
Abstract
Thrombin-binding aptamers are promising anticoagulants. HD1 is a monomolecular antiparallel G-quadruplex with two G-quartets linked by three loops. Aptamer–thrombin interactions are mediated with two TT-loops that bind thrombin exosite I. Several cations were shown to be coordinated inside the G-quadruplex, including K+, Na+, NH4+, Ba2+, and Sr2+; on the contrary, Mn2+ was coordinated in the grooves, outside the G-quadruplex. K+ or Na+ coordination provides aptamer functional activity. The effect of other cations on aptamer functional activity has not yet been described, because of a lack of relevant tests. Interactions between aptamer HD1 and a series of cations were studied. A previously developed enzymatic method was applied to evaluate aptamer inhibitory activity. The structure–function correlation was studied using the characterization of G-quadruplex conformation by circular dichroism spectroscopy. K+ coordination provided the well-known high inhibitory activity of the aptamer, whereas Na+ coordination supported low activity. Although NH4+ coordination yielded a typical antiparallel G-quadruplex, no inhibitory activity was shown; a similar effect was observed for Ba2+ and Sr2+ coordination. Mn2+ coordination destabilized the G-quadruplex that drastically diminished aptamer inhibitory activity. Therefore, G-quadruplex existence per se is insufficient for aptamer inhibitory activity. To elicit the nature of these effects, we thoroughly analyzed nuclear magnetic resonance (NMR) and X-ray data on the structure of the HD1 G-quadruplex with various cations. The most reasonable explanation is that cation coordination changes the conformation of TT-loops, affecting thrombin binding and inhibition. HD1 counterparts, aptamers 31-TBA and NU172, behaved similarly with some distinctions. In 31-TBA, an additional duplex module stabilized antiparallel G-quadruplex conformation at high concentrations of divalent cations; whereas in NU172, a different sequence of loops in the G-quadruplex module provided an equilibrium of antiparallel and parallel G-quadruplexes that shifted with cation binding. In conclusion, structures of G-quadruplex aptamers are flexible enough and are fine-tuned with different cation coordination.
Introduction
A
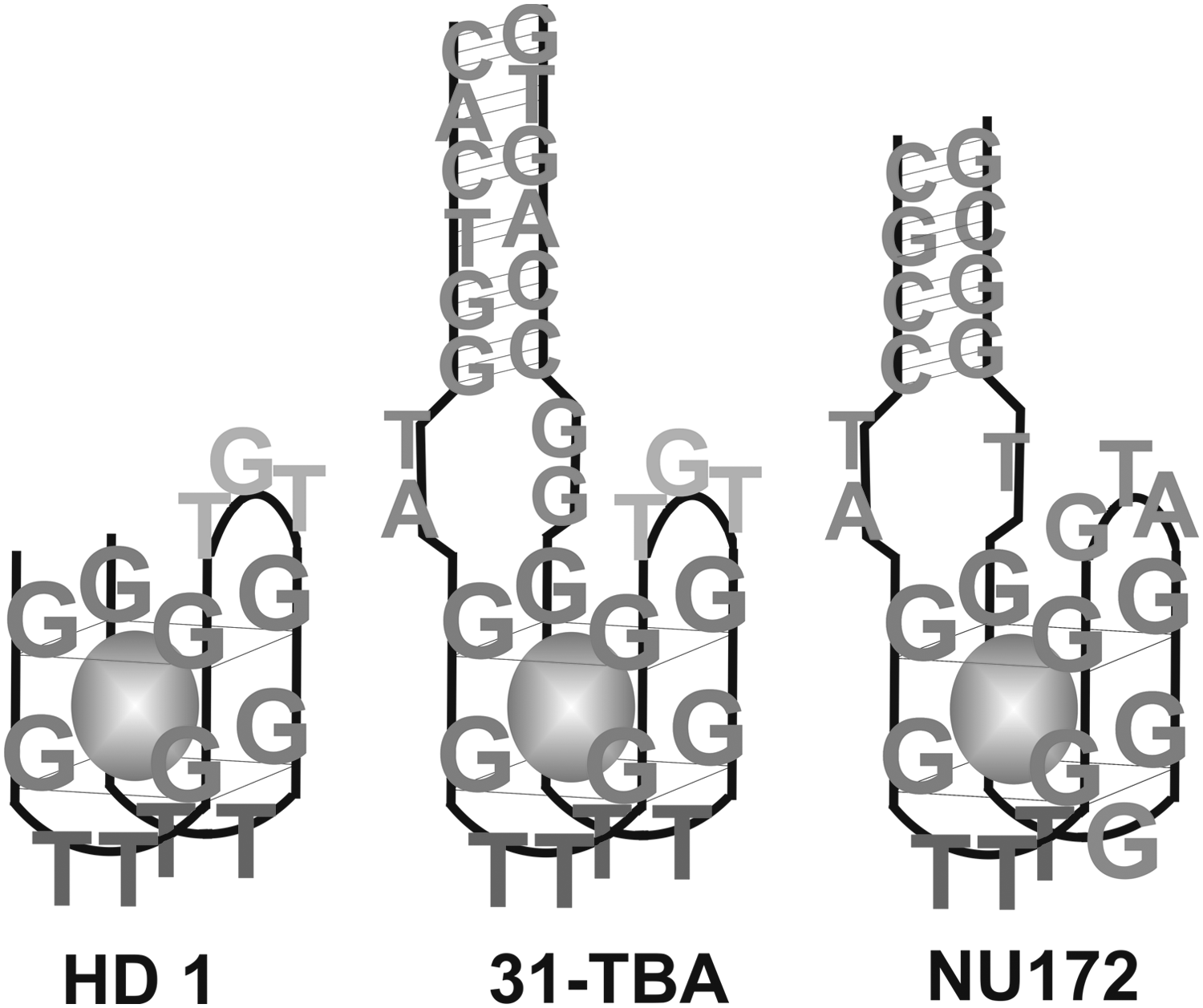
Schematic representation of the structure of aptamer HD1 and putative structures of aptamers 31-TBA and NU172. The potassium cation is shown as a sphere.
