
Abstract
The Eph receptors are transmembrane proteins that belong to the receptor tyrosine kinases superfamily. Elevated Eph/ephrin expression levels have been associated with angiogenesis and tumor vasculature in many types of human cancers, including breast, lung, and prostate cancers, melanoma, and leukemia. In glioblastoma (GBM), the dysregulated expression of Eph receptors and of corresponding ephrin ligands has been associated with higher tumor grade and poor prognosis making them effective targets for therapeutic drugs. In this study, we describe the GL43.T, an anti-Eph aptamer, able to bind at high-affinity EphB3 and EphB2. Moreover, the GL43.T aptamer inhibits the glioma cell vitality and interferes with ephrine-B1 inhibition of chemotactic serum-stimulated cell migration. GL43.T aptamer represents a promising therapeutic molecule for EphB3-dependent cancers.
Introduction
T
Because of the peculiar mode of signalling of Eph/ephrin complexes and the resulting bidirectional way of action, it is important to develop molecules able to interfere with the functional interaction of ephrins with their proper receptors. To date, several targeting molecules have been developed, including antibodies, small kinase inhibitors, interfering RNAs, peptides, and recombinant Eph/ephrin extracellular domain proteins [7].
Recently, nucleic acid aptamer ligands are emerging as highly promising neutralizing molecules for targeted RTKs. They are short-structured single-stranded RNA or DNA that, due to their specific three-dimensional conformation, bind at high specificity and affinity to their target molecules. Aptamers possess many advantages as therapeutic reagents over proteins and peptides, including low costs of synthesis with high batch-to-batch fidelity, no immunogenicity and improved rapid tissue penetration, and long-term stability by easy chemical modifications [8].
In this study, we characterized a truncated 2′-fluoro pyrimidine (2′-F-Py)-containing aptamer, named GL43.T, as a high-affinity ligand of EphB3 and EphB2 receptors. The aptamer colocalizes with the EphB3 receptor on target cells and it is rapidly internalized in the cell after 30 min of incubation. Furthermore, we show that GL43.T inhibits cell vitality and functionally interferes with ephrinB1-induced cell adhesion. The ability of aptamers to act as cell selective targeting carrier for interfering RNAs and chemotherapeutic drugs [9,10] makes GL43.T a promising new kind of therapeutic molecule for EphB-dependent cancers.
Materials and Methods
Aptamers
GL43.T, 5′ FAM-labeled GL43.T (GL43.T-FAM), control aptamers and 5′ FAM-labeled control aptamer, and biotinylated GL43.T and biotinylated control aptamer were synthesized by Tebu-bio srl (Milan, Italy), and contained 2′-F-Py. Before each treatment, aptamers were subjected to a short denaturation–renaturation step (5 min 85°C, 3 min on ice, 10 min at 37°C).
Cell lines and transfection
All cell lines were purchased from the ATCC (LG Standards, Milan, Italy). Human Glioblastoma U87MG and U251MG were maintained in Dulbecco's Modified Eagle's Medium (DMEM). NSCLC A549 cells were grown in the Roswell Park Memorial Institute medium (RPMI; Sigma, St. Louis, MO) and human glioma LN18 in Advanced DMEM (Gibco, Life technologies, Milan, Italy). Media were supplemented with 10% heat-inactivated fetal bovine serum (FBS) for U87MG, U251MG, and A549 or with 5% FBS for LN18.
For EphB3 gene silencing, U87MG (1.4 × 105 cells/2 mL of standard medium per 3.5-cm plate) were transfected with siRNA EphB3 or siRNActrl (100 nM; TriFECTa® RNAi Kit; IDT Integrated DNA Technologies, Coralville, IA) and Lipofectamine 2000 (Invitrogen, Carlsbad, CA) in Opti-MEM I reduced serum medium. After a 5-h incubation at 37°C, a complete culture medium was added to the cells and incubation was prolonged up to 24 h. For EphB3 gene expression, LN18 cells (2 × 105 cells/2 mL of standard medium per 3.5-cm plate) were transfected with pCMV6-EphB3 plasmid (1 μg/100 μL; OriGene Technologies, Rockville, MD) with Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA), according to the manufacturer's instruction. After a 5-h incubation at 37°C, the complete culture medium was added to the cells and incubation was prolonged up to 48 h.
RTK antibody arrays
U87MG cells were serum starved overnight, pretreated with 200 nM GL43.T aptamer or G0 pool as control for 3 h, and then stimulated with 20% FBS either alone or in presence of the aptamer or the pool. RTK antibody arrays (R&D Systems, Minneapolis, MN) were performed as recommended. Briefly, cells were lysed and the lysates were incubated overnight with the phospho-RTK array. Following three washes, arrays were determined by incubation with anti-phosphotyrosine horseradish peroxidase.
Pull-down with biotinylated aptamer
Biotinylated GL43.T or unrelated aptamer used as control were incubated in a serum-free culture medium at 200 nmol/concentration on U87MG cells for 30 min at room temperature. Cells were washed with phosphate-buffered saline (PBS) and lysed with 10 mM Tris–HCl pH 7.5 containing 200 mM NaCl, 5 mM EDTA, 0.1% Triton X-100, and protease inhibitors. 400 μg-cell extract was incubated with 200 μL streptavidin beads (Thermo Fisher Scientific, Waltham, MA) in 0.4 mL lysis buffer at RT for 2 h with rotation. Following three washes with PBS, bound proteins were recovered with the Laemmli buffer and analyzed by immunoblotting with anti-EphB3 antibody.
