
Abstract
New advances in oligonucleotide (ON) chemistry emerge continuously, and over the last few years, several aspects of ON delivery have been improved. However, clear knowledge regarding how certain chemistries behave alone, or in combination with various delivery vectors, is limited. Moreover, characterization is frequently limited to a single reporter cell line and, when different cell types are studied, experiments are commonly not carried out under similar conditions, hampering comparative analysis. To address this, we have developed a small “tissue” library of new, stable, pLuc/705 splice-switching reporter cell lines (named HuH7_705, U-2 OS_705, C2C12_705, and Neuro-2a_705). Our data show that, indeed, the cell type used in activity screenings influences the efficiency of ONs of different chemistry (phosphorothioate with locked nucleic acid or 2′-O-methyl with or without N,N-diethyl-4-(4-nitronaphthalen-1-ylazo)-phenylamine). Likewise, the delivery method, Lipofectamine® 2000, PepFect14 nanoparticles, or “naked” uptake, also demonstrates cell-type-dependent outcomes. Taken together, these cell lines can potentially become useful tools for future in vitro evaluation of new nucleic acid-based oligomers as well as delivery compounds for splice-switching approaches and cell-specific therapies.
Introduction
W
The most used chemical modifications in splice-switching ONs (SSOs), or splice-correcting ONs, consist of either PS backbone plus one or several nucleotides carrying ribose sugar modifications such as 2′-O-methoxyethyl, constrained ethyl [7], LNA, 2′-OMe or modified sugar-phosphate backbone (eg, morpholino and peptide nucleic acid) [4,5]. Recently, the addition of the N,N-diethyl-4-(4-nitronaphthalen-1-ylazo)-phenylamine (ZEN™) modification, a nonbase modifier, to anionic ONs was found to increase SSO melting temperature and consequently affinity to the target, without significantly affecting the specificity [8,9].
While these chemistries, alone or in combination, provide SSOs with high affinity, nuclease resistance, and binding to plasma proteins, there are some aspects that still need to be addressed, such as induction of innate immune responses, number of off-target effects, and frequently poor uptake.
Parallel to the need of new ON chemistries, ON therapies should also address problems, such as degradation in biological fluids, passage across cellular barriers, and intracellular trafficking [10]. In vitro delivery of “naked” ONs is characterized by poor internalization in cells and requires high concentrations and long periods of cellular incubation [10–12]. Delivery vectors improve the internalization of ONs, change their subcellular distribution, thereby increasing their activity. Several cationic lipids [13], particulate systems (nanoparticles, block copolymers, microspheres), dendrimers [14], and polyethyleneimine derivatives [15], as well as cell penetrating peptides (CPPs) [16–19], among others, are available and have been used extensively [20]. However, as stated by Garcia-Chaumont et al. [20], comparative studies on the efficacy and toxicity of different formulations performed under the same experimental conditions in the presence of serum and with the use of various types of cells are needed.
On the contrary, for simplicity and reduced toxicity, in vivo is often attempted using “naked” intravenous administration of the ONs, and only a few chemical modifiers improve this delivery approach. While the systemic biodistribution is broad, higher concentrations of ONs are regularly found in clearance organs, including the liver and kidney, followed by bone marrow, adipocytes, and lymph nodes [21]. Unfortunately, in most diseases, there is limited correlation between observed biodistribution and drug accumulation in the desired target tissue, especially in the case of splicing defect diseases, where the tissues targets are often very difficult to access, such as the brain or muscle [2,22,23].
The first description of splice switching of the 705 mutation and the subsequent report describing the development of the pLuc/705 splice-switching reporter system [24,25] have been cited almost 400 times. While this reporter assay is the most used in the field, its applicability in screenings of new tissue-specific SSOs or vectors is hampered by the fact that, to our knowledge, only a few stable reporter cell lines exist, namely HeLa Luc/705 (cervical cancer cell line, and the most used) [25], PC3/Luc705 (prostate cancer cell line) [26], and A375SM-Luc705-B (melanoma cell line) [27]. To enhance tissue-specific uptake, there is a need for a greater variety of relevant reporter cell lines. In this report, we present newly developed tools for this purpose.
Materials and Methods
Splice-switching ONs
All the SSOs used in the present study were synthetized using standard phosphoramidite chemistry and used in sodium salt form (Integrated DNA Technologies). The SSOs possess fully modified PS backbone with one or more ribose sugar modifications (2′-OMe; LNA), with or without ZEN. Sequences and chemical modifications are listed in Table 1. After being dissolved in nuclease-free water, the concentration of the SSOs was measured using a NanoDrop (Thermo Scientific) and stored as frozen aliquots at −20°C.
ZEN, N, N-diethyl-4-4-(4-nitronaphthalen-1-ylazo)-phenylamine; PS, phosphorothioate; LNA, locked nucleic acid.