The structure of HD1 was resolved with nuclear magnetic resonance (NMR) [7–10] and X-ray analysis [11] both for the aptamer alone and for the aptamer–thrombin complex. HD1 is a minimal pharmacophore for thrombin, being a monomolecular antiparallel G-quadruplex with three lateral loops (Fig. 2). G-quartets are almost planar, being in stacking with each other and with some bases from the loops. The central TGT-loop partially stacks on the G-quartet; two bases, guanine and one of neighboring thymines, are in stacking with the G-quartet, whereas the remaining T does not interact with the G-quartet. Two TT-loops on the opposite side of the G-quadruplex interact with thrombin through hydrophobic and polar contacts. T4 and T13 are in stacking with the G-quartet, whereas T3 and T12 are unstacked and buried in the hydrophobic pockets of thrombin exosite I.

Structure of aptamer HD1 derived from the X-ray structure of the aptamer–thrombin complex (pdb id 4DII).
HD1 G-quadruplex assembly requires a cation that is coordinated with guanines and diminishes dipole repulsion. The aptamer complexes with potassium (K+) or sodium (Na+) cations are functionally active; the structures of HD1–thrombin complexes were determined, revealing one K+ or Na+ cation inside the G-quadruplex [11]. Several other cations were shown to be coordinated inside the G-quadruplex of aptamer HD1, in particular strontium (Sr2+) [10], barium (Ba2+) [12], lead (Pb2+) [13,14], and ammonium (NH4+) [15]. Manganese (II) (Mn2+) cations were shown to be coordinated with phosphates in the minor grooves of the G-quadruplex [9]. Contrary to K+ and Na+ cations, functional activity of HD1 with other cations has not yet been shown.
The goal of this research is to explore functional results of cation coordination, namely changes in the inhibitory activity of the aptamer measured by an enzymatic approach, whereas the structure of the aptamer is followed by circular dichroism (CD). The aptamers in the study are HD1 and its extended counterparts, 31-TBA and NU172, coordinated with Na+, K+, NH4+, Ba2+, and Mn2+. The study aimed at establishing a correlation between inhibitory activity and G-quadruplex existence, as well as the effect of the duplex module on the functional properties of the aptamers.
Materials and Methods
Inorganic salts and Tris were obtained from MP Biomedicals and Sigma-Aldrich. Human thrombin with a specific activity of 4.2 kIU/mg (HTI) and fibrinogen from human plasma (Calbiochem) were used. Oligonucleotides were synthesized by Evrogene and purified by precipitation in ethanol.
Aptamer preformation
Aptamer solution (400 nM for enzymatic tests or 5 μM for CD experiments) with 0–50 mM of cation chloride was heated at 95°C for 5 min, and it was then cooled at room temperature to assemble the G-quadruplex structure.
CD experiments
Preformed aptamer was diluted to 2 μM in a buffer (20 mM Tris-HCl, pH 7.5, 0.14 M NaCl, 1 mM CaCl2, varied content of the chloride of the cation studied) and placed in a quartz cuvette with an optical pass length of 10 mm. CD spectra were obtained with a Chirascan spectrometer (Applied Photophysics Ltd.) that was equipped with a thermoelectric temperature controller. The wavelength varied from 360 to 220 nm, the spectrum of the buffer was subtracted automatically, and the temperature was 20°C. Each experiment was performed two to three times and averaged.
Turbidimetric assay
The turbidimetric assay was previously described in detail [16]. Briefly, the reaction mixture with 2.0 μM fibrinogen in the buffer (20 mM Tris-HCl, pH 7.5, 0.14 M NaCl, 1 mM CaCl2) was placed into a UV-microcuvette (Brand), and the thrombin sample was added to start the reaction (final concentration 1–5 nM). The sample turbidity was detected at a wavelength of 230 nm by a WPA Biowave II+ spectrophotometer (Biochrom). All reagents were preheated at 37°C at least for 40 min. The reaction was carried out at 20°C. The inhibition coefficient was defined according to equation (1):
where
To study inhibition of thrombin with DNA aptamers, preformed aptamer (final concentration in the range 2–200 nM) was added into the assay straight before the thrombin. Each experiment was performed two to five times, and the results were averaged. Equation (2) was used to calculate the apparent inhibition constant:
where
Cations could affect the velocity of fibrinogen hydrolysis with thrombin. The most obvious effect was in the case of Mn2+ as they possibly accelerate fibrin association. To exclude the cation effect on the reaction, a sample with the same cation content but without the aptamer was used as a reference sample in each case.
