no mRNA or protein expression was observed in normal pancreatic ductal cells. Furthermore, membrane-bound ALPPL2 can also be detected in cell secretions through the action of phospholipase-mediated cleavage of the phosphatidyl anchor. The ectopic expression of this protein in pancreatic cancer cells not only makes it a very useful serum- and tissue-based biomarker but also an attractive candidate for the development of targeted therapy. Although ALPPL2 and its closely related family member, placental alkaline phosphatase (PLAP), have not yet been studied in great detail, earlier studies on PLAP have shown that, like other GPI-APs, this protein can also be internalized, and this mainly occurs by cholesterol-dependent endocytosis [15,16].
The truncated form of the aptamer SQ2 is 25 nucleotides in length and hence suitable for efficient chemical synthesis. It binds to its target with high specificity and nanomolar affinity (Kd: 20 nM). In addition, it is possible to make modifications/conjugations at both the 5′- and 3′-end without affecting the target binding capacity. All these features make SQ2 highly suited for the development of aptamer-based targeted therapeutic strategies for cancer. In this study, we explored the use of SQ2 in the targeted therapy of pancreatic cancer. Using SQ2 as a ligand, we first confirmed that the target protein, ALPPL2, efficiently internalizes and carries the aptamer-associated cargo inside the cells. We then validated the use of SQ2 in the targeted delivery of the pancreatic cancer drug 5FdU. Treatment of ALPPL2+ve pancreatic cells with SQ2 carrying a 5FdU pentamer at the 3′-end led to inhibition of cell growth, while not affecting the normal pancreatic ductal cells that did not express the target protein.
Materials and Methods
Cell culture
The pancreatic cancer cell line Capan-1 was obtained from the American Tissue Culture Collection (ATCC) and cultured in IMDM supplemented with 20% fetal calf serum (FCS; Invitrogen) and 100 U/mL penicillin–streptomycin (Invitrogen). ALPPL2+ve Panc-1 cells (hereafter Panc-1+ve cells) were enriched as described earlier [14] and cultured in DMEM supplemented with 10% FCS. Human pancreatic ductal epithelial cells (HPDE) were obtained from Ming-Sound Tsao, UHN, Canada, and cultured in keratinocyte serum-free media (Invitrogen), as described previously [17].
Oligonucleotides
3′ TAMRA- or biotin-labeled SQ2(6–35) (5′-CUUAUUCAAUUGCCUGAAAAGCUAUCGC-3′), which is the truncated form of the 2′F-modified aptamer SQ2 (simply referred to as SQ2 in this article) and mutant SQ2 with the sequence 5′-AGCUUAUUCAAUU
Internalization assay
Cells grown in 12-well plates were treated with the previously reported concentrations of endosomal inhibitors [18], such as methyl β cyclodextrin (MβCD; 5 mM), chlorpromazine (5 μg/mL), and genistein (400 μM) (Sigma Aldrich), in serum-free medium for 1 h. The pretreated cells were then incubated with Alexa fluor 680-labeled transferrin (10 μg/mL) or BODIPY-fl-c5-lactosylceramide (LacCer, 0.8 μM) (Molecular probes) for 15 min. In the case of aptamer treatment, cells pretreated with inhibitors were further incubated with 50 nM TAMRA-labeled SQ2 or mutant SQ2 for 30 min in the same media. For treatment with RNase cocktail, cells were washed with binding buffer (Dulbecco's phosphate-buffered saline containing 4.5 g/L glucose and 5 mM MgCl2) and treated with 0.1 U/μL of RiboShredder (Epicentre Biotechnologies) in binding buffer containing 0.1% sodium azide for 10 min at room temperature (RT) [19]. The cells were collected by gentle scraping, after which the cell pellets were washed twice in the same buffer. Alternatively, for high salt wash, cells were washed in cold 0.5 M NaCl buffer and incubated further in the same buffer on ice for 10 min. Cells were washed twice before further analysis.
