
Abstract
Most therapeutical nucleic acid aptamers tend to inhibit protein–protein interactions and thereby function as antagonists. Attachment of the influenza virus surface glycoprotein hemagglutinin (HA) to sialic acid-containing host cell receptors (glycan) facilitates the initial stage of viral infection. Inhibition of the attachment may result in an antiviral effect on the proliferation of the influenza virus. To develop therapeutically interesting agents, we selected two single-stranded DNA (ssDNA) aptamers specific to the HA protein of H1N1 influenza virus (A/Puerto Rico/8/1934) through a procedure of systematic evolution of ligands by exponential enrichment. As it showed a higher binding affinity for HA protein (Kd = 78 ± 1 nM), aptamer 1 was tested for its ability to interfere with HA-glycan interactions using chicken red blood cell hemagglutination and microneutralization assays, which demonstrated that it significantly suppressed the viral infection in host cells. These results indicate that the isolated ssDNA aptamer may be developed as an antiviral agent against influenza through appropriate therapeutic formulation.
Introduction
A
Due to a variety of favorable characteristics, aptamers have been used in numerous applications such as identification of biomarkers, detection of pathogens, and development of new drugs [6–9]. They can be used for therapeutic purposes in much the same way as monoclonal antibodies. Moreover, there are several potential advantages of aptamers over antibodies. As they are much smaller in size than antibodies, aptamers are better candidates for cell penetration, blood clearance, and chemical modifications. In addition, they are readily synthesized and nonimmunogenic and therefore they do not cause side effects resulting from unwanted immune responses in hosts [10].
So far, researchers have identified high-affinity aptamers of therapeutic interest that target a broad cross section of protein families, including cytokines, proteases, kinases, cell surface receptors, and cell adhesion molecules [11]. Theoretically, aptamers can be used therapeutically in any disease for which extracellular blocking of protein–protein interactions is required. Many aptamers that are selected to bind to a specific protein may also inhibit its function. Several high-affinity DNA and RNA aptamers have so far been successfully applied in suppressing the function of viral proteins, such as human immunodeficiency virus HIV glycoprotein 120 (gp120), human hepatitis B virus polymerase (P protein), and influenza virus NS1 protein [12–14].
Influenza viruses are RNA viruses that make up three of the five genera of the family Orthomyxoviridae: influenza viruses A, B, and C. Among these types, the influenza A virus leads to seasonal epidemics and sporadic pandemics of influenza and hence it is considered the major cause of this acute infectious disease and death. Influenza A viruses are classified into 16 HA (H1-H16) and 9 NA (N1-N9) subtypes, of which H1-H3 have successfully adapted to humans [15].
Influenza virus particles contain two virus-encoded surface glycoproteins, the hemagglutinin (HA) and the neuraminidase (NA). These two surface glycoproteins are important antigens that determine antigenic variation of influenza viruses and host immunity. Variability in HA is primarily responsible for the continual evolution of new strains and subsequent influenza epidemics [16]. The HA protein of influenza virus, a trimeric spike on the viral membrane, is responsible for hemagglutination and binding of the virus particles to the susceptible host cells. This binding mediates the subsequent entry of influenza viruses into host cells through membrane fusion, which is significant for initial viral infection [17,18]. The HA protein may be cleaved proteolytically by host enzymes into HA1 and HA2 subunits that remain tightly linked by a disulfide bond. The cleavage of HA into HAl and HA2 is necessary for the virus to be infectious. HA1 has receptor-binding domains for sialic acid receptors located on the surface of host cells and hence forms a complex with host cell surface glycans through a terminal sialic acid by α2-3 and α2-6 linkages. This binding is essential for the infection, transmission, and virulence of influenza viruses [19]. The amino terminal of HA2, generated by the cleavage event, is necessary for the viral envelope to fuse with the cell membrane, an essential step in the process of viral infection. Accordingly, HA may be used as an important target for the therapeutic depression of its function as well as a biomarker for detection of the virus.