Comparison of NMR and X-ray structures of HD1
All structures were aligned in PyMOL software using the “pair_fit” tool for guanosines of the G-quartets. The TT-loop conformations were compared in the following structures: (1) NMR of HD1 with K+ (pdb id 148D), (2) X-ray of thrombin–HD1 complex with K+ (pdb id 1HAO and 4DII), (3) X-ray of thrombin–HD1 complex with Na+ (pdb id 4DIH), (4) NMR of HD1 with Sr2+ (pdb id 1RDE), and (5) NMR of HD1 with Mn2+ (pdb id 1QDH).
Results
HD1
Both CD measurements and enzymatic assays were carried out in a buffer containing 140 mM NaCl, 1 mM calcium chloride, and a varied concentration of the cation under study. Buffer composition was partially originated from the enzymatic assay. Sodium cations are required for thrombin activity; calcium cations and the overall ionic strength are important for fibrin association.
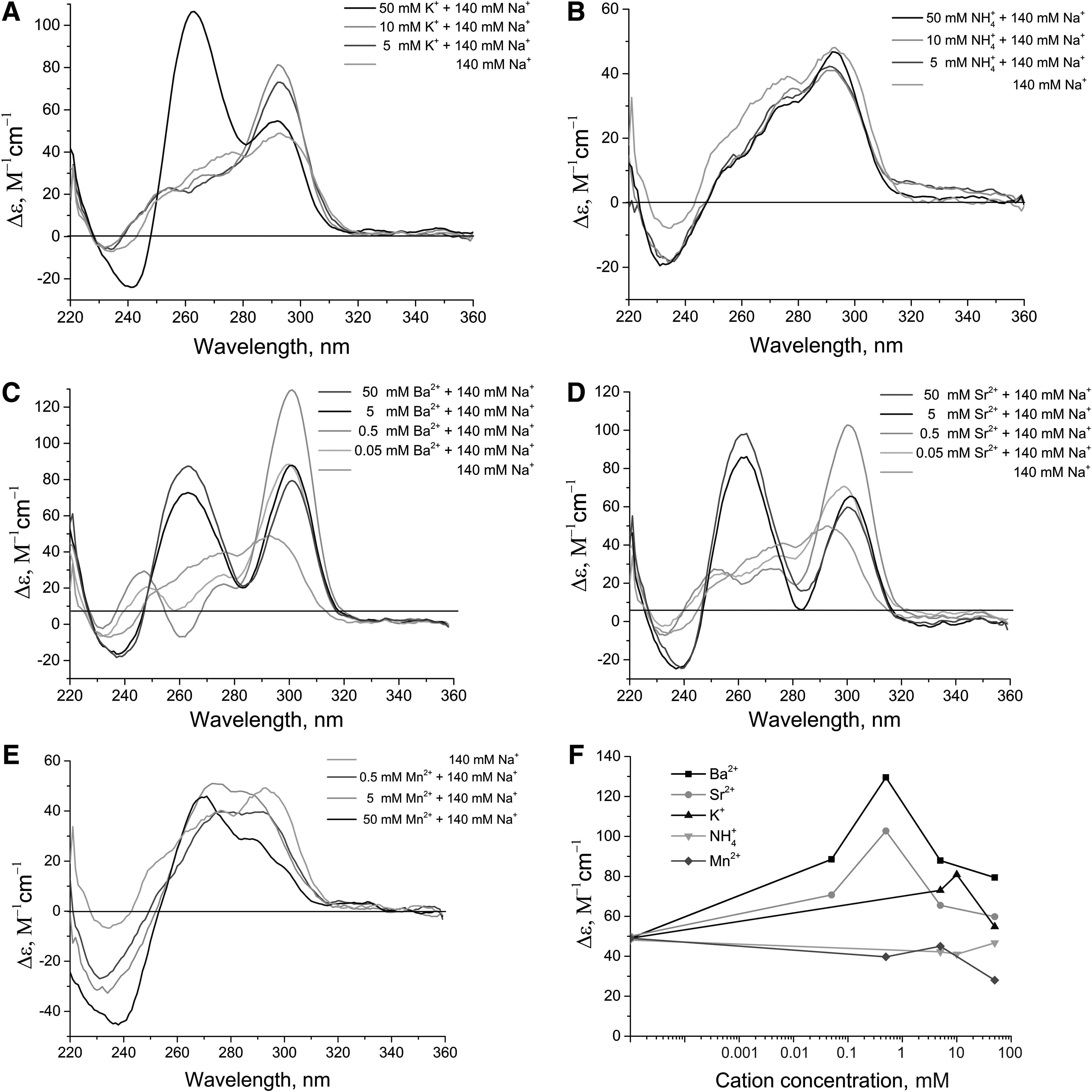
In 140 mM NaCl, the aptamer HD1 had a CD spectrum characteristic for an antiparallel G-quadruplex with two positive peaks, 251 and 294 nm, and a negative peak at 267 nm (Fig. 3). The apparent inhibition constant was 50 nM (Table 1).

