
Abstract
Aptamers are proving their utility in a number of applications. However, to be easily functionalized, their structure needs to be simplified. Therefore, we sought to truncate a 50-nucleotide aptamer specific to the transferrin receptor to its smallest functional unit using rational engineering of the predicted two-dimensional structure of the longer parent sequence. In addition, mutations were introduced into the binding loop to determine their effect on the selectivity of the aptamers. These base mutations enhanced the binding affinity of the aptamer, while retaining its specificity. The equilibrium dissociation constant (Kd) was reduced sixfold following the substitution of all four bases in the binding region. In addition, these aptamers were efficiently internalized into transferrin receptor-positive cells in a similar manner to the transferrin receptor antibody and demonstrated colocalization with this antibody. This study has shown that the smallest functional unit of this aptamer was 14 nucleotides. This small size will be advantageous for future applications, such as drug delivery or functionalization of other therapeutic modalities.
Introduction
T
A novel strategy has been to utilize receptors on the BBB to actively transport molecules into the brain by means of receptor-mediated transcytosis, with the transferrin receptor (TfR) becoming one of the most widely targeted receptors [6]. The TfR is a membrane glycoprotein involved in iron homeostasis that is highly expressed on the surface of the BBB and has been identified as an effective target for receptor-mediated transcytosis [7]. Although the TfR is saturated in most cell types it is expressed on, such as red blood cells, hepatocytes, intestinal cells, macrophages, and the brain itself, it is not saturated on the BBB, where the expression is considerably higher [7,8]. Yu et al. exploited the TfR for the purpose of active transport across the BBB using an antibody, where they demonstrated that a lower binding affinity of the antibody allowed binding, internalization, and subsequent exit from the cells of the BBB into the brain [9]. However, given that antibodies carry some immunological risk and severe side effects, a novel therapeutic strategy with fewer side effects is required.
Aptamers are small single-stranded DNA or RNA molecules that are also known as chemical antibodies due to their ability to fold into complex three-dimensional shapes and bind to their target in a similar manner to conventional antibodies. However, one key benefit of these ligands is that they are chemically synthesized, thus reducing many of the disadvantages of antibodies [10–14]. Aptamers are more stable, nonimmunogenic, and 10–20 times smaller than an antibody, thus allowing for superior tissue penetration [15]. Recently, Chen et al. developed DNA and RNA aptamers that selectively recognize the extracellular domain of the mouse TfR [16]. The originally selected DNA aptamer, GS24, was 64 nucleotides long and Chen et al. truncated this aptamer to 50 nucleotides, while retaining its selectivity [16]. This aptamer was further truncated to 35 nucleotides by Porciani et al., although with a significant loss in binding affinity, as demonstrated by the five-fold higher Kd (dissociation constant) [17]. However, through mutations in the stem region of the aptamer, Porciani et al. were able to improve the Kd and demonstrate superior properties compared to the original GS24 aptamer [17].
In this study, we truncate the DNA aptamer GS24 to a 14-nucleotide sequence using rational engineering of the predicted two-dimensional structure to determine the minimum sequence required to maintain specificity and sensitivity. Through mutations in the binding region of the aptamer, we were able to generate an aptamer with high specificity and sensitivity for further functionalization.
Materials and Methods
Cell lines and cell culture
The cell lines of human and mouse origin used in this study were purchased from the American Type Culture Collection. They are mouse brain endothelial cells, bEnd.3, and human acute lymphoblastic leukemia cells, MOLT4. Cells were grown and maintained in culture with Dulbecco's modified Eagle's medium (Life Technologies) supplemented with 10% fetal calf serum (FCS). Cells were maintained at 37°C in a 5% CO2 atmosphere.