Immunoblot analysis
Cell extracts were prepared as described [11]. For immunoblot analysis, the following primary antibodies were used: anti-EphB3 receptor antibody (ab133742), purchased from Abcam (Cambridge, United Kingdom), and anti-α-tubulin (DM 1A; Sigma). Immunoblot was performed as recommended.
Binding assay by reverse transcription quantitative polymerase chain reaction
Cells were seeded in 3.5-cm plates, transfected with siRNA EphB3, siRNActrl, or with pCMV6-EphB3 plasmid (as reported in cell line and transfection section), and left growing for 24 or 48 h, respectively. Cells were then treated with 100 or 200 nM of GL43.T for 30 min at 37°C. Unbound RNA was removed by washing thrice with ice-cold PBS. Bound RNA was recovered by TRIzol containing 0.5 pmol/mL of CL4 aptamer (CL4: 5′GCCUUAGUAACGUGCUUUGAUGUCGAUUCGACAGGAGGC3′) used as a reference control. The amount of recovered RNAs was determined by performing a two-step reverse transcription-quantitative polymerase chain reaction protocol as reported [12].
Primers used were:
GL43.T (Forward) 5′ GGGAGACAAGAATAAAC 3′;
GL43.T (Reverse) 5′ GTTGCAAGAGTAACGTT 3′;
CL4 (Forward) 5′ GCCTTAGTAACGTGCTTT 3′;
CL4 (Reverse) 5′ GCCTCCTGTCGAATCG 3′.
Data were normalized to the CL4 reference control and to cell number, as determined by counting cells cultured in conjunction with each experiment.
Immunofluorescence analyses
Cells, grown on glass coverslips overnight, were treated at different incubation times with 2.5 μM of GL43.T-FAM, dissolved in media containing 10% FBS at 37°C, washed thrice with PBS, and fixed with paraformaldehyde 4% in PBS for 10 min. Then, coverslips were mounted on microscope slides with prolong gold antifade reagent with DAPI (Invitrogen, Waltham, MA) and the cells were visualized by confocal microscopy.
To assess aptamer-EphB3 colocalization, cells were incubated in media in the presence of GL43.T-FAM for 60 min at 37°C. Then, cells were washed and incubated with anti-EphB3 antibody (Abcam) for 30 min at 37°C before fixation. Cells were then fixed, permeabilized, and incubated at 37°C for 30 min with Alexa568 secondary antibody (Invitrogen, Waltham, MA). Images were obtained using a Zeiss 510 LSM confocal microscope with a 63× oil objective. Laser intensity has been set at 30.7% for Ar 488 and 68.3% for HeNe 543.
Microscale thermophoresis
GL43.T-FAM, at constant concentration of 100 nM, was incubated with various concentrations (from 0.24 to 8.000 nM) of EphB3 purified recombinant protein or an unrelated protein used as negative control (R&D System). A serial dilution of EphB3 was prepared in the buffer consisting of 1× PBS, pH 7.4, 1 mM MgCl2, and 0.15% Tween-20. Microscale thermophoresis (MST) measurement was carried out on a Monolith NT.115 (NanoTemper Technologies GmbH, Munchen, Germany) using standard capillaries. KD was determined as described [13].
Fluorescent filter-binding assay
Fluorescent filter-binding assay was performed by incubating 5 nM purified EphB2 or EphB3 recombinant proteins (R&D System) with increasing concentrations of GL43.T-FAM (10, 25, 50, 100, 200, 500, 800 nM) in PBS supplemented with bovine serum albumin (BSA) 0.01%. After 30 min of incubation at 37°C, samples were filtered through HAWP 0.45-μm nitrocellulose filters (Millipore, Bedford, MA) pretreated with PBS. The retained RNAs were eluted by cutting the filters and incubating for 2 h at 42°C in an elution buffer (20 mM Tris–HCl pH 7.8, 0.2% sodium dodecyl sulfate (SDS) and 0.3 M sodium acetate). Then, RNAs were precipitated and loaded on acrylamide-urea gel. Increasing amounts of each purified aptamer (at the concentrations of 5, 10, 20, 40, 80, 160, 320, 640 nM in a final volume of 20 μL) were loaded as standard to quantify samples (standard curves).
The fraction of aptamers bound to the protein was visualized by Gel Doc™ EZ Imaging System with Image Lab™ (Bio-Rad Laboratories, Hercules, CA) and quantified by comparison to the correspondent standard using Image Lab™ Software Version 5.1 (Bio-Rad Laboratories).
Cell viability
Cell viability was assessed with CellTiter 96H AQueous One Solution cell Proliferation Assay (Promega, Madison, WI) as recommended. Cells (1.6 × 103 cells/well) were plated in 96-well plates and allowed to grow overnight before treatment with GL43.T or unrelated aptamer (200 nM final concentration) at different time points (1, 2, 3, 5, 7 days), renewing the treatment each 24 h. Absorbance at 492 nm was measured with MULTISKAN FC (Thermoscientific, Waltham, MA). The results were expressed as percent of viable treated cells with respect to control untreated cells.