Circular dichroism (CD) spectra for aptamer HD1 with cation concentrations in the range 0.05–50 mM:
The averaged values and standard deviations were determined using the turbidimetric assay, namely fibrinogen hydrolysis with thrombin in the reaction buffer with 140 mM NaCl.
K+ increased the intensity of both positive and negative peaks, keeping the same characteristic spectrum; the first positive peak was slightly shifted to 248 nm (Fig. 3A). A concentration-dependent shift of the spectra correlated with the decrease of the apparent inhibition constant down to 9.1 nM at 50 mM K+.
NH4+ provided a smaller increase in CD peak intensity compared with K+; the spectra are similar to those in 140 mM NaCl only (Fig. 3B). Apparent inhibition constants were increased in a concentration-dependent manner till complete disappearance of the aptamer inhibitory effect at 50 mM NH4+.
Ba2+ and Sr2+ greatly increased the intensity of peaks in CD spectra, changing some features. The second positive peak was shifted from 294 to 302 nm, whereas the negative peak at 267 nm was broadened to 267–280 nm, possibly a superposition of the 267 and 280 nm peaks (Fig. 3C, D). Both cations affected the HD1 spectrum, even at micromolar concentrations. Doubling of the intensity of the second positive peak was reached at 50 μM Ba2+ or Sr2+, whereas the same value was reached with K+ only at 50 mM, that is, by three orders of magnitude higher. At 50 mM concentration of Ba2+ or Sr2+, the second positive peak was considerably decreased (Fig. 3F); the overall spectrum was changed, possibly due to some aggregation processes. Apparent inhibition constants were increased up to 150–160 nM at 50 μM Ba2+ or Sr2+; a further increase of cation concentration completely abolished the inhibitory activity of aptamer HD1.
Mn2+ decreased the intensity of the second positive peak in a concentration-dependent manner; 0.5 mM Mn2+ shifted the peak to 297 nm, keeping the characteristic CD spectrum (Fig. 3E). The spectrum of the antiparallel G-quadruplex disappeared with the increase of Mn2+ concentration. Contrary to the cations mentioned earlier, these spectra did not contain isodichroic points, indicating the existence of multiple aptamer conformations. The apparent inhibition constants increased by 1.5 fold at 0.5 mM Mn2+; the inhibitory activity disappeared at Mn2+ concentrations higher than 5 mM.
31-TBA
Aptamer 31-TBA had CD spectra similar to aptamer HD1. In 140 mM NaCl, aptamer 31-TBA had a CD spectrum similar to the HD1 spectrum with an additional negative peak at 234 nm (Fig. 4). The apparent inhibition constant was 40 nM (Table 1).

CD spectra for aptamer 31-TBA with cation concentrations in the range 0.05–50 mM:
K+ increased the intensity of peaks at 294 and 270 nm only. Other peaks had the same intensity but were shifted: the positive peak to 249 nm and the negative peak to 231 nm (Fig. 4A). As in the case of HD1, a concentration-dependent increase of the 294 nm peak intensity correlated with the decrease of the apparent inhibition constant down to 2.1 nM for 50 mM K+ concentration.
NH4+ slightly affected CD spectra: The peak intensity increased just a little, and the peaks were not shifted (Fig. 4B). Similar to aptamer HD1, inhibitory activity of aptamer 31-TBA decreased gradually with the increase of NH4+ concentration up to the absence of inhibitory activity at 50 mM of the cation.
As in the case of aptamer HD1, Ba2+ and Sr2+ greatly increased intensity of the peaks at 294 and 270 nm, shifting the first one to 301 nm and broadening the second one to 265–280 nm (Fig. 4C, D). The positive peak was also shifted to 249 nm, but its intensity was changed without an isodichroic point. These features could be due to the influence of the additional hinge-duplex module in 31-TBA. Contrary to HD1, CD spectra of 31-TBA in 5 and 50 mM Ba2+/Sr2+ were nearly the same (Fig. 4F), possibly a result of the stabilizing effect of the extra duplex element. The apparent inhibition constant of 31-TBA was increased by 3.8 fold at 50 μM Ba2+ or Sr2+; a further increase in cation concentration led to the disappearance of inhibitory activity.
The addition of Mn2+ caused a gradual loss of the antiparallel G-quadruplex spectrum (Fig. 4E). The apparent inhibition constant was gradually increased, being twofold higher at 5 mM Mn2+; 50 mM Mn2+ concentration led to complete disappearance of the inhibitory activity of 31-TBA, which is 10 fold higher than that of HD1.