Microscopy
Cells cultured in confocal dishes were incubated with 50 nM of TAMRA-labeled SQ2 or mutant SQ2 aptamers in 150 μL of culture medium supplemented with 5 μL of 10 mg/mL yeast tRNA and 5 μL of 100 mg/mL bovine serum albumin (BSA) for 30 min at 4°C for binding or at 37°C for internalization, respectively. To inhibit the cellular energy pool, cells were treated with the metabolic inhibitor sodium azide (15 mM) in combination with 2-deoxyglucose (10 mM) (Az-dG) for 15 min in glucose-free phosphate buffer saline, followed by incubation with SQ2-TAMRA in the presence of the inhibitors. Fluorescence was detected using a CSRL700 confocal microscope (ZEISS) with Z sections of 1 μm thickness or a fluorescent microscope (Olympus IX81) with Metaview imaging software at 400× magnification. For the internalization of magnetic beads, Panc-1+ve cells grown in confocal dishes were incubated with 50 nM of SQ2-biotin or mutant SQ2-biotin in the culture medium at 4°C. The cells were then washed gently to remove unbound oligonucleotides and incubated with 5 μL of streptavidin-coated magnetic beads (Dynabeads Thermo Scientific) in the culture medium at 37°C for 1 h. The cells were washed two to three times to remove unbound beads. Images were acquired with a confocal microscope at 400× with Z sectioning. To obtain a clear view of the cell sections in the case of mutant aptamer, cells were spiked with additional beads during the image acquisition.
Internalization assay and NucleoCounter NC-3000 analysis
To quantify the amount of aptamer internalized versus that bound, cells grown in 12-well plates were incubated with 50 nM of TAMRA-labeled SQ2 or mutant in 500 μL of the culture medium supplemented with yeast tRNA and BSA for 1 h, followed by the addition of 10 μg/mL of Hoechst 33342 with further incubation for an additional 15 min. In the case of internalized fractions, the cells were then treated with RiboShredder or high salt wash, as mentioned above. The cells were resuspended in 50 μL of the culture medium and immediately analyzed on a NucleoCounter NC-3000 (Chemometec) using user-adaptable protocols at an Ex/Em of 530/620 nm. A total of 5,000 cells were analyzed, from which the mean fluorescence intensity of SQ2-TAMRA was calculated.
5FdU treatment and cell counting
Capan1 or HPDE cells grown in 96-well plates were treated in triplicate with free 5FdU and 5FdU-conjugated oligonucleotides or with control oligonucleotides in serum-free media for 2 h at the indicated concentrations. After incubation, the cells were washed twice with culture media and incubated in complete media for 48 h. The total viable cell population was determined by trypan blue dye exclusion assay and represented as percent of the untreated control.
Quantitative real-time polymerase chain reaction
Following the binding and internalization assay, mRNA was extracted using isol-RNA lysis reagent (5 PRIME, Inc.). Five hundred nanograms of total RNA was then reverse-transcribed with 50 pmoles of SQ2 reverse primer (5′ GGCGATAGCTTTTCAGGC 3′) using the ImProm-II™ Reverse Transcription System (Promega), according to the manufacturer's protocol. Aliquots of the cDNA reaction mixture were analyzed by quantitative real-time polymerase chain reaction (PCR) on a Step-One real-time PCR machine (Applied Biosystems) using SQ2 forward (5′ ATACCGGGAGCTTATTCAATT 3′) and reverse primers. Quantitative PCR data were shown as values relative to mutant SQ2.
Statistical analysis
All experiments were performed at least in triplicates for reproducibility, and the data were represented as mean±SE of three or more independent experiments. Student's t-test with a two-sided alpha of 0.05 was used for comparison of the differences between treatment groups.
