
Abstract
Successful short interfering RNA (siRNA)-based therapy for cancers depends on functional siRNA delivery specific to tumors. In our previous report, we have shown systemic siRNA delivery specific to human prostate cancer cell line PC-3 subcutaneous tumors in nude mice by atelocollagen, a collagen derivative, for formulating a complex with siRNA. We used an siRNA for human Bcl-xL as a model target. In the present study, we examined the antitumor effect on PC-3 orthotopic tumors in nude mice, as these tumors resemble the human clinical situation. The systemic intravenous administration of the complex (siRNA, 50 μg/shot) significantly reduced Bcl-xL expression and induced apoptosis in the tumors, and suppressed their growth. Liver metastasis was also inhibited in the orthotopic model. We successfully showed tumor-specific accumulation of the siRNA by Cy3-labeled siRNA and the direct quantification of the siRNA via reverse-phase high-performance liquid chromatography. The tumor-specific delivery was achieved by the enhanced permeability and retention effect, which is characteristic of macromolecular drugs. The high expression of vascular endothelial growth factor-A in the tumors provided adequate conditions to promote the permeability in the tumors, and to finally form the enhanced permeability and retention effect. In conclusion, our siRNA delivery is specific to the PC-3 orthotopic tumors in nude mice, and is practically feasible to treat tumors.
Introduction
F
The biomaterial atelocollagen, a type 1 collagen derivative from calf dermis, has the ability to bind with nucleic acid compounds via electrostatic interaction [9–11]. We have shown that atelocollagen can deliver many nucleic acids such as antisense oligodeoxynucleotides [12,13], morpholino antisense oligomers [14], and siRNAs [15,16] effectively into subcutaneously inoculated tumors in nude mice, along with the anticancer effects based on the specific knockdown of each target gene. In particular, an siRNA complexed with atelocollagen can be used as a nanoparticle-based medicine readily targeting tumor tissues [17], probably due to an enhanced permeability and retention effect (EPR effect). In that study [17], we used an siRNA targeting human Bcl-xL gene, an antiapoptotic factor that is overexpressed in prostate cancers, and we also used the human prostate cancer cell line PC-3 for subcutaneously xenografted tumors in nude mice. We formulated the siRNA with atelocollagen and intravenously administered it to the mice with PC-3 subcutaneously xenografted tumors in the lower flank. The complex was specifically delivered to the tumor tissues rather than the other normal tissues [17]. We obtained significant anticancer effects based on the specific knockdown of Bcl-xL gene [17]. Atelocollagen vehicle contributed to tumor specific delivery and uptake of the siRNA as well as maintenance on stability and bioavailability of the siRNA to prevent degradation in mouse body [15–18].
Successful studies showing antitumor effects via atelocollagen-mediated siRNA intravenous (i.v.) delivery are, however, limited in ectopically xenografted tumors under the skin of nude mice [17–20]. No reports of the therapy effect via siRNA–atelocollagen complex against orthotopically inoculated tumors have been published, to our knowledge. The orthotopic tumor model is ideal for more reliably studying the relationship between systemic siRNA delivery methods and their antitumor effects, because using the model, we can investigate the ability of the tumor microenvironment to affect and regulate our siRNA delivery method mediated by atelocollagen. The orthotopic tumors also exhibit more pathologic features similar to clinical human tumors compared to the ectopic tumors. In this regard, the orthotopic tumor models are commonly considered more valuable for the prediction of the clinical response to a therapeutic reagent [21].
In the present study, we found that the systemic i.v. injection of the complex of siRNA against human Bcl-xL gene and atelocollagen significantly reduced tumor enlargement, along with the induction of apoptosis in PC-3 orthotopic tumors. We also successfully showed the accumulation of the siRNA in the orthotopic tumors. Our siRNA delivery method significantly depended on the tumor permeability enhanced by vascular endothelial growth factor A (VEGFA) in the tumors.