NU172
In 140 mM NaCl, the CD spectrum of the aptamer NU172 indicated no predominant G-quadruplex structure (Fig. 5). The spectrum could be considered a superposition of the antiparallel G-quadruplex with a positive peak at 294 nm, and some other conformations. The primary structure of NU172 differs from HD1 both in additional modules and in the loops of the G-quadruplex (Fig. 1).
CD spectra for aptamer NU172 with cation concentrations in the range 0.05–50 mM:
K+ concentrations of 5 and 10 mM increased the positive peak at 294 nm but had no effect on other parts of the spectrum, though 50 mM K+ changed the characteristic spectrum of the aptamer. In addition to the positive peak of the antiparallel G-quadruplex at 294 nm, the spectrum had a positive peak at 263 nm and a negative peak at 241 nm, characteristic of parallel G-quadruplexes (Fig. 5A). Therefore, a high concentration of K+ caused alteration of the topology of the G-quadruplex of NU172. Both the intensity of the 294 nm peak and the apparent inhibition constant were changed nonlinearly. When the K+ concentration increased from 0 to 10 mM, the inhibition constant decreased from 35 to 2.2 nM (Table 1); at 50 mM, the constant slightly increased to 3.5 nM, probably due to a partial rearrangement of the aptamer in the parallel G-quadruplex.
NH4+ had little or no effect on the CD spectrum of NU172 (Fig. 5B). The apparent inhibition constant increased gradually, and it became nonmeasurable at 50 mM NH4+.
Ba2+ and Sr2+ induced assembly of G-quadruplexes. The antiparallel G-quadruplex was predominant at 50 and 500 μM Ba2+/Sr2+; the best results were at 500 μM Ba2+, showing fine positive peaks at 301 and 247 nm as well as a negative peak at 260 nm (Fig. 5C, D). The spectrum also had a noncharacteristic positive peak at 275 nm that could be attributed to a different conformation of the aptamer. Ba2+/Sr2+ concentrations of 5 and 50 mM caused formation of both parallel and antiparallel G-quadruplexes. Two positive peaks were observed at 263 and 301 nm, being well separated in comparison to the similar case for K+. Apparent inhibition constants increased by 2–2.5 fold at 50 μM cations and by fourfold at 500 μM cations; at higher concentrations of Ba2+/Sr2+, the inhibitory activity disappeared.
Mn2+ changed the CD spectrum of NU172, yielding a mixture of several conformations (Fig. 5E). As in the cases of aptamers HD1 and 31-TBA, apparent inhibition constants of aptamer NU172 increased gradually with the increase of Mn2+ concentration, and inhibitory activity disappeared at 50 mM Mn2+.
Discussion
G-quadruplex folding versus inhibitory activity
Interaction between aptamer HD1 and various cations has been previously studied in detail using CD, NMR, UV-melting, isothermal titration calorimetry, differential scanning calorimetry, mass spectrometry, and so on [1,17]. Here, we have described the simultaneous measurements of aptamer conformation and inhibitory activity of the aptamer in the presence of different mono- and divalent cations, in an attempt to find a structure–function correlation.
As experiments on thrombin inhibition require a buffer with a high content of Na+, the first goal was to study the effect of the buffer on cation-dependent G-quadruplex formation. CD spectra of HD1–cation complexes in 140 mM NaCl were very similar to those without NaCl that were previously published [17,18]. This means that Na+ are weak coordination agents for HD1, having little or no effect on HD1–cation complexes.
The intensity of the positive peak at 294–301 nm, which reflects antiparallel G-quadruplex formation, was changed in the following order: Sr2+ = Ba2+ >>K+ >NH4+ >Na+ (Fig. 6) and corresponds to the raw thermal stability of HD1–cation complexes [18]. This raw thermal stability nearly agrees with the apparent dissociation constants of HD1–cation complexes: 0.009 mM for Ba2 + <<0.06–0.12 mM for K+ <0.45 mM for Na+ <5.4 mM for NH4+ [12,15,19]. Notably, 5 mM of lithium cations had no effect on the spectra of HD1 in 140 mM NaCl; the HD1–lithium cation complex is less stable than that with an Na+ [1] and, probably, does not exist in the sodium buffer.