Results and Discussion
ALPPL2-binding aptamer SQ2 can be internalized into pancreatic cancer cells
As shown in our previous findings, the ALPPL2-binding aptamer SQ2 displayed binding to the pancreatic cancer cell line Capan-1 and Panc-1+ve cells (enriched population of ALPPL2-expressing Panc-1 cells). HPDE, normal pancreatic ductal cells that do not express ALPPL2, did not show any SQ2 binding (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/nat). To further explore the utility of SQ2 in targeted therapeutics of pancreatic cancer, we first tested whether the aptamer could be internalized into ALPPL2-expressing pancreatic cancer cells. Capan-1 and Panc-1+ve cells were incubated with TAMRA-labeled SQ2 at 37°C to allow aptamer internalization. For clear visualization of the internalized fraction, the cell surface-bound aptamers were removed by treatment with RiboShredder, a harsh cocktail of RNases. 2′-Fluoropyrimidine-modified RNA aptamers, while relatively resistant to nuclease degradation under physiological conditions, can be cleaved in vitro by treatment with RiboShredder [19]. It was confirmed that SQ2 could be completely degraded by RiboShredder after incubation for 10 min at RT (data not shown). In addition, the treatment was carried out in the presence of 0.1% sodium azide to inhibit internalization of any free TAMRA fluorochrome generated upon degradation of the RNA oligonucleotide. The RNase-treated cells clearly demonstrated that SQ2 can be internalized by Capan-1 and Panc-1+ve cells within 30 min of incubation (Fig. 1A). No internalization was seen in HPDE cells.
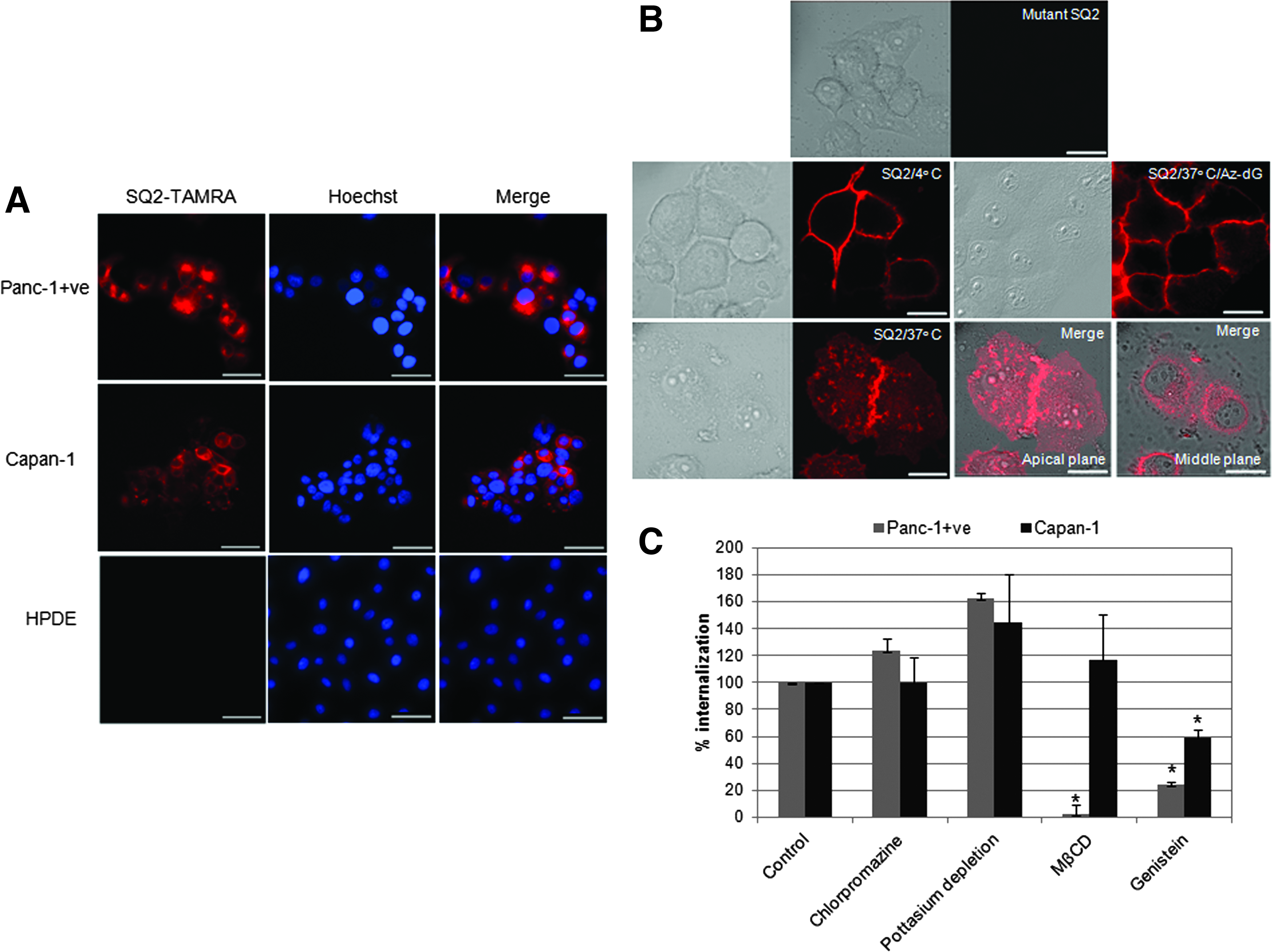
Aptamer SQ2 can specifically internalize ALPPL2+ve pancreatic cancer cells through clathrin-independent endocytic uptake.
ALPPL2-mediated clathrin-independent pathways are involved in the internalization of SQ2