Transwell migration assay
Cell migration was assessed as reported [14]. Briefly, A549 cells were treated for 3 h with 200 nM GL43.T or with unrelated aptamer and then trypsinized, resuspended in RPMI serum free, and counted. Cells (7.5 × 104 in 100 μL serum-free medium per well) were then plated into the upper chamber of a 24-well transwell (Corning Incorporate, Corning, NY) in the presence of GL43.T or unrelated aptamer and exposed to 10% FBS as inducer of migration in the absence or presence of the extracellular domain of ephrinB1 ligand (2 μg/mL; R&D Systems). After a 24-h incubation at 37°C in humidified 5% CO2, migrated cells were visualized by staining with 0.1% crystal violet in 25% methanol. Percentage of migrated cells was evaluated by eluting crystal violet with 1% SDS and reading the absorbance at 594 nm wavelength.
Annexin V-FITC apoptosis detection assay
U87MG cells were left untreated or treated with GL43.T (200 nM final concentration) at different time points (3 and 7 days), renewing the treatment each 24 h. Cells were harvested and centrifuged for 5 min. Supernatant was decanted, cells were washed twice with 1 mL of cold 1× PBS, and then suspended in 1× binding buffer at a concentration of 1 × 106 cells/mL. Annexin V-FITC and PI (Novus Biologicals, Littleton, CO) were added at 100 μL of each sample according to the manufacturer's recommendations (5 μL of Annexin V-FITC+PI). Samples were then incubated for 20 min at RT in the dark. Four hundred microliters of 1× binding buffer was added to each sample before flow cytometry analysis with BD Accuri C6 (BD Biosciences, Franklin Lakes, NJ).
Cell cycle analysis
U87MG cells were left untreated or treated with GL43.T (200 nM final concentration) at different time points (3 and 7 days), renewing the treatment each 24 h. Cells were harvested and centrifuged for 5 min. Supernatant was decanted, cells were washed with 1 mL of cold 1× PBS, and then fixed with ethanol 70% on ice for 10 min. Samples were then washed twice with 1 mL of cold PBS to completely remove ethanol. Cell pellet was suspended in 400 μL of PBS with 50 μg/mL RNase A and 50 μg/mL PI (Sigma). Tubes were incubated for 20 min at 37°C. Cell cycle was analyzed with BD Accuri C6 (BD Biosciences).