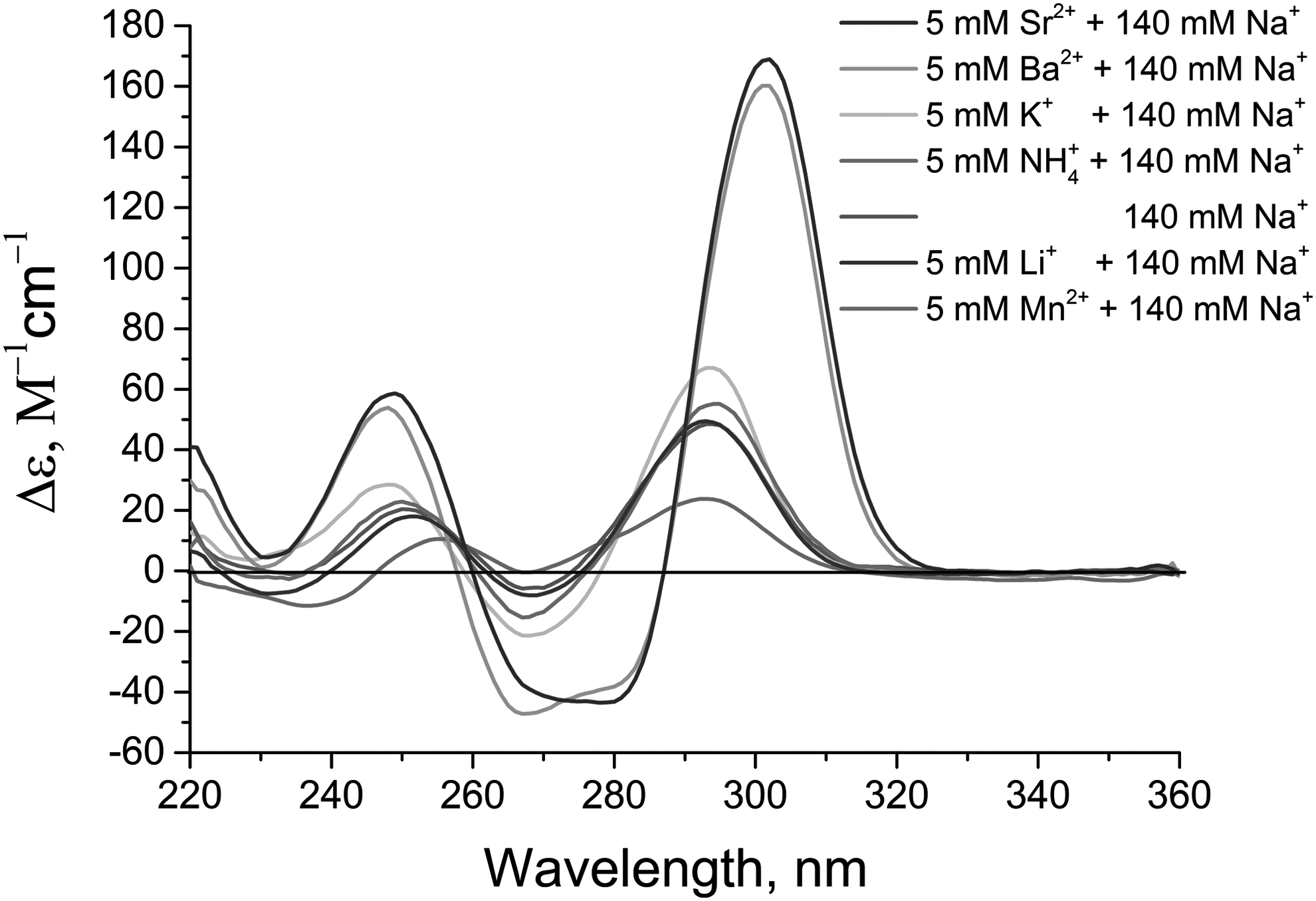
CD spectra for aptamer HD1 with cations at concentrations of 5 mM. Cation concentrations are listed in the order of curve height at 295 nM (from the highest to the lowest).
On the contrary, Mn2+ interact with HD1 aptamer under these conditions, decreasing the G-quadruplex CD peak twofold. Mn2+ were shown to be coordinated by phosphates of two minor grooves of HD1 but not by the guanines of the G-quadruplex [9]. NMR and an electron paramagnetic resonance study revealed that the cations rapidly exchange with the solution, and they dissociate under formation of the HD1–thrombin complex [9]. These peculiarities place Mn2+ aside from the other cations studied.
Surprisingly, HD1 inhibitory activity had no strict correlation with either CD spectra or the thermal stability of aptamer–cation complexes. The aptamer had low inhibitory activity in sodium buffer, having an apparent inhibition constant of 50 nM. K+ greatly increased the inhibitory activity in a concentration-dependent manner, reaching an apparent inhibition constant of 9.1 nM and, particularly for K+, the increase of peak intensity in CD correlated with the increase of the inhibitory activity of the aptamer. On the contrary, all other cations decreased aptamer inhibitory activity with an increase of the cation concentration. A reverse correlation can be found, as the activity decreased with the increase of the CD peak.
So, just the existence of the G-quadruplex itself is not enough for aptamer functional activity. As the interaction between thrombin and aptamer HD1 is mediated by two TT-loops, we compared their conformational state in different aptamer–cation complexes, as well as in aptamer–thrombin complexes.
The NMR-resolved structure of the HD1–K+ complex (pdb id 148D) was chosen as the reference structure. This complex was aligned with three X-ray structures of HD1–thrombin complexes (Fig. 7 A–C): pdb id 1HAO and 4DII–aptamer with coordinated potassium, pdb id 4DIH–aptamer with coordinated sodium. The TT-loops of all aligned structures are almost in the same plane, with some shift between the thymine centers. The only difference in the conformation of TT-loops between free HD1 and the HD1–thrombin complex is the T4–T13 pair; the pair is broken, and the T4–T13 distance is 3Å larger in the complexes with thrombin.