Although the membrane raft-mediated caveolae-dependent internalization of PLAP, a homologue of ALPPL2, was reported earlier, there is no information regarding the cellular transport mechanism of ALPPL2. ALPPL2 closely resembles PLAP, hence similar transport and sorting pathways can be expected for this protein. To first confirm that the ALPPL2-mediated cellular uptake of SQ2 is an energy-dependent process, we examined whether the aptamer internalization could be inhibited by low temperature, as well as ATPase inhibition (by treatment with Az-dG). The Panc 1+ve cells incubated with 50 nM of SQ2-TAMRA at 37°C showed the presence of SQ2 throughout the cell. However, when treated at 4°C and upon depletion of cellular energy, SQ2-TAMRA was present exclusively on the cell membrane, suggesting energy-dependent endocytotic uptake of the SQ2 aptamer (Fig. 1B). Furthermore, Z sectioning of the cells was performed to allow clear visualization of the localization of the internalized fraction. SQ2 was clearly observed in the perinuclear space, confirming its cellular uptake by endocytosis. Like most GPI-anchored proteins, PLAP is also believed to be endocytosed through raft-dependent caveolar endocytosis [15,16]. However, these findings were mainly based on the usage of primary and secondary antibodies that act as multivalent ligands, which can cause multimerization of the GPI-APs and their forced sequestration in cholesterol-rich clusters [20]. Research into the endocytosis of GPI-APs has shown that the cell surface organization of GPI-APs into monomers or multimers may directly influence the endocytic pathways involved and the subsequent intracellular destinations [20,21]. Studies on the endocytosis of noncross-linked GPI-APs using monovalent ligands have been possible with a few GPI-APs, which clearly revealed discrepancy in the endocytosis mechanisms earlier established through antibody-based studies [22]. This highlights the importance of monovalent natural or artificial ligands in delineating the endocytic pathways of GPI-APs. In this regard, unlike antibody, aptamer can be an ideal tool to study the endocytosis of GPI-APs.