Materials and Methods
Cell culture and reagents
PC-3 cells, a human prostate cancer cell line, were purchased from American Type Culture Collection (Manassas, VA) and cultured in Ham's F-12K (Kaighn's) Medium (F-12K medium, Invitrogen, Carlsbad, CA) with 10% heat-inactivated fetal bovine serum as described [15–17]. PC-3-Luc (JCRB1406) cells, which are PC-3 cells but with stable luciferase expression, were purchased from the Japanese Collection of Research Bioresources Cell Bank (Ibaraki, Japan), and cultured as described above. Cells were kept at 37°C in an incubator supplemented with 5% CO2. Cisplatin (CDDP) was purchased from Nippon Kayaku (Tokyo, Japan). Anti-human VEGFA neutralizing antibody (a monoclonal antibody raised in mouse) was purchased from LifeSpan BioScience (Seattle, WA).
siRNAs
In our earlier study [17], we described an siRNA targeting human Bcl-xL (called Bcl-xL siRNA) and a scramble siRNA (called Bcl-xL siRNA-SCR) as a control siRNA, which was designed according to the sequence of the Bcl-xL siRNA. Cy3-labeled Bcl-xL siRNA was also used for examining the delivery of the siRNA in vivo. All of the siRNAs were purchased from Dharmacon (Lafayette, CO).
Atelocollagen
Atelocollagen (Koken, Tokyo, Japan) was derived from tropocollagen of calf dermal collagen (type 1 collagen). To prepare atelocollagen, we removed telopeptides on both terminals via pepsin digestion to finally abolish the antigenicity [9–11]. Atelocollagen forms the complex with siRNA via an electrostatic interaction, resulting in a nanosized particle (100-300 nm) [18]. Atelocollagen is commercially available as a product, AteloGene®, from Koken.
Formulation procedure of the siRNA and atelocollagen complex
The complex was formulated as described [17,22,23]. In brief, an equal volume of siRNA in phosphate-buffered saline (PBS) and atelocollagen solution (0.1%) were mixed with each other at 4°C. We incubated the mixture by a gentle rotator machine for 20 min in a cold room and kept the complex at 4°C until injection. We prepared it at the time of use. The final concentration of atelocollagen for the systemic i.v. injection became 0.05%, as previously reported [17,22,23].
PC-3 orthotopic tumor model
All of the animal experiments were performed in compliance with the guidelines of the Institute for Laboratory Animal Research, Nagoya University Graduate School of Medicine. Male BALB/c athymic nude mice (9 weeks old, Japan SLC, Hamamatsu, Japan) were used. The mice were anesthetized with somnopentyl (intraperitoneal [i.p.], 5 mg/kg: Kyoritsu Seiyaku Corp., Tokyo) diluted in saline just prior to surgery. Following a lower midline abdominal incision, the prostate gland located underneath the bladder was exposed. PC-3 cells (5×105 cells in 20 μL of F-12K medium without any serum) were injected into the ventral lobe of the prostate [24,25]. We used an insulin syringe (Myjector 29G, Terumo Clinical Supply, Kakamigahara, Japan) with a fixed needle. The incision was then closed with 5-0 surgical sutures (Alfresa Holdings, Tokyo, Japan), and the mouse was then placed on a heat pad until awake.
Examination of distant metastasis from the PC-3 orthotopic primary tumors
PC-3 cells with stable expression of luciferase (PC-3-luc) were used to detect distant metastasis in vivo. We inoculated PC-3-luc cells into the prostate gland of nude mice as described in the previous section. Four weeks after the inoculation, all of the mice were sacrificed to remove the liver and lung. We homogenized the tissues using CelLytic MT Mammalian Tissue Lysis/Extraction Reagent (Sigma Aldrich, St. Louis, MO) with a Protease Inhibitors Cocktail (Sigma). The lysed samples were centrifuged and the supernatant was measured by the Dual-Luciferase Reporter Assay (Promega, Madison, WI). The values of luciferase activity (relative light units) were normalized to the total protein concentration determined by the Bio-Rad DC Protein Assay (Bio-Rad, Hercules, CA).
Cancer therapy via the systemic intravenous administration of the siRNA complex
Bcl-xL siRNA/atelocollagen complex was prepared following the protocol described in the above section. To evaluate the therapeutic efficacy of the complex, we intravenously injected the PC-3 orthotopic tumor-bearing mice with the complex (siRNA 50 μg in 200 μL, three consecutive injections per week) for 4 weeks. CDDP administration (2 mg/kg, single injection per week), was performed via an i.p. injection 1 day after the last injection of the siRNA for 4 weeks. The mice were sacrificed 28 days after the beginning of the administration, and the therapeutic effect was judged by determining the actual tumor weights.