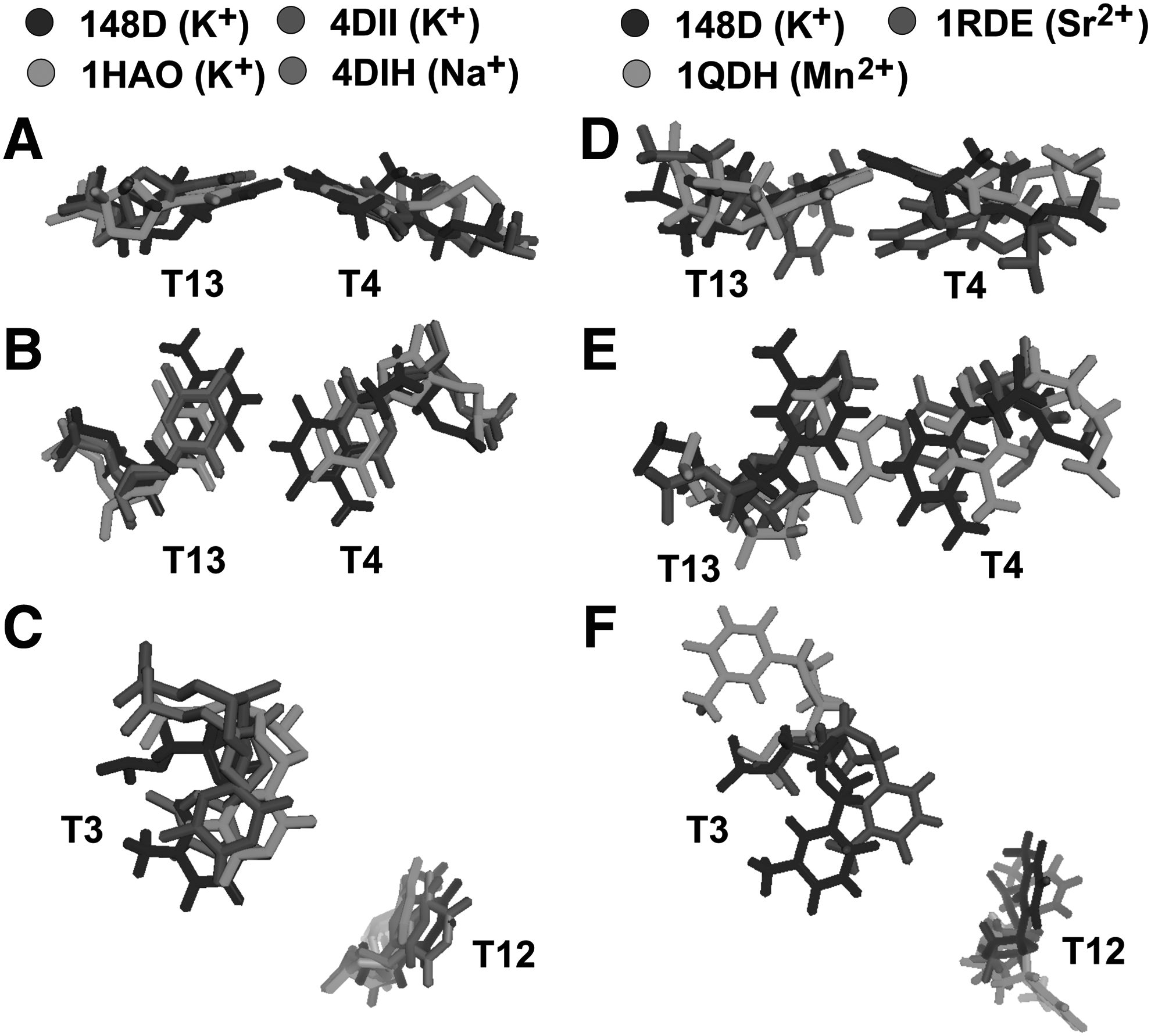
Alignment of the NMR structure of aptamer HD1–potassium cation with:
Further alignment of the NMR-resolved structures of complexes of HD1–Sr2+ (pdb id 1RDE) and HD1–Mn2+ (pdb id 1QDH) revealed that the conformation of TT-loops in these structures is significantly altered (Fig. 7 D–F). Sr2+ distorted the planarity of T4 and T13 bases, whereas Mn2+ declined T3 and T12 bases. (See Supplementary Table S2; Supplementary Data are available online at www.liebertpub.com/nat).
Keeping in mind that T4 and T13 participate in hydrophilic interactions with thrombin, whereas T3 and T12 are buried in the hydrophobic pockets of the protein, it is obvious that thymine conformations and their flexibility have a great impact on the formation of the aptamer–thrombin complex. How could cations affect the conformation of the loops? In fact, cation coordination by the G-quadruplex affects the structure, changing both the planarity of G-quartets and the rotation angle between two G-quartets (see Supplementary Table S1 for details). As TT-loops are short and ordered within the entire structure, alterations in G-quadruplex conformation unavoidably affect the conformation of TT-loops, exhibiting an example of allosteric interplay within the DNA aptamer.