Cell lines and cell culture conditions
Human tumor cell lines of different tissue origins, hepatocyte HuH7, cervix HeLa Luc/705, bone U-2 OS cell lines, and the mouse myoblast C2C12, and the mouse neuroblastoma Neuro-2a cell lines were maintained and cultivated in Dulbecco's modified Eagle's medium (DMEM) with high glucose plus 10% fetal bovine serum (FBS) at 37°C, 5% CO2 in 95% humidity. Once established, the human HuH7_705, mouse myoblast C2C12_705, and Neuro-2a_705 cell lines were maintained in a medium supplemented with 400 μg/mL geneticin, while the human bone U-2 OS_705 cell line was maintained in a medium supplemented with 200 μg/mL geneticin.
Establishing HuH7_705, U-2 OS_705, C2C12_705, and Neuro-2a_705 stable monoclonal cell lines
All the plasmid constructs were assembled by Mutagenex, Inc. Initially, the pLuc/705 plasmid (a kind gift from Ryszard Kole) was digested with XmnI/CfrI ( = EaeI) and the fragment (4.35 kb) was inserted into the pT2-HB-CAG-MCS Sleeping Beauty transposon vector, kindly provided by Abdalla J. Mohamed (built on the transposon plasmid pT2/HB [Addgene plasmid No. 26557, a kind gift from Perry Hackett) and contains the multicloning site from plasmid pSL301 (Invitrogen)]. Ligation was performed with the PmlI/NotI vector fragment, 3.5 kb from pT2-HB-CAG-MCS. The new construct has a total size of ∼7.85 kb. In addition, the neomycin expression cassette from pEGFPLuc vector (Clontech) was inserted between the NcoI and EagI sites of the previous construct. This final construct with 9.45 kb is designated pT2Kole-Neo (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/nat). In both cases, the full insert was verified by DNA sequencing.
The cell lines were transfected with 1.5 μg of the pT2Kole-Neo together with 0.5 μg of the pCMV(CAT)T7-SB100 vector (Addgene plasmid No. 34879, a gift from Zsuzsanna Izsvák) [28] in a six-well plate with Lipofectamine® 2000 (Life Technologies) for HuH7 and U-2 OS cell lines, or Lipofectamine® 3000 (Life Technologies) for C2C12 and Neuro-2a cell lines following the manufacturer's protocol. Twenty-four hours posttransfection, the medium was removed and new medium was added. Forty-eight hours posttransfection, the medium was changed to a selection medium [DMEM plus 10% FBS supplemented with 800 μg/mL (HuH7, C2C12, and Neuro-2a cell lines) or 600 μg/mL (U-2 OS cell line) geneticin antibiotic (Life Technologies)]. The cultures were expanded for several cell passages and between day 14 or 20 posttransfection, cells were plated as single-cell cultures in 96-well plates to isolate monoclonal cell lines. Up to five monoclonal cultures of each cell line were selected, expanded, and evaluated in terms of endogenous versus SSO-induced splice correction at the RNA level as well as in luciferase protein activity. The clones with lower endogenous splice correction and higher SSO-induced splice correction were chosen for further studies.
SSO transfection with Lipofectamine 2000
Cells were seeded at a density of 7 × 104 (U-2 OS_705; C2C12_705 and HeLa Luc/705), 1 × 105 (HuH7_705), and 1.2 × 105 (Neuro-2a_705) cells per well in a 24-well plate the day before transfection, to reach ∼70%–80% confluency the next day. Transfection with different concentrations of the SSOs was performed with Lipofectamine 2000 (Life Technologies) in a ratio of 1 μg of SSO:2.5 μL of Lipofectamine 2000, formulated in OptiMEM for 20 min at room temperature and added to the cells in DMEM +10% FBS, according to the manufacturer's protocol. The complexes were left in the culture for 24 h, after which cells were harvested for RNA isolation or luciferase measurement.
SSO transfection with PepFect14
Cells were seeded at a density of 7 × 104 (U-2 OS_705; C2C12_705 and HeLa Luc/705), 1 × 105 (HuH7_705), and 1.2 × 105 (Neuro-2a_705) cells per well in a 24-well plate the day before transfection, to reach ∼70%–80% confluency at the time of transfection. Several concentrations of SSOs were mixed with PepFect14 at 5:1 molar ratios in nuclease-free water in 10% of the final treatment volume (ie, 50 μL). Nanocomplexes were formed for 30 min at room temperature. Before the addition of the complex to the cells, the medium in the wells was replaced with fresh serum containing DMEM (450 μL). The complexes were left in the culture for 24 h, after which cells were harvested for RNA isolation or luciferase measurement.
“Naked” uptake in Ca2+-enriched medium
Cells were seeded at a density of 8 × 103 (U-2 OS_705; C2C12_705; HuH7_705 and Neuro-2a_705) and 1 × 104 (HeLa Luc/705) cells per well in a 96-well plate or at 4 × 104 (U-2 OS_705; C2C12_705; HuH7_705 and Neuro-2a_705) and 5 × 104 (HeLa Luc/705) cells per well in a 24-well plate the day before treatment, to be approximately 40% confluent the next day. Medium was removed and a fresh medium supplemented with 9 mM of CaCl2, together with or without 0.5, 1, or 4 μM of the respective SSO, was added. After 48 h, another 50 or 100 μL, depending on the well size, of fresh normal medium was added and the cultures were incubated for another 24 h. Cells were harvested for RNA and luciferase measurements 72 h after the initial addition of the SSOs.