Aptamers
The truncated GS24 aptamer, TfRA1, the three mutated versions of the TfR aptamer, TfRA2, TfRA3, and TfRA4, and the scrambled sequence aptamer were purchased from Integrated DNA Technologies (IDT) and HPLC purified. All oligonucleotide sequences were labeled with a TYE665 fluorophore on the 3′ end. The sequences were as follows:
TfRA1: 5′-GC GTG TGCA CAC GC-3′ TfRA2: 5′-GC GTG GGCC CAC GC-3′ TfRA3: 5′-GC GTG GTCA CAC GC-3′ TfRA4: 5′-GC GTG GTAC CAC GC-3′ Scrambled: 5′-GC GCG TGCT CGC GC-3′
Determination of aptamer affinity
The Kd of the DNA aptamers to TfR-positive cells (bEnd.3 cells) and TfR-negative cells (MOLT4 cells) was determined using flow cytometry. The aptamers were prepared by folding into their three-dimensional structure using a thermocycler (PerkinElmer) (85°C for 5 min, slow cooling to 22°C over 10 min, and 37°C for 15 min). The cells (5 × 105) were first incubated with a binding buffer for 1 h on ice (Dulbecco's phosphate-buffered saline with 1 mM MgCl2, 0.1 mg/mL tRNA, 1 mg/mL bovine serum albumin, and 10% FCS) followed by two washes with the binding buffer before incubation with serial concentrations of TYE665-labelled aptamer in a 100 μL volume of binding buffer for 30 min at 37°C. The cells were washed thrice with the binding buffer, resuspended in 150 μL binding buffer, and subjected to flow cytometric analyses (FACS Canto II flow cytometer; Becton Dickinson).
Confocal microscopy
Twenty-four hours before labeling, cells were seeded at a density of 75,000 cells/cm2 in an eight-chamber slide (Lab-Tek II; Nunc). Each aptamer was prepared in the same manner as for flow cytometry. Following removal of media, cells were incubated in the binding buffer at 37°C for 15 min, and washed twice in binding buffer before incubation with 200 nM aptamer for 60 min at 37°C. Bisbenzimide Hoechst 33342 (3 μg/mL; Sigma) was added to the cells during the final 10 min of incubation. The aptamer solution was removed and the cells washed thrice for 5 min each in the binding buffer before visualization using a FluoView FV10i laser scanning confocal microscope (Olympus). To assess the mechanism of cellular uptake, the internalization of a phycoethyrin-labeled rat anti-mouse TfR monoclonal antibody (R17217; Abcam) was also visualized by confocal microscopy, by coincubating the antibody with the aptamer at 1 μg/mL for 30 min. The captured images were then analyzed using an ImageJ correlation analysis to calculate the Pearson's correlation coefficient, with 50 cells analyzed in total [18].
Results and Discussion
A recent promising technique for the treatment of neurodegenerative disorders has been the development of therapeutic modalities capable of transcytosing into the brain by receptor-mediated transcytosis [6]. The potential for the TfR to be utilized in this manner has previously been explored through the application of therapeutic antibodies [7,19]. Through this preliminary work, it was shown that by altering the binding affinity of the therapeutic antibody, to a moderate to low affinity for the TfR, the antibody was capable of entering the brain, rather than remaining attached to the transferrin receptor [9]. However, given the limitations associated with therapeutic antibodies, especially when contemplating modalities capable of entering the brain, a less toxic compound, such as a chemical antibody, is needed [20–22]. Therefore, we sought to engineer an aptamer with similar characteristics to these therapeutic antibodies, but which have fewer side effects.