Influenza spreads around the world in a yearly outbreak, resulting in about 3–5 million cases of severe illness and about 250,000–500,000 deaths. In the past 100 years, large outbreaks known as pandemics occurred in 1918, 1958, and 1968, each causing more than a million deaths. An outbreak of a new type of influenza, A/H1N1, in 2009 also caused several hundred thousand deaths. It is quite clear that influenza epidemics and pandemics are great and constant threats to public health. In recent years, the increasing prevalence of drug-resistant viruses has greatly contributed to the increased difficulties in the control and treatment of influenza and become a major public health concern, highlighting the urgent need for the development of alternative therapeutic approaches.
Although several high-affinity DNA and RNA aptamers have been successfully applied in suppressing the function of viral proteins [20–23], few studies have been reported on the aptamers used for H1N1 (A/Puerto Rico/8/1934) influenza virus suppression. Hence, we developed a DNA aptamer that specifically binds to the HA protein of the H1N1 virus and investigated its antiviral activity in host cells. Our results showed that this aptamer could efficiently inhibit the HA-glycan interactions, suggesting that this aptamer has potential applications in the development of H1N1-specific virucidal agents.
Materials and Methods
Materials
All DNA oligonucleotides, that is, the initial ssDNA library, the aptamers with known sequences, and the forward and the reverse primers, were synthesized by Shanghai Sangon Biological Engineering Technology & Services Co., Ltd. (Shanghai, China). Madin-Darby canine kidney (MDCK) cells and influenza virus A/Puerto Rico/8/1934(H1N1) were maintained in Molecular Virology Laboratory of Hunan Normal University. Dulbecco's modified Eagle's medium (DMEM) and fetal bovine serum (FBS) were purchased from HyClone (https://promo.gelifesciences.com/gl/hyclone). The recombinant HA protein of avian influenza virus (AIV) subtype H1N1 (A/Puerto Rico/8/1934) with a concentration of 0.25 mg/mL was offered by Sino Biological, Inc. (Beijing, China). DNA markers, DNA recovery Kit, polymerase chain reaction (PCR) Kit, phosphate buffer saline (PBS), and trypsin were purchased from Shanghai Sangon Biological Engineering Technology & Services Co., Ltd. Yeast tRNA and bovine serum albumin (BSA) were obtained from Brother Biological Science and Technology Co., Ltd. (Changsha, China). All the other chemicals were analytical reagent grades and were used without further purification. The 96-well microtiter plates were purchased from Gelin NanoTech Co., Ltd. (Changsha, China). All materials used in the experiment were dissolved in distilled water purified by a Milli-Q water purification system (electric resistivity 18 MΩ cm−1). The UV spectrophotometer used for absorbance detection at 260 nm was purchased from Tianmei Techcomp (Shanghai, China). The fluorescent emission spectra were recorded in a quartz cuvette by an LS55 luminescence spectrometer (PerkinElmer, Wokingham, UK).
Methods
In vitro selection of ssDNA aptamers against HA protein
A synthetic ssDNA library containing a central random sequence of 35 nucleotides (nt) flanked by primer hybridization sites of 23 nt at the 5′ end and 20 nt at the 3′ end (5′-GGGAGCTCAGAATAAACGCTCAA-N35-TTCGACATGAGGCCCGGATC-3′) was used for SELEX. A 96-well microtiter plate was coated with HA protein dissolved in 200 μL of 0.05 M NaHCO3 solution (pH 9.6) at 4°C overnight, followed by an addition of 200 μL of 3% BSA at 37°C for 2 h to block the wells coated with HA. The ssDNA library was dissolved in 200 μL of binding buffer (20 mM HEPES buffer pH 7.35, 120 mM NaCl, 1 mM MgCl2, 1 mM CaCl2, and 5 mM KCl) blended with yeast tRNA for the purpose of decreasing background binding. The library was denatured at 95°C for 5 min, cooled immediately in ice water for 10 min, and transferred to BSA-blocked blank wells and maintained at 37°C for 40 min. The uncombined ssDNAs were subsequently transferred to the wells coated with HA protein at 37°C for 40 min. The HA-coated wells were washed five times with washing buffer (binding buffer + 0.05% Tween 20) and then filled with eluting buffer (20 mM Tris-HCl, 4 M guanidinium isothiocyanate, 1 mM DTT, pH 8.3) at 80°C for 10 min. The solution (counted as one volume) was subsequently transferred to a clean EP tube and three volumes of dehydrated ethanol and one-tenth volume of 3 M sodium acetate (pH5.2) were added and kept at −80°C for 40 min and centrifuged at 30,857 g for 10 min at 4°C. The sediment was washed twice by absolute ethyl alcohol and dried and dissolved in 20 μL double-distilled water.