Results and Discussion
GL43.T aptamer interacts with EphB3
By differential cell SELEX on human glioma cell lines, we have generated the 2′-F Py-containing RNA aptamer, named GL43, and identified the functional truncated sequence consisting of residues from bases 1 to 39 (herein named GL43.T, Fig. 1A). The aptamer was identified following 14 rounds of positive selection on high malignant human glioma U87MG cells, each preceded by counter-selection on poorly tumorigenic T98G glioma cells [15].
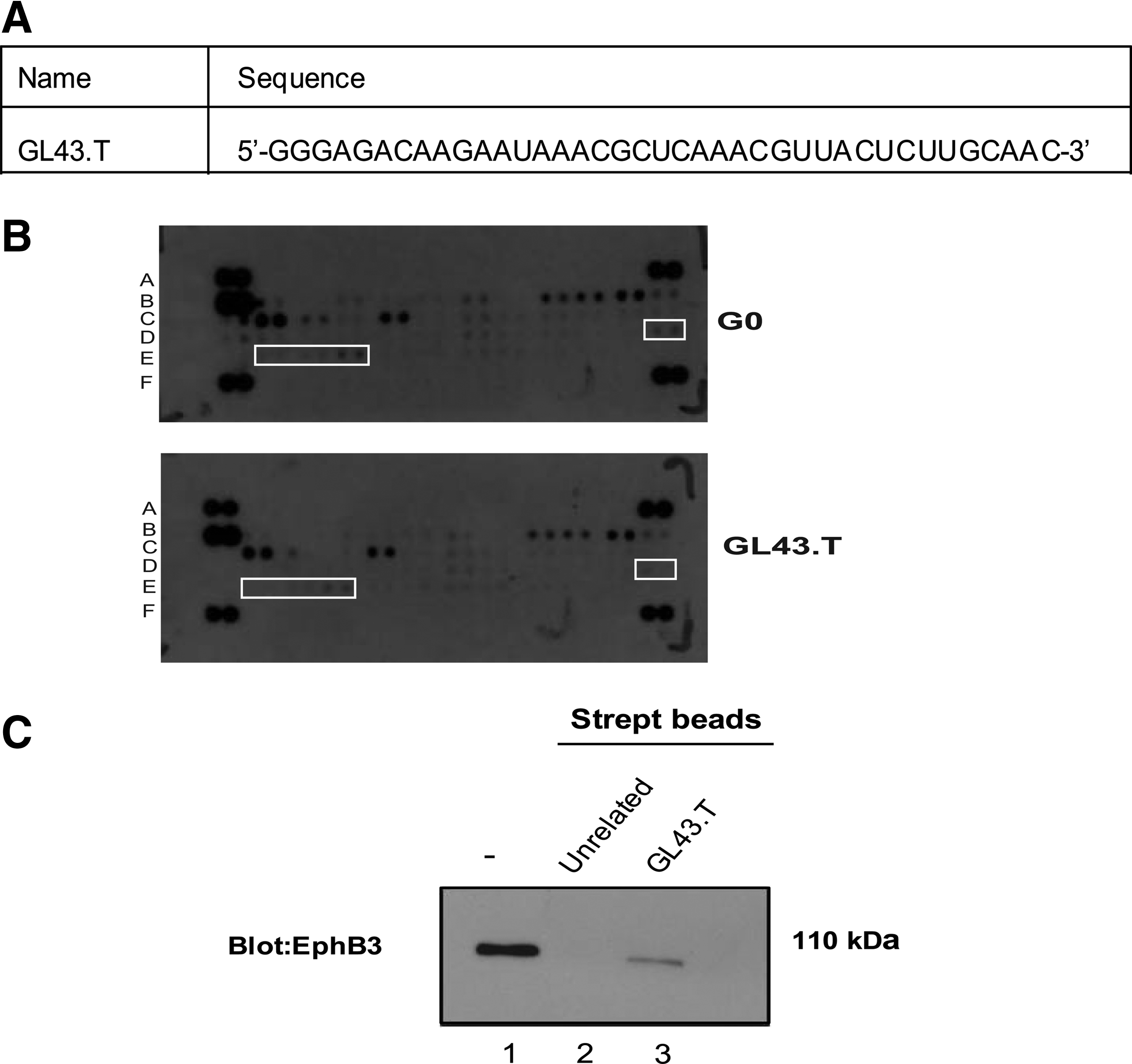
The anti-Eph GL43.T aptamer.
To identify the functional targets of GL43.T aptamer sequence, we performed a phospho-RTK antibody array analysis. As shown in Fig. 1B, the treatment of U87MG target cells with GL43.T decreased the phosphorylation of detectable Eph receptors, suggesting a functional interaction of the aptamer with members of the Eph receptor family.
Next, we used GL43.T to pull down proteins expressed on the U87MG cell surface and determined whether the GL43.T aptamer could bind EphB3, a member of the less-characterized EphB receptors subfamily that was not present in the array. To this end, U87MG cells were treated for 30 min with biotinylated GL43.T aptamer or with an unrelated sequence, used as a negative control. Cell extracts were purified on streptavidin-coated beads and analyzed by SDS-polyacrylamide gel electrophoresis (PAGE), followed by immunoblotting with anti-EphB3 antibodies. As shown in Fig. 1C, GL43.T aptamer, but not the unrelated sequence, was able to pull down the EphB3 receptor indicating the direct binding with this receptor protein.