RNA extraction, RT-PCR, and quantitative RT-PCR
Total RNA was isolated from PC-3 orthotopic tumors by an RNeasy mini kit (Qiagen, Hilden, Germany) to perform reverse transcription–polymerase chain reaction (RT-PCR) and quantitative RT-PCR. The total RNA (1 μg) was reverse-transcribed into cDNA by using an Omniscript RT Kit (Qiagen), and then PCR was performed with a Hotstar Taq Master Mix Kit (Qiagen). The primers and the PCR conditions for detecting human Bcl-xL were described [17]. Human glyceraldehyde 3-phosphate dehydrogenase was used for normalization. For the qRT-PCR, total RNA (800 ng) was reverse-transcribed by a High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Tokyo, Japan), and PCR was performed with a TaqMan probe specific to human Bcl-xL as per the supplier's instructions (Applied Biosystems). Hypoxanthine guanine phosphoribosyl transferase 1 was used for normalization [17].
Enzyme-linked immunosorbent assay specific to human Bcl-xL protein
Lysates from the excised tumor tissues were prepared as described [17]. Bcl-xL protein levels in the lysates were determined with the human Total Bcl-xL DuoSet IC ELISA Kit (R&D Systems, Minneapolis, MN). The total protein concentration measured with the Bio-Rad DC Protein Assay (Bio-Rad) was used for normalization [17].
Measurement of tumor permeability
A PC-3 orthotopic tumor-bearing nude mouse was injected i.p. with anti-human VEGFA neutralizing antibody (100 μg) or isotype-matched control immunoglobin G (IgG) (mouse IgG1, 100 μg). The injection volume was uniformly 100 μL. After the two injections over 2 days, we directly measured the resistance (Ohms) and current (Amperes) in the tumors when we loaded the voltage (70 V) in the tumors by our previously reported procedures using needle-type electrodes [26]. The summarized procedures with photographs are shown in Supplementary Fig. S1 (Supplementary Data are available online at www.liebertpub.com/nat). The tumor resistance and current in the tumors when electronically pulsed are a reliable index for measuring the tumor permeability [26].
Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling assay
Parts of the tumor tissues were snap-frozen into Tissue-Tek® O.C.T. compound (Sakura Finetek Japan, Tokyo, Japan) using liquid nitrogen and kept frozen in a deep freezer at −80°C until use [12,15–17]. Sections (4 μm thick) were cut with a Research Cryostat system (Leica CM3050S, Leica Japan) and fixed in acetone. After drying the acetone and washing with PBS, we obtained frozen sections for staining analysis.
We performed terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) staining for the frozen sections using a MEBSTAIN® Apoptosis Kit II according to the manufacturer's protocol (MBL, Nagoya, Japan). Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI) to determine the total cell numbers. TUNEL-positive cells were counted under a fluorescent microscope (Olympus) as reported [17].
Quantitation of Bcl-xL siRNA delivered in PC-3 orthotopic tumors and several other organs
The nude mice with orthotopic PC-3 tumors were i.v. injected with Bcl-xL siRNA/atelocollagen complex (siRNA, 50 μg/mouse). The mice were sacrificed at 15 min or 30 min post injection. The orthotopic PC-3 tumors and the other organs (liver, lung, kidney, spleen, and brain) were harvested, immediately immersed in RNAlater® solution (Ambion, Tokyo, Japan), and lysed by homogenization as described [17,22]. The lysates were hybridized with a fluorescence-labeled oligoribonucleotide probe (5′-GGUAUUGGUGAGUCGGAUC-3′), which is complementary to the antisense strand of Bcl-xL siRNA, and finally detected and quantified with reverse-phase high-performance liquid chromatography (HPLC) using the standard of the siRNA [17,22].
Administration of Cy3-labeled siRNA
We i.v. injected tumor-bearing mice with Cy3-labled Bcl-xL siRNA/atelocollagen complex (siRNA, 50 μg/mouse). The tumors were harvested 24 h after the injection, embedded in optimum cutting temperature compound, and frozen in liquid nitrogen. The frozen sections (3 μm) were cut and nuclear staining was performed with DAPI. Images of the sections were obtained with fluorescent microscopy (Olympus).
Histopathology and immunohistochemistry
The orthotopic PC-3 tumors were harvested and fixed in 4% paraformaldehyde in PBS overnight at 4°C. After dehydrating and clearing by ethanol and xylene, the tumor tissues were embedded in paraffin and then thin sections (6 μm) were made from the paraffin blocks. Hematoxylin and eosin (H&E) staining was performed as described elsewhere [22,23].