RNA expression analysis
Total RNA was isolated with Tri-Reagent® (Sigma-Aldrich) according to the manufacturer's protocol. Quantity and quality of the total RNA were determined by NanoDrop 2000 (Thermo Scientific) and the 260/280 nm and 260/230 nm ratios analyzed. Five hundred nanograms of total RNA was used for cDNA synthesis with the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems) according to the manufacturer's protocol. Reverse transcription polymerase chain reaction (RT-PCR) was performed with 12.5 ng of cDNA in each reaction (total volume per reaction was 25 μL) using the HotStarTaq Plus DNA polymerase kit (QIAGEN) following the manufacturer's protocol. Primers had the following sequences: Forward primer 5′-TTGATATGTGGATTTCGAGTCGTC-3′ and reverse primer 5′-TGTCAATCAGAGTGCTTTTGGCG-3′ (CyberGene); and the PCR program used was 5 min at 95°C; (30 s 95°C, 30 s 55°C, 30 s 72°C) for 29 cycles and 10 min at 72°C for final extension. The PCR products were analyzed in a 2% agarose gel in 0.5 × Tris/Borate/ethylenediaminetetraaceticacid (EDTA) buffer and visualized by SYBRsafe (Invitrogen) staining. Gel images were captured on a Fluor-S gel documentation system (Bio-Rad) with the Quantity One software (Bio-Rad).
Luciferase assay
To measure luciferase activity, the medium was removed, wells washed twice with 1 × phosphate-buffered saline (PBS), and cells lysed in 150 or 25 μL of 1 × PBS with 0.1% Triton-X 100 per well for 24- or 96-well plates, respectively. Cells were incubated for 20 min at 4°C, followed by a frost/defrost cycle at −80°C. After this, the lysates were kept on ice until use. Twenty microliters of the lysates were mixed by an injector with 100 μL of the luciferase assay reagent [29] (1 mM EDTA pH 8.0, 20 mM Tricine pH 7.8, 1 mM MgCO3 pH 7.8, 5 mM MgSO4, 25 mM 1,4-dithiothreitol, 1 mM adenosine 5′-triphosphate disodium salt hydrate, 25 μM Coenzyme A, and 1 mM
Data analysis
Data are expressed as mean ± standard error of the mean (SEM). Values were tested for normality by the D'Agostino–Pearson normality test (omnibus K2). Statistical significance was determined by one-way analysis of variance followed by comparison of each treatment with the group control by Fisher's least significant difference test (GraphPad Prism 6 Software; GraphPad Software, Inc.). In all cases, P < 0.05 was considered significant.
Results
Reporter cell lines
To promote research and development into splice-switching therapies, we decided to create four new reporter cell lines derived from muscle, neuron, liver, and bone cell lineages (C2C12, Neuro-2a, HuH7, and U-2 OS, respectively). We generated several monoclonal stable cell lines carrying the pLuc/705 splice-switching reporter [24,25]. It consists of a luciferase-encoding gene interrupted by a mutated β-globin intron 2. The existence of this mutation in the intron creates an aberrant 5′ splice site that activates a cryptic 3′ splice site, resulting in aberrant splicing of luciferase pre-mRNA and the translation of nonfunctional luciferase [25]. When an SSO masks the aberrant site, splicing is redirected generating the correct mRNA and consequently the luciferase activity is restored. The reporter vector used in this study was obtained by engineering the initial pLuc/705 construct developed by Kang et al. [25] and the full construct is described in Supplementary Fig. S1.
Figure 1 shows the expression of the luciferase mRNA in all cell lines. In contrast to the previous HeLa Luc/705 cells [25], a slightly higher endogenous level of correctly spliced mRNA is observed in the different cell lines (between 7.3 ± 2.4% and 18.0 ± 12.1% in the case of C2C12_705). However, and importantly, these values are similar over several passages of the cultures, demonstrating the robustness of these reporters (Supplementary Fig. S2). The total amount (by comparison of the gel band intensities) of luciferase mRNA expressed by these cell lines is also lower compared to the HeLa Luc/705 cell line, presumably as a consequence of the transposase-based approach used here, which limits the number of insertions into the genome. The morphology of the new reporter cell lines is similar to the parental cell lines, and their culture doubling times are 19 ± 0, 34 ± 3, 28 ± 3, and 21 ± 3 h for C2C12_705, U-2 OS_705, HuH7_705, and Neuro-2a_705, respectively.