In addition, we analyzed the relative expression levels of the EphB3 receptor in a panel of stable cell lines in which the binding efficiency of the GL43 aptamer has been previously determined [15]. As shown, EphB3 was strongly expressed in three human glioma-derived cell lines (U87MG, T98G, U251MG) in the mouse fibroblast NIH3T3 cells and in the human NSCLC A549 cell line, but poorly in the LN18 human glioma cells (Fig. 2A). The expression levels of EphB3 well correlate with the previous reported aptamer cell-binding efficacy, further supporting GL43.T ability to recognize EphB3.
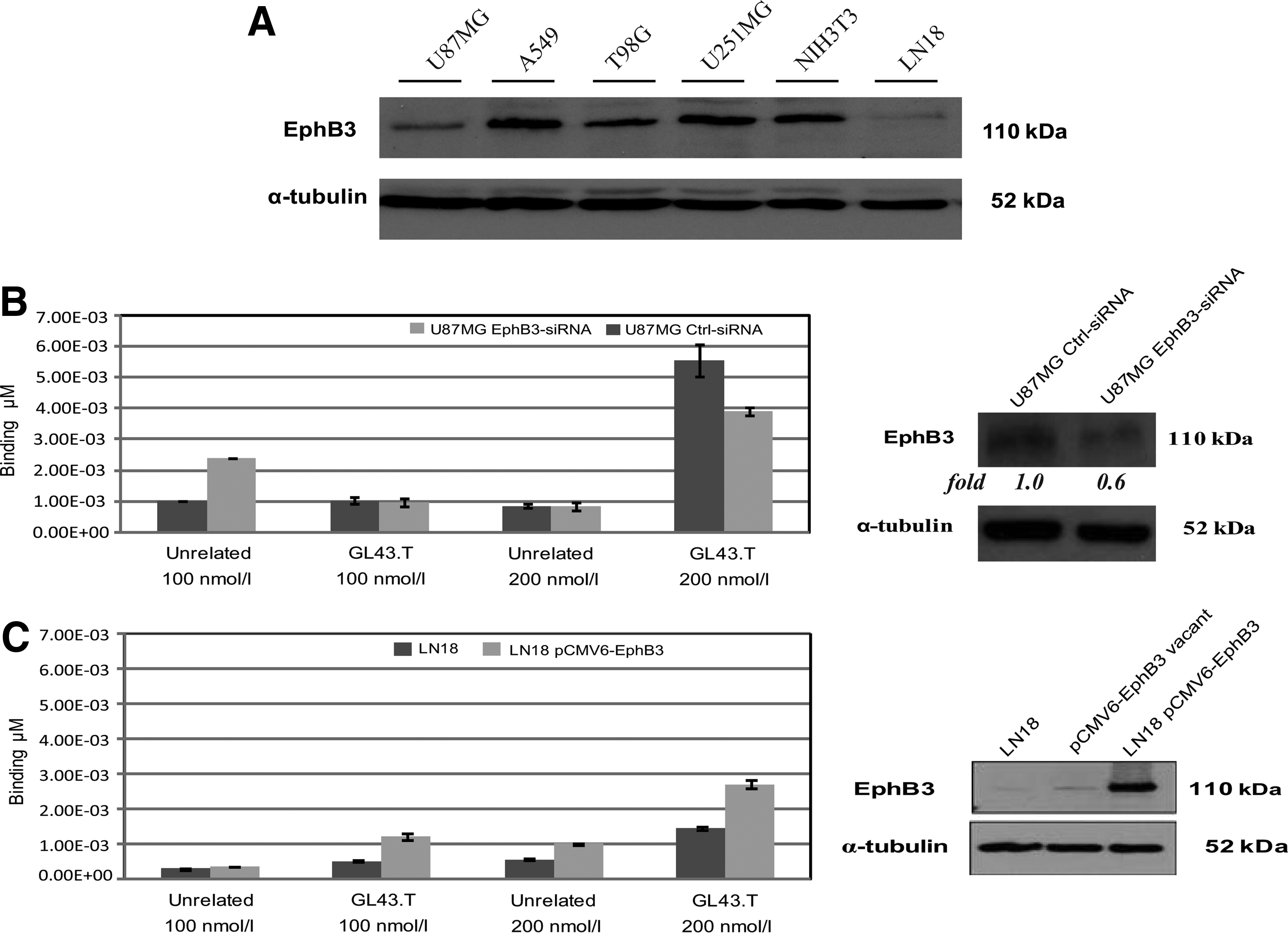
GL43.T-EphB3 interaction characterization.
Accordingly, the transfection of U87MG cells with EphB3-specific siRNA decreased the corresponding cellular protein levels of about 40% (Fig. 2B, right), consequently reducing GL43.T binding of ∼30% (Fig. 2B, left). Conversely, the forced expression of the EphB3 receptor in low-expressing LN18 cells (Fig. 2C, right) elicited the GL43.T binding to the LN18 cells, both at 100 and 200 nM (Fig. 2C, left). Taken together, these data indicate the EphB3 as a target of GL43.T.
Binding affinity measure by MST analysis
To measure the binding affinity of GL43.T aptamer and EphB3 receptor, we performed MST analysis. GL43.T-FAM aptamer was incubated with increasing concentrations of purified recombinant EphB3 protein or unrelated recombinant protein used as negative control. Our results showed that GL43.T aptamer bound to the EphB3 receptor exhibiting a KD of 433.5 nM (±136.6), whereas did not show a measurable affinity to the unrelated protein (Fig. 3). This result confirms the aptamer ability to recognize EphB3 on tumor cell surface as well as the soluble recombinant protein.
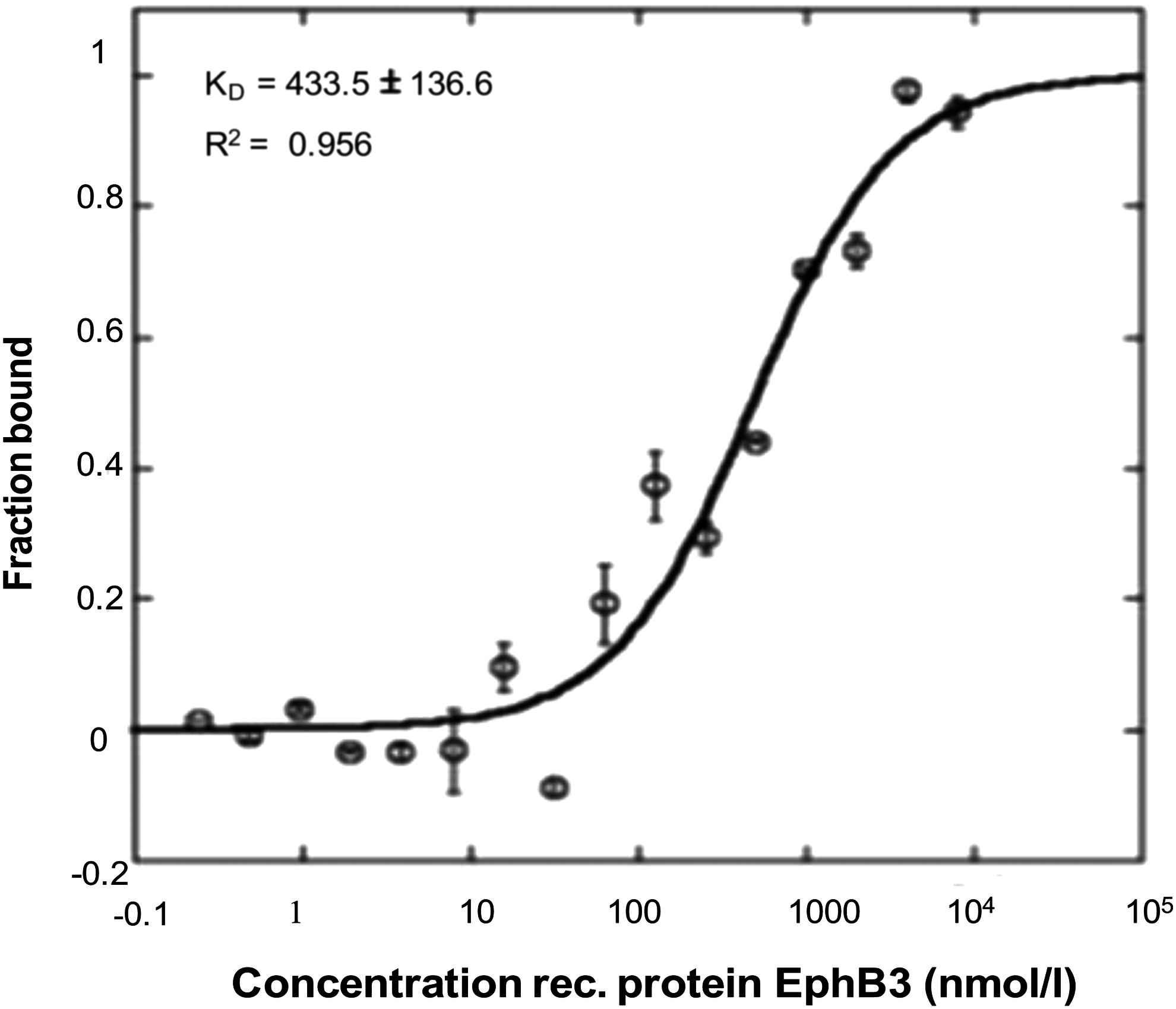
Binding curve of GL43.T-EphB3 protein. Interaction of GL43.T and EphB3 protein was analyzed by MST by incubating 100 nM of GL43.T with increasing concentration (from 0.24 to 8.000 nM) of EphB3 purified recombinant protein. The binding data were fitted, and the dissociation constant (KD) was calculated.