Statistical analysis
The statistical significance was examined by Mann-Whitney U-test; p values <0.05 were accepted as significant.
Results
Successful orthotopic tumor model inoculated with PC-3 cells
PC-3 cells were orthotopically inoculated into the prostate of BALB/c nude mice, and orthotopic tumors were successfully formed there 2 weeks after the inoculation. The PC-3 orthotopic tumors grew as time passed (Fig. 1A). A typical macroscopic view of the tumors 6 weeks after the inoculation is shown (Fig. 1B). The tumor formation in the prostate was pathologically confirmed by H&E staining (Fig. 1C). Finally, RT-PCR showed that human Bcl-xL gene was constantly expressed in the tumors at all of the time points examined up to six weeks (Fig. 1D). Quantitative real-time RT-PCR with a TaqMan probe for the same gene showed similar results (data not shown).

PC-3 orthotopic tumor model in nude mouse.
Determination of optimal Bcl-xL siRNA dose against orthotopic PC-3 tumors to significantly downregulate target gene
We have previously reported an siRNA targeting human Bcl-xL gene [17]. In the present study, three doses of Bcl-xL siRNA (25, 50, and 100 μg/injection) complexed with atelocollagen were intravenously injected into the tail vain of nude mice harboring PC-3 orthotopic tumors (Fig. 2A). These injections were repeated for three days. We examined all of the PC-3 orthotopic tumors to quantitate the expression levels of Bcl-xL (Fig. 2A). Both the qRT-PCR and the enzyme-linked immunosorbent assay, specific to human Bcl-xL, revealed a significant downregulation of the gene in the tumors (Fig. 2B, C). These suppressions reflected the dose-dependence of the siRNA injected. Accordingly, we found that the optimal dose of the siRNA was 50 μg/injection over 3 consecutive days.
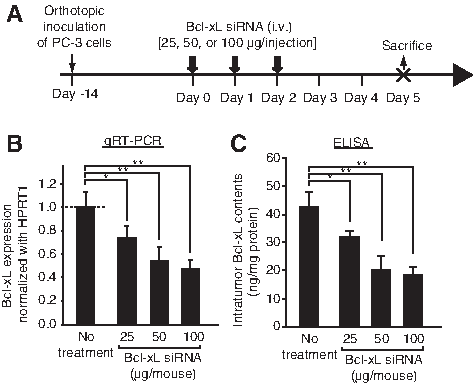
Examination of an optimal dose of the short interfering RNA (siRNA) to functionally downregulate human Bcl-xL expression on the PC-3 orthotopic tumors.
Significant antitumor effects of Bcl-xL siRNA complexed with atelocollagen on the orthotopic PC-3 tumors
The four sets of the three consecutive injections of the complex (Bcl-xL siRNA with atelocollagen; siRNA dose, 50 μg/injection) showed significant antitumor effects on the orthotopic PC-3 tumors compared with Bcl-xL siRNA-SCR with atelocollagen (p<0.01, Fig. 3A, B). Atelocollagen alone did not show any antitumor effects, which is consistent with our previous results [17]. Bcl-xL siRNA without atelocollagen as well as Bcl-xL siRNA-SCR without atelocollagen showed no effects (Fig. 3B). Combinational injections of a low dose of CDDP (2 mg/kg, i.p.) increased the antitumor effects (Fig. 3B) as shown previously in ectopic PC-3 tumors [17]. When we combine with Bcl-xL siRNA/atelocollagen complex and CDDP, the optimal injection timing of CDDP was also examined in our orthotopic tumor model (Supplementary Fig. S2). The timing, just after the three consecutive siRNA injections, showed stronger and significant antitumor effects, compared to the others (just before the three consecutive siRNA injections, or the intermediate timing between two groups of the three consecutive siRNA injections). These results suggested that just after the knockdown of Bcl-xL gene via siRNAs was the best timing to combinationally inject a chemotherapeutic, CDDP.

Significant antitumor effect of the siRNA complexed with atelocollagen on PC-3 orthotopic tumors.