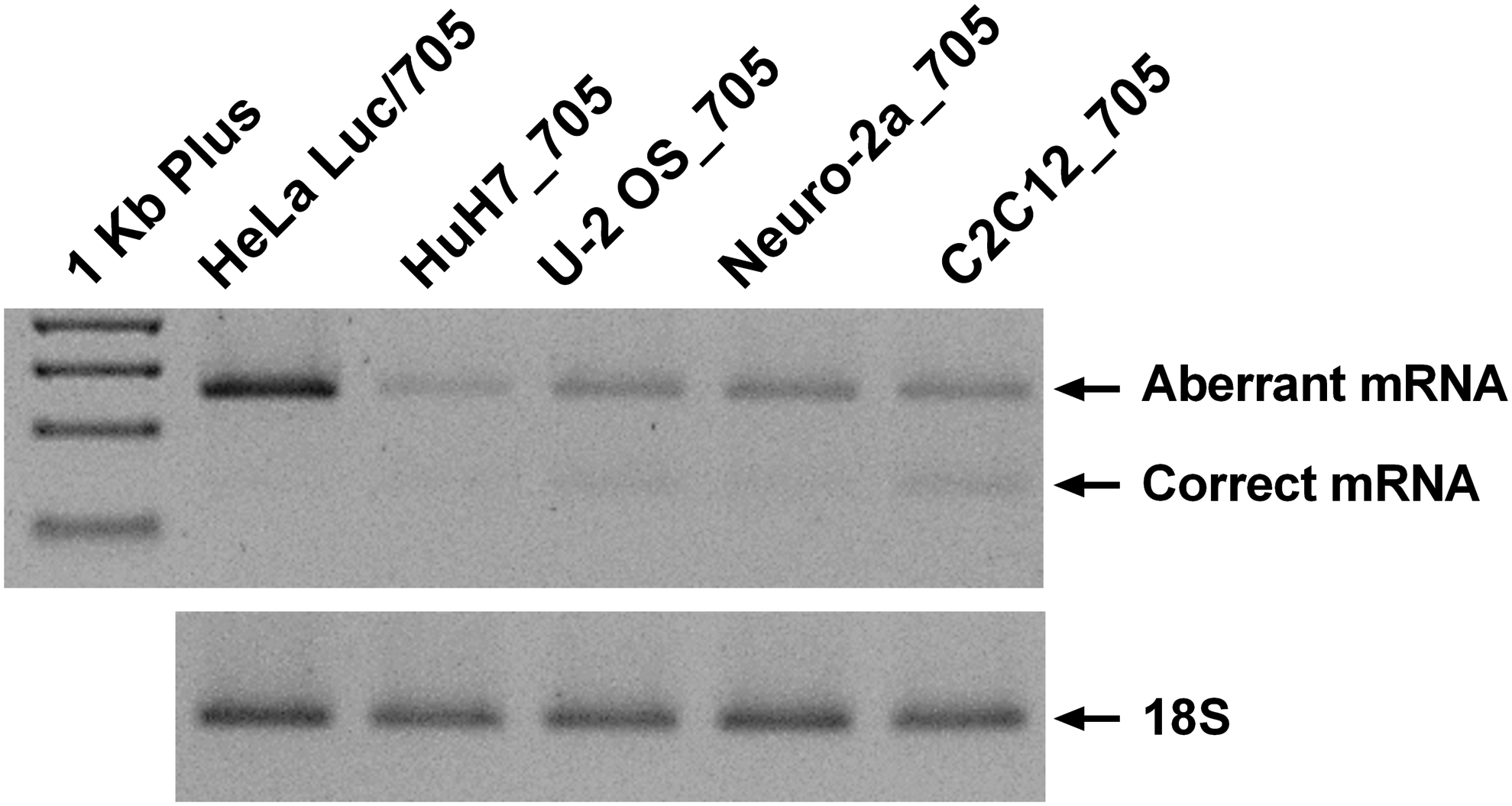
Expression of aberrant and correct luciferase mRNA and 18S in different reporter cell lines. Representative gel of reverse transcription polymerase chain reaction (RT-PCR) of luciferase mRNA and 18S RNA in all reporter cell lines without being subject to any treatment. Fragments were analyzed in 2% agarose gel with SYBR safe staining.
To evaluate if the influence of the ON chemistry and/or the delivery vector on the biological activity is cell-type dependent, all the new reporter cell lines together with the already existent HeLa Luc/705 reporter were either transfected with SSOs together with Lipofectamine 2000, treated with CPP nanoparticles, or assayed for “naked” ON uptake in Ca2+-enriched medium [30]. Three different SSOs possessing a fully modified PS backbone with one or several nucleotides carrying a ribose modification (2′-OMe; LNA) and, with or without, the ZEN modification were investigated (Table 1).