The ssDNA was amplified by PCR with the following protocol: PCR system included 0.1 μg template, 2 μL 10× PCR buffer, 1.5 mM MgCl2, 200 μM dNTP, forward and reverse primers with 0.5 μM for each, Taq DNA polymerase 1 U, addition of sterilized distilled water to a total reaction volume of 20 μL; 20 cycles of 5 min at 95°C, then 0.5 min at 95°C, 0.5 min at 65°C, and 0.5 min at 72°C, followed by 5 min at 72°C. The forward and reverse primers were 5′-GGGAGCTCAGAATAAACGCTCAA-3′ and 5′-GATCCGGGCCTCATGTCGAA-3′, respectively. The asymmetric PCR (template 0.1 μg, 10 × PCR buffer 2 μL, MgCl2 0.75 mM, dNTP 200 μM, forward primers 20 μM and reverse primers 0.5 μM, Taq DNA polymerase 1 U, addition of sterilized distilled water to a total reaction volume of 20 μL; 40 cycles of 5 min at 95°C, then 0.5 min at 95°C, 0.5 min at 65°C, and 0.5 min at 72°C, followed by 5 min at 72°C) was used to generate the ssDNA as an enriched library for the next round of SELEX procedure. The amount of ssDNAs at each round of selection was measured by a UV-Vis spectrophotometer. The aptamer binding ratio was obtained by calculating the percentage of the eluted ssDNA amount to the total amount of ssDNA invested to this round of selection. An increasing stringency of selection was achieved by the use of increasing amounts of yeast tRNA and decreasing amounts of ssDNA and HA protein while the selection cycles proceeded (Table 1).
HA, hemagglutinin; SELEX, systematic evolution of ligands by exponential enrichment; ssDNA, single-stranded DNA.
After 13 rounds of enrichment, the PCR products selected from the 13th round were subcloned and sequenced by Shanghai Sangon Biological Engineering Technology & Services Co., Ltd. The secondary structures of the selected aptamer were predicted using the DNAMAN software. The dissociation constant was calculated by the use of the equation, Y = BmaxX/(Kd+X), where Kd is the dissociation constant, Bmax is the maximal fluorescent intensity measured by this experiment, Y represents the mean fluorescent intensity, and X is the concentration of the aptamer.
Preparation of virus and cells
The virus was cultivated in the allantoic cavities of 10-day-old embryonated chicken eggs and maintained at 35°C. After 72 h of culturing, the allantoic fluid was harvested and centrifuged. The supernatant was collected and stored at −70°C before use. The titer of virus used for infection was evaluated by the infection of MDCK cells [24] and hence virus titer was expressed as the tissue culture infective doses leading to 50% infected cells (TCID50). MDCK cells were maintained in our laboratory in DMEM supplemented with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin. Chicken red blood cells (RBCs) were recovered by means of centrifugation. The recovered RBCs were washed four times and suspended in physiological saline, with a concentration kept at 0.5% (v/v).
Hemagglutination inhibition assay
The ability of the selected aptamer 1 to interfere with the agglutination of chicken RBCs was evaluated with an agglutination inhibition assay. The assay was conducted on a round-bottomed 96-well plate. Fifty microliters of PBS buffer (pH 7.4) and 50 μL of 500 TCID50A/Puerto Rico/8/1934 virus were mixed and added to each well. The serially diluted recombinant HA aptamer 1 and Salmonella Paratyphi A aptamer with binding buffer were denatured at 95°C for 5 min and cooled in ice water for 10 min. These aptamers were then added into the wells with different concentrations (500–12,000 nM) and incubated with virus mixtures at room temperature for 40 min before chicken RBCs of 100 μL were added to each well and incubated at room temperature. All agglutination reactions were allowed to last for 30 min before photographing. PBS was set as a negative control.