The PC-3 orthotopic tumors frequently metastasized to livers and lungs, whereas the ectopically inoculated PC-3 tumors in the lower flank hardly metastasized anywhere (data not shown). Our siRNA-mediated therapy showed significant inhibition of the liver metastasis on day 28 as the end point (p<0.001, Fig. 3C). Bcl-xL siRNA/atelocollagen combined with CDDP (2 mg/kg) showed a significant and potent antimetastasis effect on livers (p<0.001) compared with the control (Fig. 3C). On lungs, although we observed such antimetastasis effect, unfortunately due to a large deviation of the control group, we did not obtain significant results (data not shown).
Bcl-xL siRNA treatment significantly increased apoptotic cell death in orthotopic tumors
At the end point of the therapy (day 28), we sacrificed all of the mice. The obtained orthotopic PC-3 tumors were subjected to the preparation of frozen sections and TUNEL staining. The tumors treated with Bcl-xL siRNA complexed with atelocollagen showed significantly increased TUNEL-positive cells on the section compared to Bcl-xL siRNA-SCR with atelocollagen (p<0.001, Fig. 4A, B). Without atelocollagen, almost no TUNEL-positive cells were observed in Bcl-xL siRNA. The combination of Bcl-xL siRNA with atelocollagen plus CDDP significantly increased TUNEL-positive cells (p<0.001).
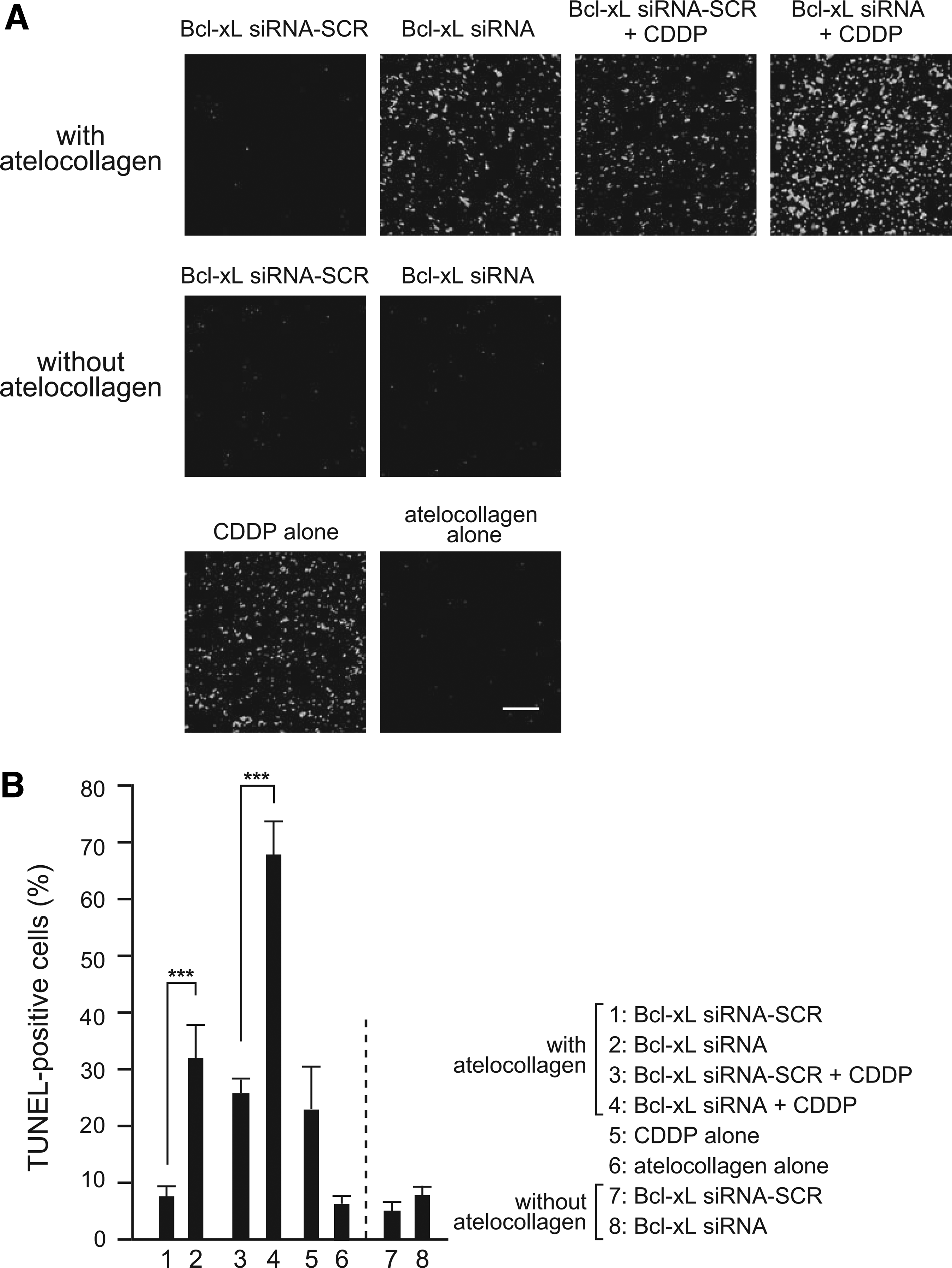
Significant apoptotic cell death induced by our therapy.