Splice correction mediated by Lipofectamine 2000
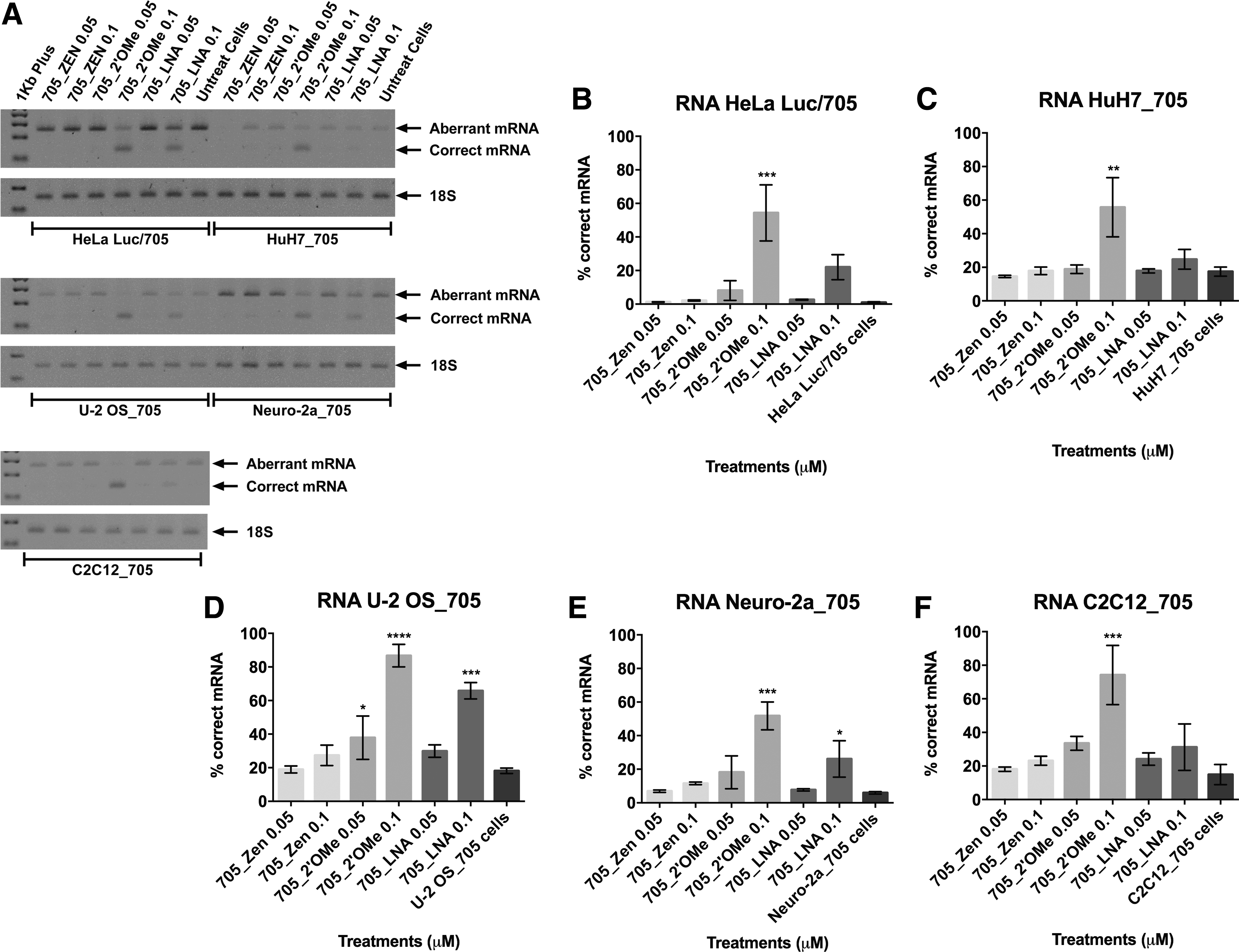
Transfection of the various SSOs with Lipofectamine 2000 induced splice correction in all cell lines in a dose-dependent manner for every chemistry tested. The results are presented in Figs. 2 and 3, where the corresponding percentages of correct mRNA are shown along with the protein levels, as measured by relative luminescence units (RLU) of luciferase, normalized to total protein. The 705_ZEN SSO performed exceptionally well in all cell lines, even at the lower concentrations, reaching in most cases almost 100% splice correction. Interestingly, cell-type specificity was observed between the 705_2′OMe and 705_LNA SSOs. The LNA substituted SSO was more effective than the 705_2′OMe at both concentrations tested in the

Efficiency of splice correction using different ON chemistries with Lipofectamine® 2000 is cell-type dependent. RT-PCR of aberrant (upper band) and correct (lower band) luciferase and 18S RNAs in all reporter cell