Microneutralization assay
The antiviral effect of the selected aptamer 1 was further determined by using the microneutralization assay [25]. In this procedure, MDCK cells were cultured for 24 h in a 96-well plate (70,000/well). The MDCK cells were infected by a mixture containing 1,000 TCID50 (100 μL) of A/Puerto Rico/8/1934 virus treated with serially diluted aptamers (0–10,000 nM) and incubated in a 5% CO2 humidified atmosphere at 37°C for 1 h. Subsequently, the liquid cultures were removed. The cells were washed with PBS once and incubated in DMEM supplemented with 2% fetal calf serum under the same condition for 72 h. A Salmonella Paratyphi A aptamer (10,000 nM) instead of aptamer 1 was set as a control and treated similarly in parallel. Viral titers of the culture media were measured by the hemagglutination assay similar to the abovementioned procedure. Briefly, 100 μL of serially diluted culture media (20–26-fold, diluted with PBS) was added into a microtiter plate, followed by an addition of 100 μL of 0.5% chicken RBCs. All experiments were conducted in triplicates in 96-well microtiter plates.
Results
Selection of ssDNA aptamers against HA protein
The selection of ssDNA aptamers against HA was performed using a regular SELEX protocol. The procedure used a commercially available target, the recombinant HA protein of AIV subtype H1N1 (A/Puerto Rico/8/1934). As the cycles of selection proceeded, an increasing stringency of selection was introduced by increasing the amounts of BSA and yeast tRNA accompanied by decreasing amounts of ssDNA and HA protein (Table 1). Such modification increased the competition between the desired target (HA proteins) and nontarget (BSA molecules) for aptamer molecules, and the tRNA was used to compete with the aptamer sequences for target binding sites. On the other hand, the use of decreasing amounts of ssDNA and HA protein also reinforced the competition and promoted the specific binding between HA proteins and the aptamer molecules. Enrichment of specific aptamers was determined during the selection process. The percentages of bound ssDNA against HA protein were observed.
As shown in Fig. 1, the aptamer binding ratio began to increase at round 5 as 14%. At round 9 and round 11, the binding ratios were 23% and 36%, respectively. However, the affinity of the ssDNA pool to HA protein appeared to be nearly constant after round 11, indicating that the binding of the ssDNA pool to HA protein became saturated. After 13 rounds of selection, the aptamers showed strong affinities for HA protein. Therefore, the ssDNA molecules selected from the 13th round were cloned and sequenced. Two independent candidates were obtained and named aptamer 1 and aptamer 2. Their secondary structures were also predicted by DNAMAN software. As shown in Fig. 2, both aptamers contained a number of stem-and-loop structures. Conserved sequences (GGCAC) were also found residing at the stem structure flanking loop region. As it showed higher binding affinity for HA protein (Kd = 78 ± 1 nM) in comparison with aptamer 2, aptamer 1 was selected for further experiments.