Truncation of the GS24 aptamer
Recently, a DNA aptamer to the extracellular domain of the mouse transferrin receptor (mTfR) was generated [16]. This GS24 aptamer was originally 64 nucleotides in length. Through truncation, it was reduced to 50 nucleotides, with its specificity confirmed using mTfR-transfected HEK293T cells [16]. As both the full length and truncated version of the GS24 aptamer possessed a loop region of 12 nucleotides (Fig. 1A, B), we postulated this to be the primary binding region of the aptamer to the mTfR. Accordingly, we performed a truncation to determine if this would lead to a suitable minimized sequence for further functionalization. Truncation of aptamer sequences has several advantages, notably a reduction in production costs and a potential increase in aptamer affinities [23]. Therefore, this 12-nucleotide sequence was interrogated for its secondary structure using the structure prediction software, MFold. A 14-nucleotide sequence, TfRA1, was synthesized based on the initial full-length GS24 structure and the truncated version generated by Chen et al. [16] (Fig. 1C). In addition, mutations were introduced into the binding region to modulate the binding affinity of the truncated aptamer to the mTfR (TfRA2, TfRA3, and TfRA4). These mutations were introduced into the binding region for similar reasons to those observed in the study by Porciani et al., whereby the original GS24 aptamer possessed a binding affinity of 250 nM, but the truncated 35-nucleotide aptamer lost some of its sensitivity (with a five-fold higher Kd) [17]. Interestingly, their mutated version of GS24, DW4, was demonstrated to have a superior binding affinity to both the full-length GS24 and the GS24min (DW4 Kd = 190 nM), suggesting that mutations may increase the binding affinity. However, mutations introduced into the stem or loop region may also lead to a loss of sensitivity and/or specificity of the aptamer [24]. Therefore, it is imperative to confirm that the mutated aptamers are still capable of recognizing their target with a high degree of sensitivity.
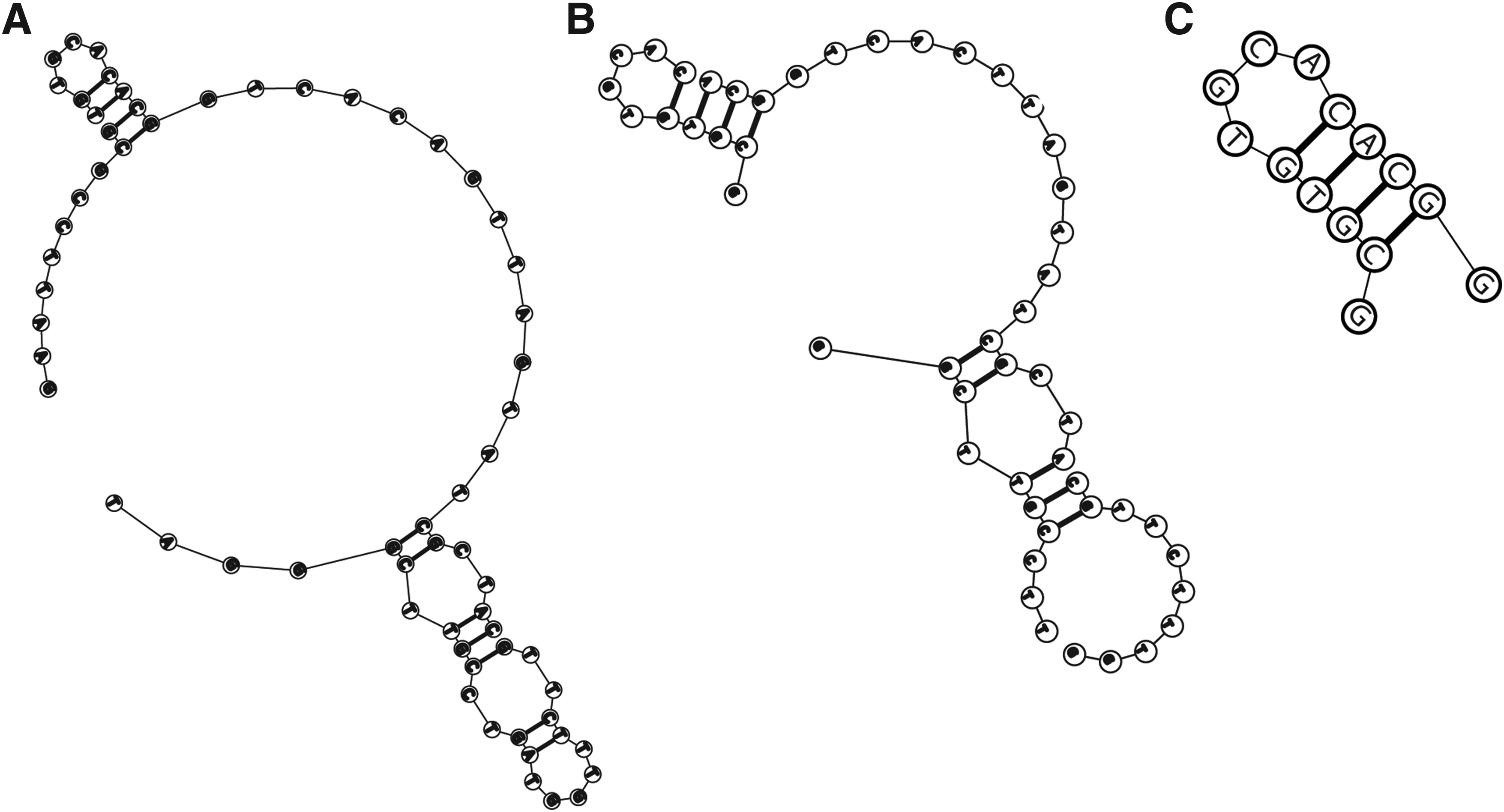
Postselection engineering of the transferrin receptor aptamer.