lines when transfected at concentrations of 0.05 and 0.1 μM of the SSOs 705_ZEN, 705_2′OMe, or 705_LNA with Lipofectamine 2000 for 24 h. Fragments were analyzed in 2% agarose gel and a representative gel from three independent experiments is shown

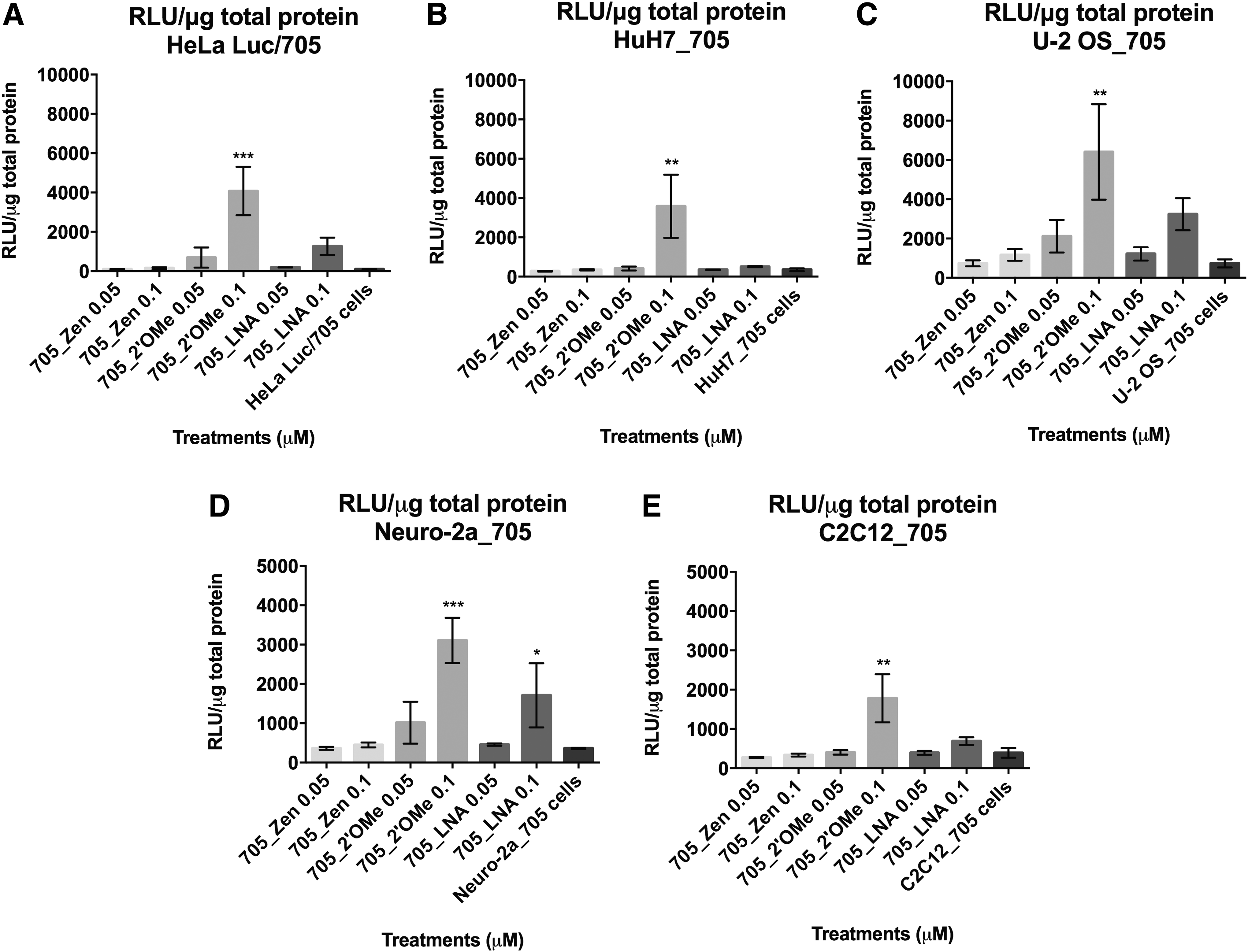
Luciferase production following splice correction with different ON chemistries using Lipofectamine 2000 is cell-type dependent. Graphs represent the RLU normalized by micrograms of total protein from HeLa Luc/705
When splice-corrected mRNA is translated, the same pattern of chemistry-related efficiency can be observed for the different SSOs in all cell lines (Fig. 3). However, statistical significance among treatments is only observed in cases where splice correction reaches values >50% or 60% depending on the cell line. No toxicity was found among the treatments in any of the cell lines and there was no significant variation observed in the particle size of the respective formulations (Supplementary Figs. S3 and S4, respectively).