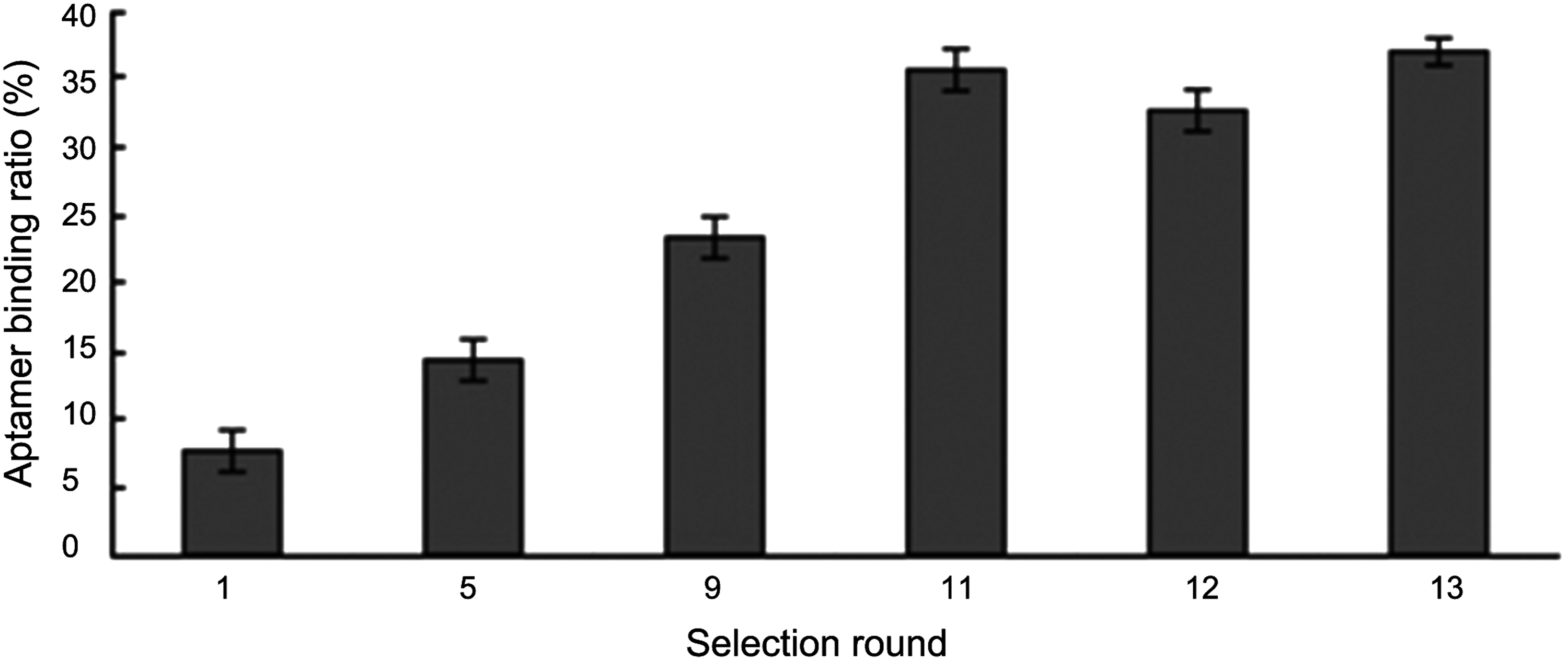
Enrichment of HA protein-specific aptamers during the selection cycles. The bar graph reveals the ratio of the amount of ssDNA eluted from 96-well plates coated with HA proteins to the total amount of ssDNA used in this round of selection. HA, hemagglutinin; ssDNA, single-stranded DNA.

Predicted secondary structure of the selected ssDNA aptamers 1 and 2 using DNAMAN software. The underlined letters represent the central random 35 nucleotides. The conserved sequence in each aptamer is indicated by shading on each structure.
Hemagglutination inhibition assay
Hemagglutination inhibition assay was conducted to evaluate the inhibitory activity of the ssDNA aptamer against viral HA protein. The assay was performed with the process of hemagglutination, in which the viral HA proteins were bound to the sialic acid receptors on the RBC surface and created a lattice structure of interconnected RBCs and virus particles, and so the hemagglutination was achieved. The agglutinated RBCs were maintained in a suspended distribution and viewed as a diffuse reddish solution. However, when the binding of HA proteins to the sialic acid receptors was blocked by inhibitors, such as aptamers, no hemagglutination would occur and the RBCs were not constrained by the lattice and should settle to the bottom of the well. To confirm the inhibition effect of the selected aptamer on the binding of the virus to RBCs, we cultivated the virus in the allantoic cavities of 10-day-old embryonated chicken eggs. The allantoic fluid was harvested, and the titer of virus in the supernatant was evaluated and expressed as TCID50. The inhibitory potency of aptamer 1 was measured by hemagglutination inhibition assay. RBCs may be agglutinated if no aptamer is added into the system. With the addition of a specific aptamer, however, no agglutination would be observed.