Enhanced sensitivity of the mutated aptamers
The binding affinities of the four newly generated aptamer derivatives, TfRA1, TfRA2, TfRA3, and TfRA4, to target cells were evaluated using flow cytometry. For future therapeutic applications, it is important to confirm that the aptamers can recognize the target in its native conformation. Therefore, a biological assay using the mouse brain endothelial cell line, bEnd.3, which expresses the mTfR, was used for this characterization. The binding affinities determined for each of the four truncated aptamers demonstrated that the mutations introduced into the binding region had a varied impact on sensitivity. Surprisingly, the initial truncation, TfRA1, which was a direct truncation of the original GS24, possessed the weakest binding affinity (Kd = 2.25 ± 0.9 μM) (Fig. 2A). As the binding region of this particular aptamer consisted of only four nucleotides, there were limited mutations that could be introduced into this four-nucleotide loop. The T6G and A9C mutation introduced into the TfRA2 aptamer resulted in an almost sixfold increase in the binding affinity (Kd = 595.7 ± 154.9 nM) (Fig. 2B), whereas the T6G and G7T mutation introduced into TfRA3 resulted in a slightly lower binding affinity compared to TfRA2 (Kd = 496.3 ± 87.2 nM) (Fig. 2C), suggesting that the change in the nucleotide at position six might have been responsible for the increased binding affinity observed for TfRA2 and TfRA3, compared to TfRA1. When all four nucleotides in the binding region were mutated (T6G, G7T, C8A, and A9C), the binding affinity of TfRA4 increased only slightly compared to TfRA2, suggesting that the nucleotides in positions six and nine are important for aptamer–target interaction, while nucleotides in positions seven and eight had a limited effect (487 ± 108.8 nM) (Fig. 2D and Table 1). None of these mutations had an effect on the specificity, as determined using the MOLT4 cell line, which is a human cell line and therefore negative for the mouse TfR (Kd > 10,000 nM). While it has been previously recognized that the mutation, or scrambling, of sequences in the binding region is one way of generating a nonbinding species, the increase in binding affinity is not unexpected, given that rational design can play a part in enhancing the properties of an aptamer [25,26]. Rational manipulation of nucleotide sequences or composition can actually improve binding properties of an aptamer as this will alter the tertiary structure of the aptamer [26]. Indeed, it is this tertiary structure that is critical for determining the interaction between the aptamer and its target [12]. Therefore, it is possible that mutations in the binding loop altered or stabilized the tertiary structure of the ligand-binding conformation of the aptamers, thus increasing the binding affinity. Finally, it is important to note that the binding assays performed by Porciani et al. were completed with purified protein, rather than in cell lines, and internalization assays were performed using a mouse fibroblast cell line, whereas this study utilized a mouse brain endothelial cell line. Cell lines possess different characteristics from that of purified proteins, with differing levels of antigen density and presentation of a particular cell surface receptor, or epitope availability, a phenomenon previously seen with antibodies and aptamers [27–29]. Therefore, a cell line, which closely resembles the future therapeutic target, such as the bEnd.3 cell line as a model of the cells of the BBB, was used in this study [30].