GL43.T targeting by confocal immunofluorescence microscopy assay
To further investigate the ability of GL43.T aptamer to recognize EphB3 receptor, we then performed an immunofluorescence assay.
As shown in Fig. 4A, GL43.T-FAM preferentially binds U87MG EphB3-positive cells compared to LN18 cells that show low EphB3 expression level (Fig. 2A). Indeed, U87MG cells, treated with GL43.T-FAM, showed clearly visible fluorescent spots that increased with increasing incubation times (Fig. 4A). Conversely, LN18 cells showed a low fluorescent signal as consequence of a poor aptamer–receptor interaction.

Immunofluorescence analyses.
The binding identity of GL43.T aptamer to EphB3-expressing cells was further characterized by colocalization experiment between GL43.T-FAM and EphB3 stained with its specific antibody. As shown in Fig. 4B, in U87MG target cells, signal overlap was observed both on the cell surface and intracellularly. Intracellular aptamer fluorescent spots (in green) that do not colocalize with the antibody red fluorescence were as well visible, likely because of partial dissociation of aptamers from receptors upon internalization. As expected, very low signal was instead observed in LN18 cells. Furthermore, to assess the aptamer ability to internalize into target cells, we carried out a Z-stack acquisition. Following 30 min of treatment, the aptamer appeared to be internalized (Fig. 4C) in agreement with the rapid internalization rate of EphB3 [4].
Target specificity of GL43.T
Since EphB3 shows high homology to EphB2 [16], we compared the binding of GL43.T aptamer to these receptors by performing a fluorescent filter-binding assay. Increasing amounts of GL43.T-FAM were incubated with EphB3 or EphB2 purified protein. An unrelated sequence of GL43.T was used to estimate nonspecific binding. As shown, GL43.T was able to bind both EphB2 (Fig. 5A) and EphB3 (Fig. 5B) more effectively compared to the control aptamer.