As shown in Fig. 3, a top-down view of the hemagglutination reaction mixtures was displayed in a round-bottomed microwell plate after the addition of aptamer 1 (Fig. 3A). Clear agglutination was observed in wells containing 0, 500, 1,000, and 2,000 nM of aptamer 1, and partial agglutination was observed in the cases of 4,000, 6,000, and 8,000 nM. At the concentration of aptmer 1 higher than 10,000 nM, a sharp button of RBCs at the bottom of the microtiter well was observed. This suggested that aptamer 1 exerted a significant inhibition effect on the binding of the influenza viruses to the RBCs. In contrast, the hemagglutination reaction was observed at all concentrations when the unspecific Salmonella Paratyphi A aptamer was mixed (Fig. 3B), indicating that the unspecific aptamer exhibited no inhibition effect.

Inhibition of chicken RBC agglutination by HA aptamer 1. The serially diluted HA aptamer 1
Microneutralization assay
The antiviral activity of the selected aptamer 1 was further confirmed by microneutralization assay. MDCK cells were infected with a virus solution containing serially diluted DNA aptamers, followed by incubation at 37°C for 1 h. The liquid culture media were then removed so as to remove the unbound viruses. After renewal of the culture media, incubation was maintained for an additional 72 h to let the virus grow. Viral titers of the culture solutions were then measured by hemagglutination assay in the presence of 0.5% chicken RBCs.
As shown in Fig. 4, agglutination in the 0 nM aptamer group was not abolished until the dilution times of culture solution reached 25-fold. Agglutination in the 6,000 nM group, however, appeared to be inhibited when the culture solution was only doubly diluted. In addition, more evident results of agglutination inhibition could be seen in the 8,000 nM and 10,000 nM groups. Notably, the 10,000 nM Salmonella Paratyphi A aptamer group showed a very similar agglutination pattern to that of the 0 nM aptamer 1 group, suggesting that the control aptamer did not exert a noticeable antiviral effect. These results proved that aptamer 1 exhibited a significant inhibition of viral infection.

The antiviral activity of the selected HA aptamer 1. Madin-Darby canine kidney cells were infected with 1,000 TCID50 H1N1 (A/Puerto Rico/8/1934) virus solution containing serially diluted HA aptamer 1 (0–10,000 nM) through incubation at 37°C for 1 h. After renewal of culture media, the incubation continued and lasted for 72 h. Viral titers of the serially diluted culture media (20–26-fold) were determined by the hemagglutination assay in the presence of 0.5% chicken RBCs. The unspecific Salmonella Paratyphi A aptamer (10,000 nM) was used as a control. Negative control was performed with PBS.
Discussion
In this study, two ssDNA aptamers against HA protein of H1N1 influenza virus with high affinity were obtained after 13 rounds of iterative SELEX cycles. Their secondary structures were predicted by DNAMAN software. We found that each of the two selected aptamers contains a short conserved sequence residing at the stem structure flanking loop region (Fig. 2). The conserved sequence appeared to be an important structure for the binding of an aptamer to its target. It has been demonstrated that some RNA aptamers often contain a conserved sequence of several nucleotides, most of them reside in the loop structure or in the stem-and-loop region [26,27]. Park et al. believed that the conserved sequences in RNA aptamers are exposed, which facilitate interactions with the targets, and the conserved sequences residing at the RNA aptamer stem-and-loop region could constitute a binding motif structure to the HA1 subunit of HA. Similarly, the conserved sequence residing at the stem-and-loop structures of the two DNA aptamers in this study may represent the active binding sites. Aptamers often form secondary structures when bound to their targets. Apart from stems and loops, hairpins, pseudoknots, and G-quadruplexes are also present, with a different structure probably binding to a different epitope of the target.
As aptamer 1 showed higher binding affinity for HA protein, its antiviral effect was evaluated. The hemagglutination inhibition assay proved that aptamer 1 against HA of the influenza virus, H1N1, exhibited a significant inhibition effect on the virus-induced hemagglutination of chicken RBCs. Further confirmation of the reactivity of aptamer 1 with the influenza virus was also conducted with microneutralization assay, which demonstrated that aptamer 1 decreased viral titers of the host cell culture media and exhibited an efficient inhibition effect on viral proliferation.