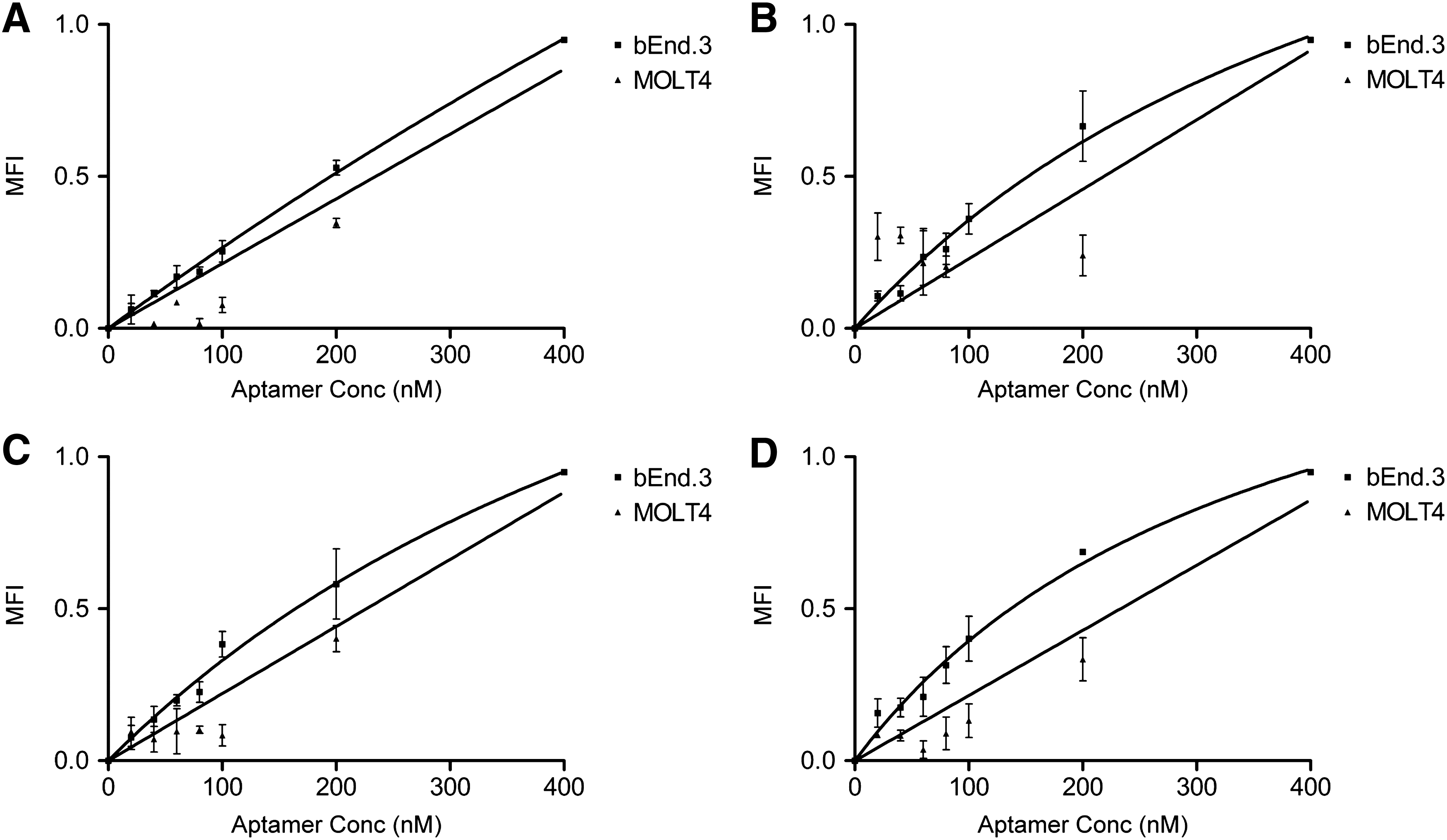
Specificity of the mTfR aptamers. TYE665-labeled aptamers were incubated with the bEnd.3 or MOLT4 cell line and analyzed by flow cytometry. The MFI was plotted against varying concentrations of mTfR aptamers (1–400 nM) at a cell density of 5 × 105 cells/mL.