As the endocytosis of PLAP is poorly understood, using SQ2 as a ligand, we performed additional studies which could provide insight into the mechanism of endocytosis of plasma membrane-tethered ALPPL2. The endocytosis of SQ2-TAMRA was examined by internalization assay in cells treated with several well-established endosomal inhibitors. Treatment with chlorpromazine, a cationic amphiphilic drug, and potassium depletion, both believed to block clathrin-coated pit formation, were used to block clathrin-mediated endocytosis [23,24]. For clathrin-independent pathways, MβCD and genistein were employed. MβCD blocks cholesterol-dependent endocytic processes by reversibly extracting the steroid from the plasma membrane [25], whereas genistein is a tyrosine kinase inhibitor that causes local disruption of the actin network at the site of endocytosis and inhibits the recruitment of dynamin II. Both these inhibitors block events that are indispensable for caveolae-mediated endocytosis [26]. We first confirmed that the concentrations of inhibitors used for this study caused no toxicity. Although some morphological changes were observed, no toxicity was observed by the MTT cytotoxicity assay upon treatment with the endocytic inhibitors (data not shown). To verify the action of the inhibitors, Alexa fluor 680-labeled transferrin was used as a positive control for clathrin-mediated endocytosis, and BODIPY-fl-c5-lactosylceramide (LacCer) as a positive control for caveolae-mediated endocytosis. Microscopic analysis of Capan-1 and Panc-1+ve cells treated with the inhibitors and probed with the respective positive and negative controls confirmed that the concentrations of endocytic inhibitors used in this study were appropriate (Supplementary Fig. S2).
Next, the effects of these inhibitors on the internalization of SQ2-TAMRA were examined. In both the Capan-1 and Panc-1+ve cells, inhibition of clathrin-mediated endocytosis did not cause any reduction in the levels of SQ2 internalized. Conversely, a slight increase in the SQ2 internalization was observed. It is well known that the inhibition of one endocytic pathway can lead to upregulation of other alternative pathways as a compensatory mechanism [18,27]. If such alternative pathways are utilized for SQ2 internalization, then the increased cellular uptake can be explained. Unlike the other inhibitors, treatment with MβCD resulted in a 40% reduction of SQ2 binding in Panc-1+ve cells (Supplementary Fig. S3) and nearly a complete loss of aptamer internalization (Fig. 1C). This suggests that the ALPPL2 is present largely in cholesterol-rich fraction, and upon depletion of cholesterol from the plasma membrane, a part of this GPI-AP sheds off and most of the remaining becomes incapable of facilitating caveolae-mediated endocytosis. This is in complete agreement with the earlier published studies on PLAP [16]. Strangely, MβCD had no effect on the binding and internalization of SQ2 in the Capan-1 cells. At higher concentration of 10 mM, distinct morphological effects of MβCD on Capan1 were visible, yet no effect on SQ2 binding and internalization was observed, suggesting that ALPPL2 membrane localization and endocytosis are not dependent on cholesterol in these cells. However, upon treatment with genistein, Capan-1 (40%) and Panc-1+ve (78%) cells showed significant reduction in the SQ2 internalization.
Taken together, these results clearly implicated caveolae-mediated endocytosis as the predominant mechanism of ALPPL2 internalization in Panc-1+ve cells. In contrast, for the Capan-1 cells, ALPPL2 endocytosis seemed to be independent of both caveolae- and clathrin-mediated endocytosis. Caveolae-1, which is an indispensable protein for the formation of caveolae, is expressed at extremely low levels in Capan-1 cells [28,29]. This suggests the presence of alternative caveolin-independent pathways, as seen earlier in caveolae-null mice [30]. Flotillin, which has a similar topology to caveolae, generates structures that look like caveolae [31] and were reported to be employed in GPI-AP endocytosis [32]. This pathway is dependent on dynamin II and Src-family kinases [33], which can be inhibited by genistein. Although studies with additional more specific inhibitors are warranted to pinpoint the exact endocytic pathway operative in the endocytosis of ALPPL2 in Capan-1 cells, the current data suggest that ALPPL2 is internalized through a clathrin- and caveolae-independent, but dynamin-dependent, pathway. Thus, in both the pancreatic cancer cells, SQ2 internalize through distinct endocytic pathways.