Splice correction mediated by PepFect14 nanoparticles
PepFect14 is a stearylated CPP, derived from the stearyl-transportan10 [31] by replacement of lysines and isoleucines with ornithines and leucines. It was previously reported to be efficient for SSO delivery with low toxicity [16]. To elucidate if the type of delivery vector would also affect the activity differently depending on ON chemistry and cell line used, CPP nanoparticles were investigated. For the delivery by PepFect14 nanoparticles (Figs. 4 and 5), all SSOs were complexed with the same molar ratio 5:1 of PepFect14:SSO, regardless of the SSO concentration. This molar ratio was previously reported [16] as the best for PS 2′-OMe SSO delivery in HeLa Luc/705 cells and, hence, was our choice for evaluating the influence of the cell origin on the activity of the delivery vector, as well as changes in the nanoparticles depending on the SSO chemistry. As expected, a highly significant splice correction was observed with the 705_2′OMe SSO at 0.1 μM in all cell lines, with a maximum of 86.7 ± 6.7% in the
Splice correction upon delivery of different ON chemistries using PepFect14 nanoparticles is cell-type dependent. RT-PCR of aberrant (upper band) and correct (lower band) luciferase mRNA and 18S RNA in all reporter cell lines when concentrations of 0.05 and 0.1 μM of the SSOs 705_ZEN, 705_2′OMe, or 705_LNA were delivered in PepFect14 nanoparticles and the effect measured after 24 h. Fragments were analyzed in 2% agarose gel and a representative gel from three independent experiments is shown
Luciferase production following splice correction with different ON chemistries using PepFect14 nanoparticles is cell-type dependent. Graphs represent the RLU normalized by micrograms of total protein from HeLa Luc/705 (
Similar to Lipofectamine 2000, PepFect14 nanocomplexes also produced increased levels of the reporter protein with the same pattern of correlation to the various chemical modifications (Fig. 5). Similarly, statistical significance between treatments was only observed when splice correction reaches values >50% or 60%. Moreover, under these conditions, no toxicity was detected for any of the treatments and, furthermore, no variation in the particle size of the respective formulation was found (Supplementary Figs. S3 and S5, respectively)
Splice correction mediated by “naked” uptake in Ca2+-enriched medium
According to the protocol reported by Hori et al. [30], in which the medium is supplemented with CaCl2 to potentiate the ON effect at lower concentration and shorter incubation time in a gymnotic experiment, we decided to test our SSOs and reporter cell lines under these new conditions using 0.5, 1 and 4 μM for 72 h. The effects of this treatment on the biological activity are presented in Fig. 6.

Luciferase production following splice correction with different ON chemistries by “naked” delivery in Ca2+-enriched media is cell-type dependent. Graphs represent the RLU normalized by micrograms of total protein from HeLa Luc/705
The analyses demonstrate that
We also evaluated the pre-mRNA splice correction in the “naked” uptake experiments. While the HuH7_705 and U-2 OS_705 cell lines showed between 70% and 100% splice correction, use of the other cell lines resulting in values between 10% and 50% (data not shown). These splice correction percentages explain the low significance found in the luminescence reading for some cell lines since, as verified before, statistical significance was only achieved when cells reach between 50% and 60% of splice correction.
Considerable variability was observed between the “naked” uptake in Ca2+-enriched medium experiments (evidenced by high SEM values); however, the same efficiency pattern was consistently seen. While the cause of this variability is unknown, it was not due to toxicity as shown in Supplementary Fig. S6. Helmfors et al. [29] reported that the maximum readout of luciferase activity is dependent on the number of cells plated. We observed that even if the same number of cells were plated in the beginning of several independent experiments, at the end of the experiment the quantity of total protein obtained from cell lysates could vary. We noticed that luciferase values decreased with increased quantities of total protein, especially in adherent monolayer cell cultures, possibly due to cell contact inhibition (data not shown).
Discussion
In this report, we present four new reporter cell lines that enable screening of SSO chemical modifications, alone or in combination with delivery vectors, in a cell-type-specific context. To evaluate the applicability of the new stable cell lines, we analyzed the effect of three SSO chemistries, as well as three delivery approaches, which are well known and efficient in the field of splice-switching therapies. All SSOs in this study had a PS backbone, which improves biological stability, bioavailability, and cellular uptake [32–34], but also decreases the binding affinity to the target [6]. To increase target affinity, additional modifications such as 2′-OMe [6], LNA (with increased half-live in blood and thus potency, but also increased risk for off-target effects) [6,35,36], or ZEN (high target affinity but being less active in terms of “naked” uptake) [8,9] were included.
Previous reports demonstrated that a 705_2′-OMe SSO was efficient when delivered to HeLa Luc/705 cells using Lipofectamine 2000 [9,25] or PepFect14 [16], with no cytotoxic effects detected. Equally, Hammond et al. [9] show that a ZEN SSO was more efficient than a 705_LNA SSO when delivered through lipofection in HeLa Luc/705 and also here no cytotoxic effects were reported. Toxicity can be cell-type dependent, and thus, we evaluated the cytotoxicity for all the formulations used in this study with the WST-1 reagent. No adverse effects were observed in any of the cell lines (Supplementary Fig. S3).
SSO formulation efficiency could be dependent on the size and number of the particles formed. PepFect14 nanoparticles were only characterized and described for the 705_2′-OMe SSO [16]. Since different chemistries can formulate differently, particle size determination by nanoparticle tracking analysis was performed for all the formulations used here (Lipofectamine 2000 and PepFect14). No significant differences due to SSO chemistry were found between the size or number of particles for either delivery vector (Supplementary Figs. S4 and S5).
Based on the previous assessments, we would expect that since there is no variation in particle size and the formulations are not cytotoxic, the readout should be mainly dependent on SSO chemistry and, based on the HeLa Luc/705 data previously reported, the 705_ZEN SSO should be more potent than the 705_LNA SSO, followed by the 705_2′-OMe SSO.
However, our observations clearly showed that depending on the cell line, different ON chemistries in a lipoplex context or in PepFect14 nanoparticles showed different activities.