Underlined region represents binding loop, and bold indicates mutated region of aptamer.
Kd, equilibrium dissociation constant.
Internalization of the engineered aptamer suggests potential therapeutic applications
The uptake of aptamers into cells is an important factor in determining the potential for their use as therapeutics [31–34]. Hence, it is critical to ensure that these aptamers are capable of being internalized into cells rather than merely attaching to the cell surface. Therefore, we sought to determine if these aptamers were internalized after binding to the transferrin receptor. To that end, 200 nM of each aptamer was incubated with bEnd.3 cells for 60 min and their ability to internalize was visualized using confocal microscopy. TfRA1, TfRA2, TfRA3, and TfRA4 were internalized to a varying degree into the bEnD.3 cells, as demonstrated by the punctate pattern of intracellular distribution, which is indicative of endocytosis (Fig. 3A), while the scrambled aptamer showed no internalization [35]. In addition, the specificity of cell entry was confirmed by the lack of internalization into MOLT4 cells. It is interesting to note that all four binding aptamers, regardless of their binding affinity, were internalized by the bEnd.3 cells. This phenomenon has been previously described in antibodies, whereby the binding affinity has influenced the rate of internalization into the cell [36]. However, other factors have also been shown to influence internalization rates, such as dissociation from the receptor and retention on the cell surface, which may be due to the part of the epitope that the binding modality binds to [36,37]. Further studies are thus required to determine the effect that these factors may have on the internalization rate of TfRA2. Internalization of TfRA1, TfRA3, and TfRA4 appeared to reach maximal internalization at 60 min, although the distribution pattern throughout the cytoplasm for TfR4A appeared to indicate a superior endocytosis profile. To confirm that the punctate pattern was due to specific internalization by receptor-mediated endocytosis as seen with transferrin, and not due to nonspecific uptake by pinocytosis or another pathway of cellular internalization [38], this aptamer, TfRA4, was incubated with the bEnd.3 cells in the presence of the anti-TfR antibody, which is known to be internalized through receptor-mediated or clathrin-mediated endocytosis [39]. While similar patterns were observed for both the aptamer and the antibody within the cell, colocalization of the aptamer and antibody was also observed (Pearson's correlation coefficient, 0.72 ± 0.05), suggesting that both modalities enter the cell in a similar manner (Fig. 3B).