Atelocollagen mediated the siRNA delivery into the orthotopic PC-3 tumors
Bcl-xL siRNA labeled with a red fluorescent dye (Cy3) was complexed with atelocollagen, and intravenously injected into nude mice with orthotopic PC-3 tumors. The tumor sections from the injection group of Cy3-labeled Bcl-xL siRNA with atelocollagen showed potent red fluorescence, whereas the injection group without atelocollagen showed almost no fluorescence (Fig. 5). For all of the sections, the number of both red fluorescence-positive cells (siRNA-delivered cells) and DAPI-positive cells (total cells) was determined: the average transfection efficiency of the siRNA was 64% as in the case of complex with atelocollagen, and 3% without atelocollagen.
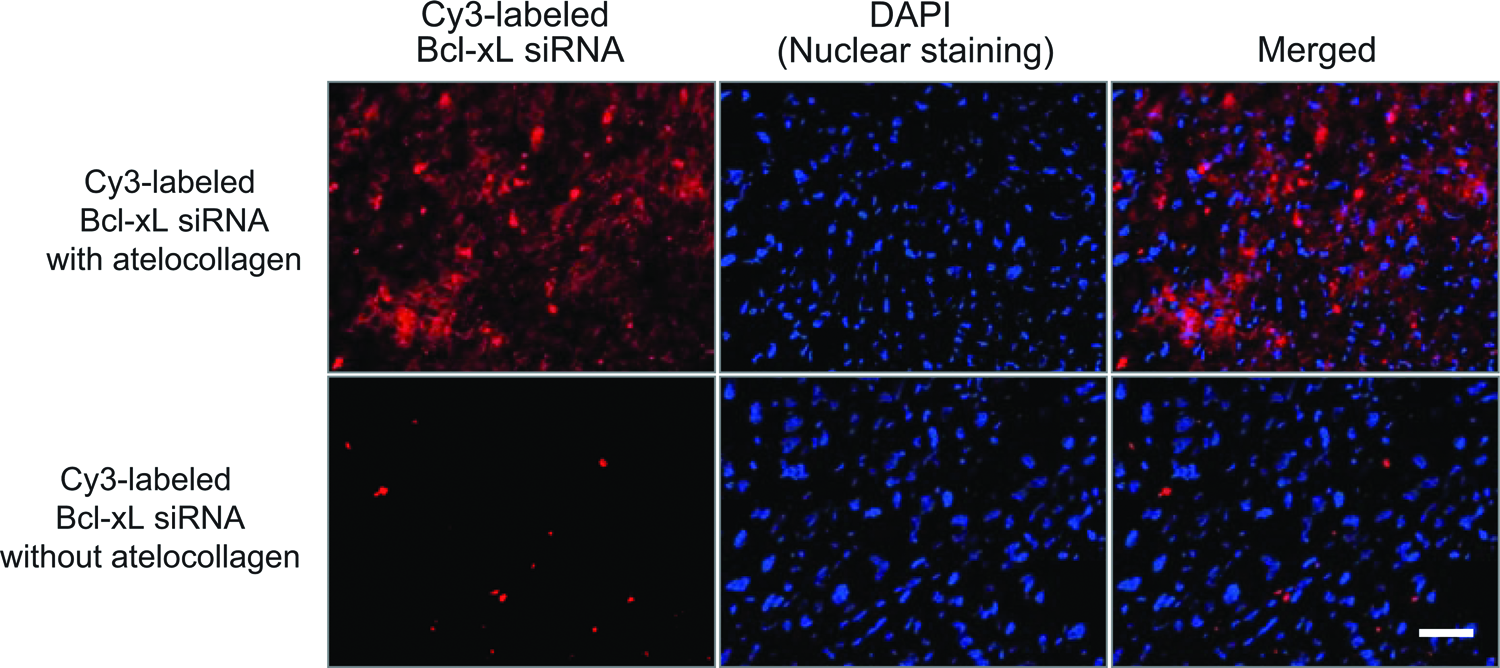
Cy3-labeled Bcl-xL siRNA complexed with atelocollagen was delivered to PC-3 orthotopic tumors. Frozen sections were made from the tumors after the i.v. injection of Cy3-labeled Bcl-xL siRNA (50 μg/injection) with (upper) or without (lower) atelocollagen. DAPI staining was also performed. All of the sections were photographed by fluorescence microscopy. Bar, 50 μm. Color images available online at www.liebertpub.com/nat
The siRNA complex was specifically delivered to the orthotopic PC-3 tumors
We quantitated the delivered siRNA amount into the orthotopic PC-3 tumors and normal tissues in nude mice as described earlier [17,22] and in the “Materials and Methods” section. Most of the Bcl-xL siRNA/atelocollagen administered via the i.v. route was accumulated in the orthotopic PC-3 tumors (Fig. 6A). The delivered amounts of the siRNA in the other normal tissues such as liver, lung, and kidney were quite small. In the spleen and brain, the siRNA was under the detectable level (Fig. 6A). The siRNA delivered into the tumors could be measured at least 30 min after the injection (Fig. 6B).

Tumor-specific delivery of Bcl-xL siRNA after i.v. administration mediated by atelocollagen.
Neutralizing anti-human VEGFA antibody impaired atelocollagen-mediated siRNA delivery
The siRNA/atelocollagen complex is a macromolecular compound classified as a nanomedicine that has an enhanced permeability and retention (EPR) effect [27,28] for targeting tumors. VEGFA is a major factor to maintain the enhanced permeability in many types of tumors [29,30], and it acts to induce the EPR effect. The PC-3 cells and their xenografted tumors in nude mice vigorously secreted VEGFA [15], and thus we attempted to inhibit the activity by anti-VEGFA neutralizing antibody. Figure 7A explains the experimental procedures. After the injection of the antibody, tumor