As mentioned above, the HA protein of influenza virus is responsible for hemagglutination and binding of the viral particles to the susceptible host cells and hence mediates the subsequent entry of influenza viruses into host cells through membrane fusion, which is significant for initial viral infection. The binding of aptamer 1 to the HA protein is critical for the inhibition of HA-receptor (glycan) interactions. Several results suggest that aptamer 1 in this study can recognize and bind to the HA protein. Recently, we have developed a label-free aptasensor for rapid detection of H1N1 virus by using a DNA hairpin probe containing this aptamer 1 [28]. The binding of HA protein to the probe triggered the release of probe from grapheme oxide and so the Klenow fragment of DNA polymerase lengthened the probe to a longer ssDNA product, resulting in a strong increase in the fluorescence intensity when the SYBER Green I was introduced. The binding of aptamer 1-containing probe to HA protein was further demonstrated by electrophoresis, in which the sample of HA-aptamer 1 complex exhibited a different electrophoretic behavior in contrast with the probe or the HA protein used as a sample alone (data not shown). The reactivity of aptamer 1 with influenza virus to inhibit the virus-induced hemagglutination also lends support to the binding of aptamer 1 to the HA protein. According to Park, the mere binding between them is not enough to guarantee the suppression of hemagglutination activity. Only by the tight binding to the critical site of HA protein can aptamer 1 exhibit hemagglutination inhibition activity. Therefore, it may be a competent candidate for detection of the critical binding site of HA protein. As HA is known to be cleaved into HA1 and HA2 subunits, of which HA1 mediates initial contact with the cell membrane [29], aptamer 1 is presumably bound to the HA1 subunit. Further analysis is necessary to elucidate their exact binding sites.
At present, the influenza treatment relies largely on two classes of conventional antiviral drugs, the M2 ion channel blockers and the NA inhibitors [30]. Because of the ever-increasing viral drug resistance, the efficacies of all these drugs are compromised. The possibility of targeting virus–host interaction and host cellular mechanism to develop new treatment regimens has attracted much attention. Many novel classes of antiviral compounds or biomolecules targeting the viral replication mechanism are therefore under development. They focused mainly on the HA-glycan interactions, and a number of DNA and RNA aptamers against influenza viruses, H1N1, H2N2, H3N2, H5N1, H5N2, H7N7, H9N1, and H9N2, and influenza B virus have been developed, of which over a dozen have been developed for therapeutic purposes and almost all of them target HA [31]. They are capable of inhibiting the HA capacity of the virus and preventing the viral infection in cell culture or in animal models. Generally, they exert such effects at nanomolar to micromolar ranges. The wide variations of aptamer dosage could be due to differences in the HA affinities to the glycans among different influenza subtypes. In the present study, aptamer 1 that we isolated well inhibits the hemagglutination or viral infection to the cultured MDCK cells at micromolar ranges. Unlike RNA aptamers, aptamer 1 is more stable to nucleases. As few studies have so far been reported on the aptamers used for H1N1 (A/Puerto Rico/8/1934) influenza virus suppression, we believe that aptamer 1 can be a promising candidate for the development of antiviral agent.
In conclusion, two ssDNA aptamers were obtained by using a recombinant HA protein of AIV subtype H1N1 (A/Puerto Rico/8/1934) as a target, of which aptamer 1 was tested and proved to have an efficient antiviral activity by interfering with HA-glycan interactions. This selected aptamer therefore has potential applications in the development of H1N1-specific virucidal agents.
Footnotes
Acknowledgments
The authors would like to thank the National Natural Science Foundation (81271660), the Specialized Research Fund from the Ministry of Education for the Doctoral Program of Higher Education (20114306110006), and the Cooperative Innovation Center of Engineering and New Products for Developmental Biology of Hunan Province for financial support.
Author Disclosure Statement
No competing financial interests exist.