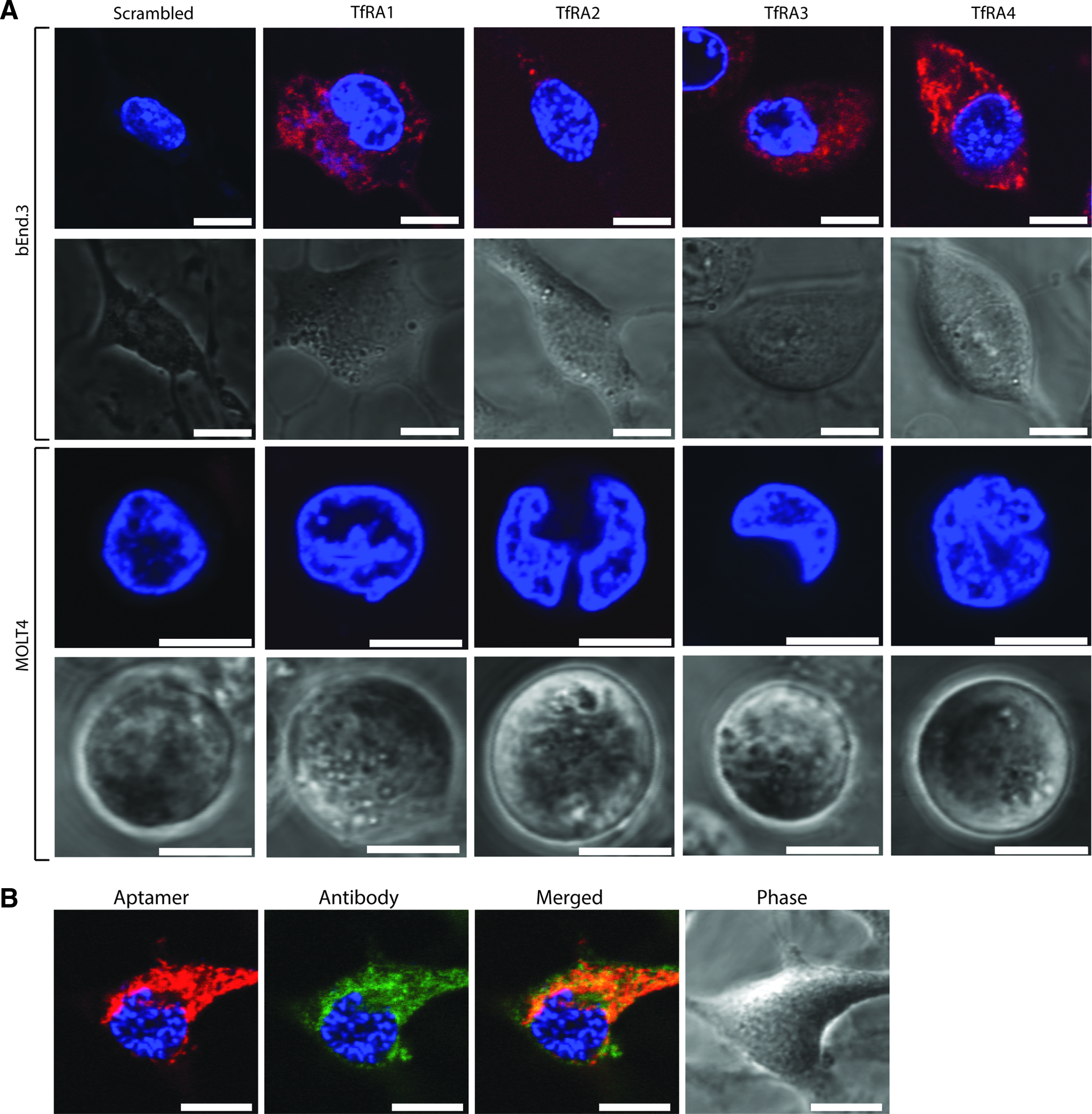
Internalization of the mTfR aptamers.
Conclusions
This study has shown that the rational truncation of an aptamer generated against the mouse transferrin receptor successfully generated an aptamer with both specificity and sensitivity. It has previously been shown that therapeutic TfR antibodies with a lower binding affinity not only displayed a better safety profile but also a greater uptake into the brain [9,40]. In contrast, high-affinity antibodies have a reduced brain uptake as they resulted in the lysosomal degradation of both the antibody and the receptor [41]. The highest amount of uptake by the brain was observed utilizing an antibody with a binding affinity of 111 ± 16 nM to TfR, while the lowest uptake was seen with the higher affinity antibody (1.7 ± 0.1 nM to TfR) [9]. Therefore, the TfRA4 aptamer characterized in this study, which possessed a moderate binding affinity (487.3 ± 108.8 nM), is likely to successfully transcytose across the BBB. Aptamers, which are identical to monoclonal antibodies in their ability to bind targets with a high degree of selectivity, can be functionalized without loss-of-function, and this truncated aptamer is currently being developed as a novel therapeutic for neurodegenerative disorders.
Footnotes
Acknowledgment
This work was supported by a Deakin University Central Research Grant, RM27686.
Author Disclosure Statement
No competing financial interests exist.