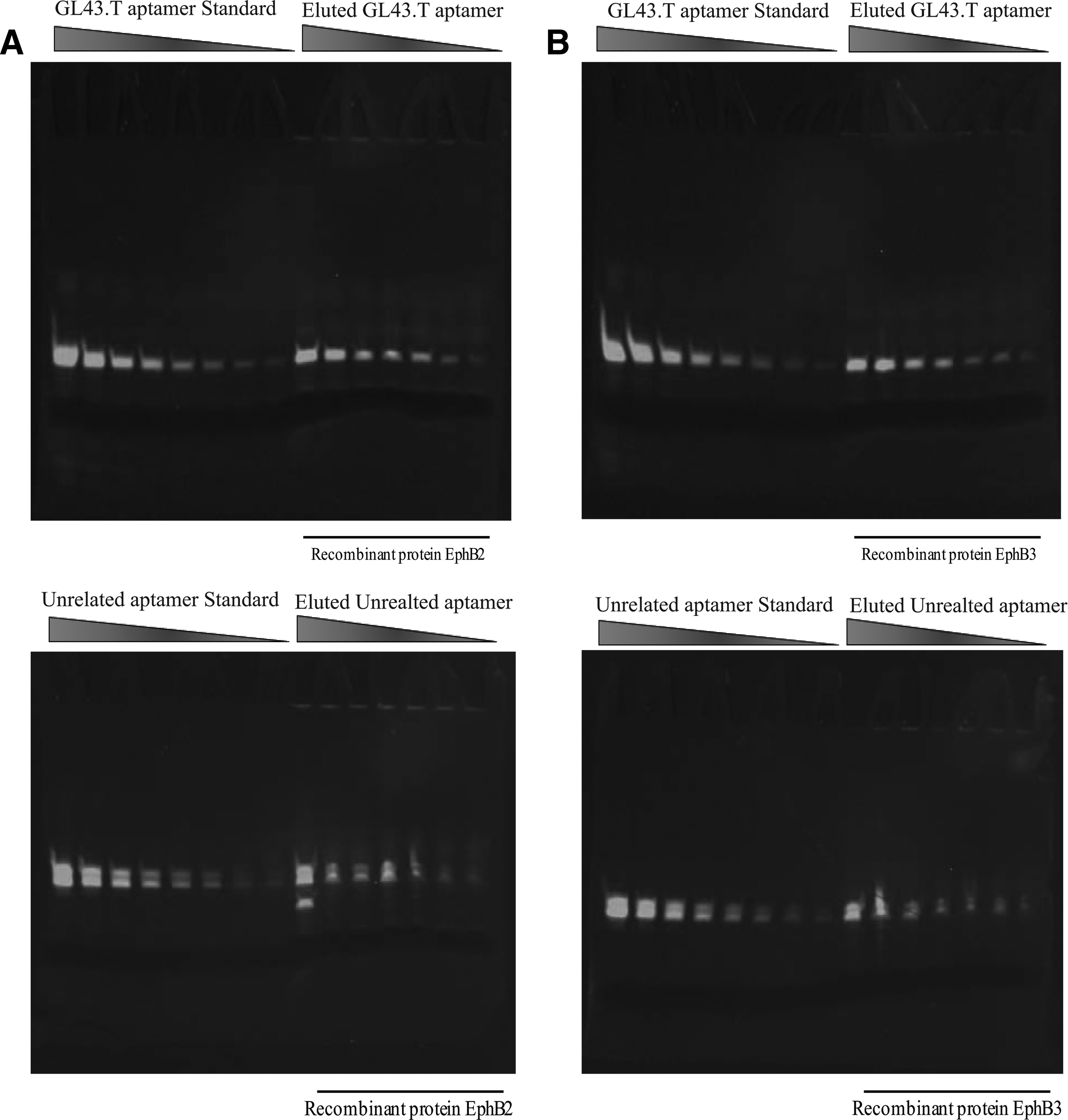
Fluorescent filter-binding assay. Samples of GL43.T-FAM (upper panels) or unrelated-FAM aptamer (lower panels) incubated with EphB2
GL43.T blocks cell proliferation and antagonizes migration
Intracellular signaling initiated by Eph receptors in glioma has been reported to be involved in cell proliferation and migration [4]. Thus, we asked whether, GL43.T may suppress cell proliferation through inhibiting EphB3/EphB2 receptor. To this end, we determined cell viability by MTT assay in U87MG. Cells were plated in 96-well plates and allowed to grow overnight before exposure to GL43.T for different times of treatment. Our data showed that GL43.T strongly inhibited cell viability in a time-dependent manner compared to a control unrelated aptamer (Fig. 6A, left). The same evidence was also found in U251MG glioma cells (Fig. 6A, middle). In accordance with low binding ability, no pronounced effect on cell viability was determined on EphB low-expressing LN18 cells (Fig. 6A, right), thus confirming the dependence of GL43.T action on the expression levels of the receptor. In addition, we found that GL43.T treatment resulted in the reduction of U87MG cell number at 7 days (almost 50%) without affecting the distribution of cell cycle phases following PI incorporation (Fig. 6B, C). Furthermore, as assessed by flow cytometry, the aptamer determined an increase in Annexin V/PI-positive cells at 7 days of treatment (Fig. 6D). These results suggest that GL43.T treatment decreases cell survival by triggering cell death. As reported by Wang et al. [17], overexpression of the EphB2 receptor may uncouple the effects on glioblastoma (GBM) tumorspheres by promoting cell migration, while inhibiting cell proliferation.