Modifying HD1 with additional duplex and altered G-quadruplex loops
Addition of fragments with complementary sequences to the ends of the minimal G-quadruplex could yield an aptamer with the putative structure shown in Fig. 1. Dolinnaya et al. demonstrated coexistence of G-quadruplex and duplex modules in aptamer 31-TBA [20]. Just recently, it has been shown that an additional duplex module does exist in the thrombin complex with a homologous aptamer [21]. The additional duplex module could considerably increase the inhibitory activity of aptamer HD1 [5,22,23]. As no additional interactions between the duplex module and thrombin have been found in the thrombin complex with a homologous aptamer [21], the high inhibitory activity is probably a consequence of stabilization of the HD1 structure in a functionally active conformation.
The cations altered apparent inhibition constants similarly for both aptamers HD1 and 31-TBA. Inhibitory activity was increased with the increase of concentration of K+ (Table 1). Aptamer 31-TBA was a more efficient inhibitor of thrombin than aptamer HD1, which agrees with previous data [5,23].
CD spectra of aptamers HD1 and 31-TBA were mostly very similar to each other under all conditions studied (Figs. 3 and 4). Among the differences was a distinct negative peak at 234 nm in all 31-TBA spectra, which could be attributed to the contribution of the duplex module. Values of molar ellipticity of the peak were slightly lower in the case of 31-TBA than for HD1 with all cations studied. The most striking difference between the two aptamers was found at 50 mM Ba2+ or Sr2+ (Fig. 8B).

CD spectra for aptamers HD1, 31-TBA, and NU172 in 140 mM NaCl with cations at concentrations of 50 mM:
CD spectra of 31-TBA were nearly the same at 5 and 50 mM of the divalent cations (Fig. 4C, D), being similar to the spectra of aptamer HD1 at 5 mM of the cations (Fig. 3C, D); whereas the spectrum of HD1 was significantly distorted at 50 mM of the cations, possibly a result of some nonspecific interactions. The effect could be attributed to the stabilization role of the duplex module that fastens the ends of the G-quadruplex.
The stabilization effect of the additional duplex could also be demonstrated in the case of Mn2+. The spectra of HD1–Mn2+ had no isodichroic points (Fig. 3E), whereas the spectra of 31-TBA–Mn2+ contained common segments (Fig. 4E) that could be a result of the reduction of conformational diversity in the samples due to fastened G-quadruplex ends. Similarly, the inhibitory activity of aptamer 31-TBA was decreased with a higher concentration of Mn2+ than aptamer HD1.
Aptamer NU172 has a minimal G-quadruplex with altered loops that are flanked with the short complementary sequences at the ends (Fig. 1). The 3D structure of the aptamer has not yet been studied, so the putative structure was drawn simply by similarity with 31-TBA. Different sequences of G-quadruplex loops yield a greatly increased polymorphism of 3D structures of this aptamer. It seems to be that the main reason for that is a possibility of G-sliding within G-tracks, when guanines of GTA- and GT-loops could enter the G-quadruplex, forming altered antiparallel or even parallel G-quadruplexes. As a result, all the CD spectra of NU172 represent a mixture of several conformers, including an antiparallel G-quadruplex (Fig. 5). G-quadruplex formation was greatly promoted by adding cations that give the most stable complexes with G-quadruplexes—Ba2+, Sr2+, and, to a lesser extent, K+ (Fig. 5A, C, D).
In the case of NU172, the characteristic peak of the antiparallel G-quadruplex at 294 nm increased twofold in 10 mM K+, but it then decreased in 50 mM K+ due to partial rearrangement into a parallel G-quadruplex (Fig. 8A). Here, the inhibitory activity altered simultaneously with the height of the characteristic peak of the antiparallel G-quadruplex—it increased in 10 mM K+ and decreased in 50 mM K+. Ba2+ and Sr2+ promoted the formation of a parallel G-quadruplex even at 5 mM concentrations, possibly due to the formation of more stable complexes with G-quadruplex structures. So, alterations in G-quadruplex loops led to conformational flexibility of aptamer NU172. Among a variety of forms, the functionally active conformation is an antiparallel G-quadruplex that is coordinated with potassium or, to a lesser extent, sodium.
In conclusion, the data have clearly shown that the functional activity of particular aptamers to thrombin is provided by the conformational state of the TT-loops that interact with thrombin. Subtle changes in G-quadruplex conformation during cation binding affect the conformation of the TT-loops, which, in turn, greatly affects the inhibitory activity of the aptamers. Additional duplex modules in bipartite aptamers provide a stabilization effect by fastening the ends of the G-quadruplex. On the contrary, some alterations of loop structures of the G-quadruplex provide the additional possibility to fold into different conformers, in particular into a parallel G-quadruplex. All these properties have to be taken into account during the design of novel aptamers.
Footnotes
Acknowledgments
This work was supported by the Russian Science Foundation (grant no. 15-13-00033) in characterizing the conformational polymorphism of the G-quadruplex aptamer. This work was also supported by the Russian Foundation for Basic Research (grant nos. 14-04-01757_a and 16-03-00136_a) in characterizing the functional activity of the aptamers and analyzing the structure–activity correlations. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.