permeability was reduced (Fig. 7B): compared with the control IgG group, the index of resistance of the tumors was significantly increased and the other index of current in the tumors was significantly decreased. The tumor microvessel density (tumor angiogenesis) determined by staining with anti-mouse CD31 antibody was not affected by the administration of the antibody in a short period (data not shown). Thus, the neutralizing antibody only reduced the permeability in the tumors. With these experiments, we showed the significant reduction of the RNA interference effect by the Bcl-xL siRNA in the anti-VEGFA neutralizing antibody group compared to the control IgG group (Fig. 7C, D).

High tumor permeability induced by vascular endothelial growth factor A (VEGFA) in the orthotopic tumors was essential for the atelocollagen-mediated siRNA delivery.
Discussion
Prostate cancer is a major malignant tumor and is the second leading cause of cancer related death for men in the United States [31]. Hormone ablation therapy is commonly adopted for early-stage androgen-dependent prostate cancer. However, prostate cancer often progresses to a more aggressive form and acquires resistance to any existing medical treatments including hormonal therapy, radiotherapy and chemotherapy [32,33]. Indeed, in clinical practice, castration-resistant prostate cancer (CRPC) patients are often treated with microtubule inhibitors such as docetaxel [34] or cabazitaxel [35], but these therapeutic compounds have shown only limited effects, along with occasionally severe adverse effects. The resistance of CRPC to various drugs is attributed mainly to its genetic and epigenetic mutations in intracellular signaling pathways [36]. Thus to date, we have almost no functional therapy ideas to combat CRPC. Accordingly, therapy strategies based on the biological features of CRPC are urgently required.
Bcl-xL, which belongs to the Bcl-2 family, is an antiapoptotic factor on mitochondria [37] and is also highly expressed in CRPC [38]. The elevated Bcl-xL in CRPC causes these cancers' resistance to apoptosis [38–40]. In earlier studies, the functional inhibition of Bcl-xL resulted in the apoptosis of prostate cancer cells including human androgen-nonresponsive prostate cancer cells (i.e., PC-3 cells) [17,38–40], suggesting that Bcl-xL is an adequate target for treating CRPC. In the present study, using the siRNA for targeting human Bcl-xL gene and atelocollagen systemic siRNA delivery techniques, we observed significant antitumor effects against PC-3 orthotopic tumors (Fig. 3), suggesting the feasibility of these techniques to treat CRPC.