ALPPL2-targeting aptamer can be used as a tool for targeted delivery
Although the efficiency of aptamers in targeted delivery approaches can be limited by various factors, the basic rules that govern aptamer efficacy are its target binding affinity, tissue specificity, and the intracellular concentrations that can be achieved. SQ2 binds to ALPPL2 with a dissociation constant of 20 nM, which falls in the low nanomolar range and is well suited for such purposes [34]. In terms of specificity, SQ2 is outstanding, as ALPPL2 is an ectopic protein and is expressed on pancreatic cancer cells and tissues, but not in normal pancreatic cells [35]. To ascertain the amount of SQ2 that enters the cells, we used RNase treatment and high salt wash, two well-established protocols, to study aptamer internalization. Removal of the surface-bound aptamers with RNase cocktail showed that 26% and 20% of the bound aptamer was internalized into Panc-1+ve and Capan-1 cells, respectively (Fig. 2A). No internalization was observed upon treatment with the mutant SQ2. Upon high salt wash, higher internalization of SQ2 was observed than obtained through the RNase treatment. Being a GPI-anchored protein, high salt wash may not be sufficient for the complete removal of the microdomain-associated membrane fractions of ALPPL2. In view of the other published aptamers that have been used for targeted delivery, the amount of SQ2 internalized by the cells was quite appreciable, especially considering that the target protein is not a membrane receptor. Next, we tested the cargo carrying capacity of the aptamer using biotinylated SQ2 and streptavidin-coated magnetic beads. The biotinylated aptamer was allowed to bind to the ALPPL2-expressing Panc-1+ve cells and was further probed with streptavidin magnetic beads. The beads were allowed to be internalized by incubating the cells at 37°C. Confocal microscopy indicated the presence of streptavidin magnetic beads inside the Panc-1+ve cells (Fig. 2B). No bead internalization was observed with the mutant SQ2. These magnetic beads were 2.8 μm in diameter, which suggests that SQ2 could efficiently mediate the internalization of nanoparticles and other large molecules.

SQ2 can be used as a tool for pancreatic cancer targeted therapeutics.
SQ2 aptamer delivers nucleoside drug 5FdU to ALPPL2+ve cells resulting in inhibition of cell growth
Cytotoxic nucleoside and nucleobase analogues, such as 5FdU, 5FU, and dFdC, are advantageous drugs for receptor-mediated active targeting, as they benefit from intracellular turnover. Recent studies on the aptamer-mediated delivery of cytotoxic nucleobase analogues employed a common strategy, wherein T7 RNA polymerase variant Y639F was used to incorporate the nucleoside drug analogue directly into the aptamer RNA backbone [36] or an RNA oligo tag that can be annealed to the aptamer through a complementary linker sequence extension [37]. Although this approach is quite economic, we found that the nucleoside-analogue-incorporated transcript had extremely low yields. Importantly, in the case of nucleoside drug incorporation into the aptamer sequence itself, the position of incorporation cannot be controlled and may consequently cause significant misfolding of the aptamer and reduction in the target binding affinity. In addition, as these nucleoside drugs are pyrimidine analogues, replacement of 2′F-modified pyrimidines may alter the nuclease-resistant properties of the aptamer.