Comparison of lipid-assisted (Figs. 2 and 3) with peptide-mediated delivery (Figs. 4 and 5) showed that the 705_LNA SSO did not display any cell-type variation in its activity (better in both cases in U-2 OS_705, followed by HeLa Luc/705, Neuro-2a_705, C2C12_705, and HuH7_705). In contrast, a cell-type-specific pattern was found with the 705_2′OMe SSO. Both delivery approaches showed higher activity in U-2 OS_705; but then, for Lipofectamine 2000, the rank order of activity was HeLa Luc/705 > Neuro-2a_705 cell lines, while PepFect14 is best in the C2C12_705 cell line. Surprisingly, whereas the 705_ZEN SSO was the best reagent in all cell lines when delivered using the cationic lipid reagent, its activity was very low (U-2 OS_705 and C2C12_705) or negligible using PepFect14 nanoparticles. The reason for this variability remains unknown and requires further study.
While delivery vectors are very useful to assess the activity of new ON analogs in vitro, ON delivery in vivo remains a major challenge. In vivo experiments normally require large doses of ONs and, thus, preliminary screenings of chemistries in this context are rather expensive and often cannot be fully explored. To overcome this problem, in vitro experiments that can mimic or present a good correlation with the in vivo context are desired. During the last years, many efforts have been made to improve the understanding of “naked” uptake of ONs in vitro [37,38]. Recently, Hori et al. [30] demonstrated that it was possible to potentiate the ON effect in a gymnotic experiment and, importantly, a good correlation with the activity in vivo was observed. Supplementary Figure S6 shows that no cytotoxic effect is observed when the different reporter cell lines were treated with medium supplemented with calcium in the presence or absence of SSO, demonstrating the applicability of this method in these new cell lines.
Our data show that “naked” uptake of different chemistries is cell line dependent. The 705_LNA SSO seems to be more effective in cell lines considered as poor in “naked” uptake, such as HeLa Luc/705, Neuro-2a_705, and C2C12_705, while the 705_2′-OMe and 705_ZEN SSOs perform better in cells such as HuH7_705 and U-2 OS_705. Our data also corroborate the in vivo [9] findings for low activity of 705_ZEN in muscle cells upon naked systemic delivery of the SSO. Of notice is the low activity of SSOs such as 705_LNA and 705_ZEN in the HuH7_705 hepatocyte cell line, leading us to postulate that the higher activities normally observed for these SSOs in vivo in the liver could be a consequence of the high vascularization of this organ or the profound fenestration rather than a sign of cell specificity or other fundamental differences between primary hepatocytes and this laboratory-adapted cell line.
In addition, a “typical” gymnotic experiment (described in the “Materials and Methods” section but without calcium supplementation of the media) was performed and, although the same chemistry-related pattern among the reporter cell lines was found, the luminescence ratio values were very low, rendering it impossible to make a clear discrimination at lowest concentrations (data not shown). These findings reinforce the advantage of the calcium-enriched medium approach, supporting a correlation between the two gymnotic approaches regarding ON activity, as previously reported by Hori et al. [30]. More studies regarding how the different cell lines take up SSOs of different chemistries are needed, especially regarding the differential cellular expression and the role of endocytosis adaptors or cell receptors involved in the cellular uptake and intracellular trafficking of SSOs [10].
In conclusion, we demonstrate that the biological effect of ONs is dependent on the particular chemical modification as well as on the delivery vector, both activities being cell line dependent. Attending to the type of cell tissue selected to develop these new reporters, we believe that these new cell lines have high potential as models for cell-type-specific screening of different ON chemical modifications, transfection reagents, nanoparticles, delivery peptides, or aptamers or simply in the form of ”naked” uptake, for splice-switching therapies. In addition the biological effect can be easily evaluated by RT-PCR or by a Luciferase assay, allowing for high-throughput analysis. We hope to be able to make the cell lines available through an international cell repository.
Footnotes
Acknowledgments
The authors thank Professor Ryszard Kole, School of Pharmacy of the University of North Carolina, for the kind gift of the pLuc/705 plasmid and the HeLa Luc/705 cell line. The authors also thank Prof. Dr. Zsuzsanna Izsvák, Max-Delbrueck-Center for Molecular Medicine, Berlin, for the pCMV(CAT)T7-SB100 vector; Prof. Perry Hackett, College of Biological Sciences, University of Minnesota, for the pT2/HB vector; and Prof. Abdalla J. Mohamed, University Brunei Darussalam, Brunei Darussalam, for the pT2-HB-CAG-MCS Sleeping Beauty-transposon vector. This work was supported by the Swedish Medical Research Council. S.E.L.A. is supported by the Swedish Research Council and the Swedish Society of Medical Research (SSMF).
Author Disclosure Statement
M.A.B. is employed by IDT, which offers oligonucleotides for sale similar to some of the compounds described in the article. IDT is, however, not a publicly traded company and he personally does not own any shares/equity in IDT. No competing financial interests exist for the other authors.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.