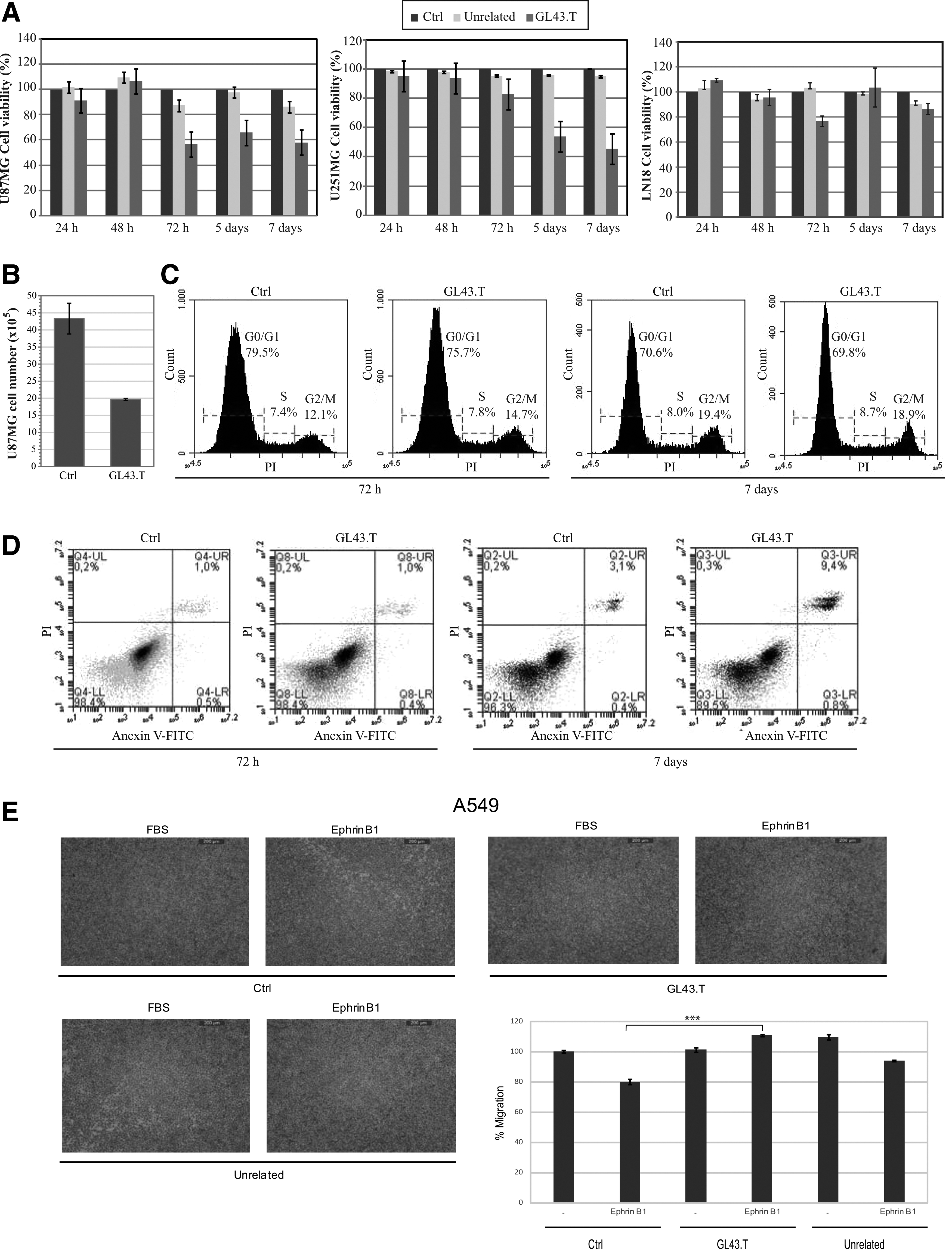
GL43.T functional analyses.
Furthermore, it has been reported in A549 NSCLC cells that upon treatment with the ephrin-B1 soluble ligand, the activation of the EphB3 receptor resulted in the inhibition of cell migration without affecting cell proliferation [6]. Therefore, to investigate the ability of GL43.T to influence cell migration, we decided to use the A549 cells. According to Li et al. [6], by binding to EphB3, ephrin-B1 negatively regulates serum-stimulated A549 cell migration with a reduction of ∼20% (Fig. 6E). Consistently, treating cells with the GL43.T aptamer antagonizes ephrin-B1, restoring serum-dependent stimulation of cell migration (Fig. 6E). No interference with the inhibitory effect of ephrin-B1 was instead observed when A549 cells were treated with an unrelated aptamer.
Because of the promiscuity of target recognition, the GL43.T aptamer plausibly does not discriminate between EphB receptors likely binding EphB2/EphB3 and antagonizing the ephrin/EphB interaction. Taken together, our results indicate that by binding to EphB2/EphB3 receptors, GL43.T interferes with regulation of cell proliferation and migration, thus acting as a functional inhibitor of receptors.
GL43.T aptamer would be suitable as a therapeutic molecule for recognition and drug targeting in EphB-dependent cancers.
Footnotes
Acknowledgments
This work was supported by funds from the following: MIUR grant, MERIT RBNE08YFN3_001 (VdF), and AIRC No. 13345 (VdF); from the Italian Ministry of Economy and Finance to the CNR for the Project FaReBio di Qualità (VdF) and Compagnia San Paolo No. 2011.1172 (VdF); Italian Ministry of Health, GR-2011-02352546 (CLE). We wish to thank L. Baraldi for technical assistance, and acknowledge L. Cerchia and T. Schubert for advice.
Author Disclosure Statement
No competing financial interests exist.