We therefore designed a more defined targeting module, which would not alter the original properties of the aptamer and can be chemically synthesized and used directly for in vivo studies. The 3′-end of the truncated aptamer SQ2 was labeled with five repeats of 5FdU joined through a phosphorothioate backbone, as shown in Fig. 3A. Thus, one aptamer can deliver five monomer units of the drug, while the phosphorothioate backbone ensures that the drug is not cleaved extracellularly by the action of serum nucleases. We first confirmed that SQ2-5FdU was bound and internalized by Panc-1+ve cells with similar efficiency to that of the unmodified SQ2 (Fig. 3B). Next, normal pancreatic ductal cells, HPDE, and pancreatic cancer cells, Capan-1 and Panc-1+ve, were treated with SQ2-5FdU for 2 h. The mutant SQ2-5FdU and unmodified SQ2 were used as controls, and the effects were compared to that of free 5FdU. Although SQ2-5FdU is stable in 10% serum, the experiment was carried out in serum-free media to avoid potential nonspecific internalization due to the presence of serum. After 2 h of incubation, the cells were washed and the media was replaced with complete media. Cells were cultured further for 48 h to allow the drug to show its effects. 5FdU can be phosphorylated by thymidine kinase within the cell. The resulting 5-FdUMP, a strong thymidine synthase inhibitor, leads to a decline in cell proliferation, causing cell death. Cell viability was measured through the trypan blue dye exclusion assay. As shown in Fig. 3C, treatment with free 5FdU inhibited cell growth in both HPDE and Capan-1 cells, but the SQ2-mediated targeted delivery of 5FdU resulted in inhibition of cell growth only in Capan-1. SQ2-5FdU, at concentrations of 5 and 10 μM, inhibited Capan-1 cell growth by 40% and 52%, respectively. No significant inhibition was observed upon treatment with free drug or SQ2-5FdU at concentrations below 5 μM. Although some inhibition was also observed with mutant SQ2, this could be the result of nonspecific internalization through macropinocytosis, which is generally operative at high oligonucleotide concentrations [38,39]. In case of Panc-1+ve cells, even 1 mM of free 5FdU did not cause any effect on cell growth (data not shown). Therefore, the effects of SQ2-drug conjugates could not be studied on this cell line. Panc-1 insensitivity toward 5FdU and other prodrugs like gemcitabine has been reported by other groups as well [40,41].