Throughout the course of the therapy, we observed no liver damage as assessed using aspartate aminotransferase and alanine transaminase or renal damage as assessed by creatinine and blood urea nitrogen (data not shown). No severe loss of body weight in the treated mice was also observed. Interferon-induction effect, which is a kind of adverse action caused by double-stranded RNAs including siRNAs, was not observed in our atelocollagen-mediated siRNA therapy, consistent with our previous studies [17,22]. Taken together, our findings indicate that our proposed therapy method is safe to treat cancers.
When the animals were treated with siRNA formulated with atelocollagen and CDDP at the same time, the antitumor effect was enhanced without any additional adverse effects, compared to the groups treated with either of them. This result suggests that the combination of chemotherapy and our Bcl-xL siRNA-based therapy could provide an advanced cancer therapy method attaining both efficacy and safety at a high level, which would be appropriate even for patients suffering from CRPC.
One of the major concerns about cancer therapy is how to combat against metastasis. In the present study, we obtained a proper animal model to examine the inhibition of metastatis: The PC-3 orthotopically inoculated tumor model but not the PC-3 subcutaneously inoculated tumor model [17] yielded efficient cancer metastasis from prostate to liver. Our therapy method demonstrated significant antimetastasis effects in the orthotopic model, although a simple treatment with CDDP never achieved such sufficient inhibition (Fig. 3). Thus, our siRNA-based therapy using atelocollagen as a delivery vehicle showed superior potency from four different viewpoints: antitumor effect, antimetastatic effect, safety, and potential for combination with chemotherapeutics.
Next, we investigated how our therapy works in the body of animals. As one strong possibility, we devoted our attention to the theory of EPR effect, which enables macromolecule delivery specific to tumors with high permeability due to leaky and incomplete vessels [28,41]. We found that the atelocollagen-mediated siRNA delivery method was PC-3 orthotopic tumor-specific by investigating the direct quantification of the siRNA via reverse-phase HPLC (Fig. 6). The EPR effect was essential to achieve our atelocollagen-mediated siRNA i.v. delivery (Fig. 7). The abundant VEGFA expression in the PC-3 orthotopic tumors contributed to the upregulation of tumor permeability [26,29–30] and to the EPR effect. Indeed, the i.p. administration of anti-human VEGFA-neutralizing antibody impaired the RNA interference efficacy via the atelocollagen-mediated siRNA i.v. delivery (Fig. 7).
What is the molecular mechanism underlying the ability of the siRNA/atelocollagen complex accumulated in tumor tissues via the EPR effect to finally penetrate into tumor cells? The EPR effect must be essential for our atelocollagen-mediated siRNA-based therapy; however, it is not enough to explain the delivery mechanism. We have been studying it, especially focusing on a molecule, Endo180 (also known as macrophage mannose receptor C type 2, urokinase plasminogen activator receptor-associated protein, or CD280 protein), on the cellular membrane [42,43]. Endo180 has an ability to bind with type 1 collagen on the extracellular domain, and Endo180 takes the type 1 collagen into the cells [43]. In breast and pancreatic cancers, Endo180 is overexpressed and promotes the tumor growth in vivo [44,45]. Further, we found Endo 180 is also overexpressed in prostate cancers (Takei Y, unpublished results). We have already obtained several promising data to support our hypothesis and will describe the results of our further experiments elsewhere in the future.
In conclusion, our atelocollagen-mediated siRNA delivery is specific to the PC-3 orthotopic tumors in nude mice and is feasible for the treatment of tumors.
Footnotes
Acknowledgments
We thank Drs. Kazuo Kita, Takayuki Okubo, and Koji Hanai for their helpful suggestions regarding the experiments and manuscript. We also thank Dr. Guodong Shen, Dr. Naomi Ohnishi, Ms. Naomi Maruyama, and Ms. Akiko Suzuki for their excellent technical assistance, and Koken Co., Ltd. for generously providing atelocollagen.
This work was supported in part by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology of Japan (17016030) and from the Japan Society for the Promotion of Science (21590305 and 24590348).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.