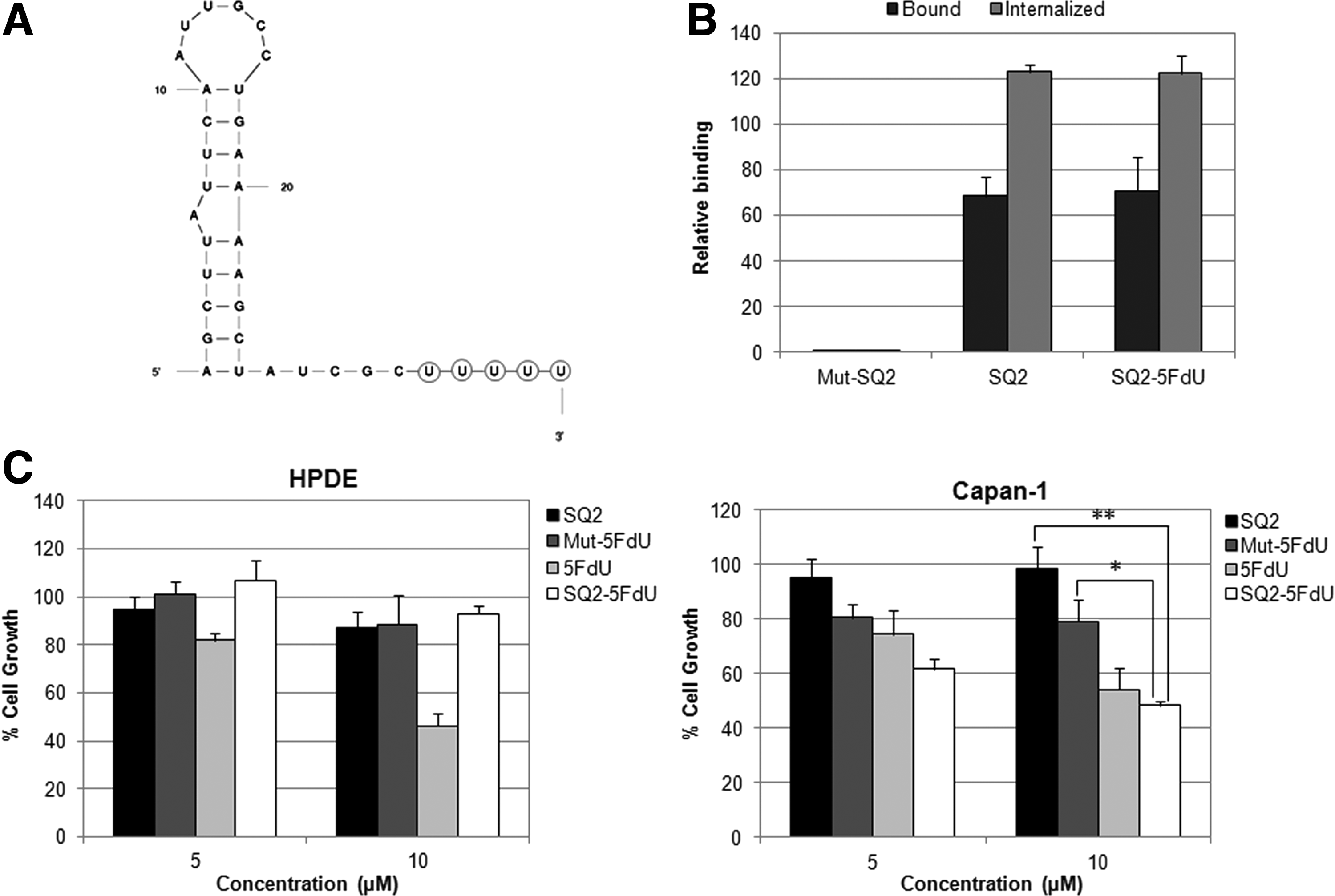
SQ2-5FdU inhibits Capan-1 cell growth but do not cause any effect in HPDE cells.
In conclusion, we demonstrated that the ALPPL2 aptamer SQ2 can be internalized into pancreatic cancer cells through clathrin-independent and caveolae-dependent or dynamin-mediated cell-type-dependent endocytic pathways and can be used for the targeted delivery of therapeutic molecules. SQ2 could successfully deliver the anticancer nucleoside drug 5FdU to ALPPL2-expressing pancreatic cancer cells but did not cause any toxicity to ALPPL2−ve normal pancreatic ductal cells. Thus, SQ2-mediated targeted delivery of 5FdU to pancreatic cancer cells could maximize the drug effectiveness, while concurrently minimizing the toxic side effects by reducing uptake into normal cells. In addition, this may also help in overcoming drug resistance associated with the cellular uptake of 5FdU. It should be noted that there is a lot of scope for improvement of the therapeutic index of this SQ2 targeted approach. For example, the length of the 5FdU homopolymer can be increased further or it can be replaced with more potent drugs. With an altogether different approach, SQ2-functionalized nanoparticle-mediated drug delivery can also be employed. This strategy would presumably be more efficient for increasing the intracellular drug concentration, as nanoparticle-mediated multimerization of aptamer would in turn lead to ALPPL2 aggregation and faster internalization. Finally, SQ2-mediated targeted delivery approaches are not limited to drugs alone but can also be applied for other therapeutic cargo, such as siRNA, radionucleotides, and toxins.
Footnotes
Acknowledgments
This work was supported by the Global Research Laboratory grant from the Ministry of Education, Science, and Technology of Korea (no. 2008-00582) and Agency for Defense Development through Chemical and Biological Defense Research Center (no. UD140017ID). P.D. was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF 2013R1A1A2062908). S.K. was supported by the Korea Ministry of Environment as “EI project” (ERL E211-41003-0007-0).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.