
Abstract
Circulating tumor cells (CTCs) are cells that shed from a primary tumor and freely circulate in the blood, retaining the ability to initiate metastasis and form a secondary tumor in distant organs in the body. CTCs reflect the molecular profile of the primary tumor, therefore studying CTCs can allow for an understanding of the mechanism of metastasis, and an opportunity to monitor the prognosis of cancer. Unfortunately, the detection of CTCs is a considerable challenge due to their low abundance in the bloodstream and the lack of consistent markers present to recognize these cells. The aim of this review is to summarize some of the aptamer-based affinity methods for the detection of CTCs. The basic biological concept of how metastasis occurs and the role of CTCs in this process are presented. Some methods of CTC detection employing antibodies or peptides are mentioned here for comparison. The review of present literature suggests that aptamers are emerging as competitive technology in the detection of CTCs, especially due to their unique properties, but there still remain several challenges to be met, including the need to improve the throughput and sensitivity of such methods.
Introduction
C
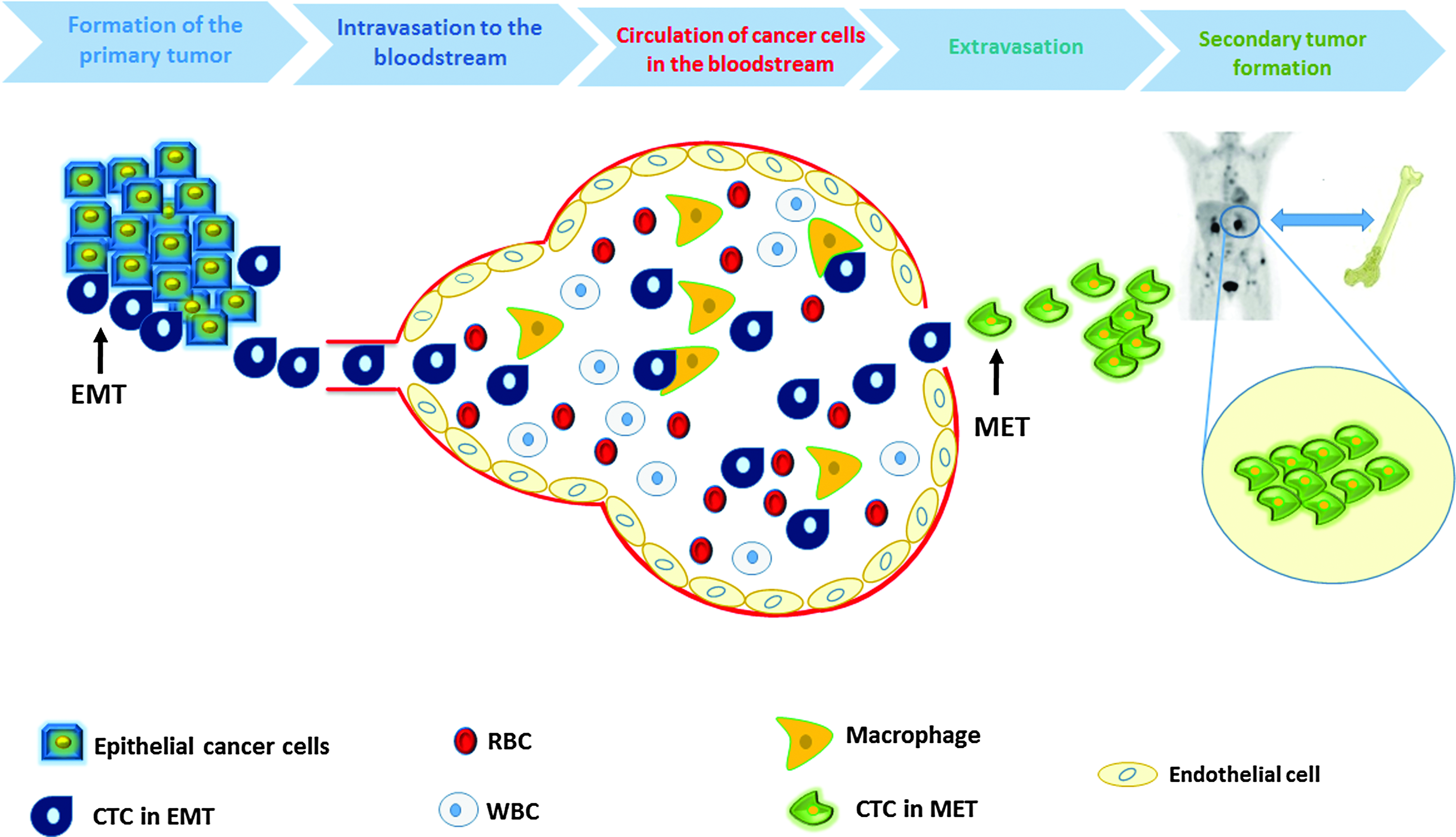
Metastasis cascade. L–R formation of primary tumor: cancer cells start to invade the local epithelial tissue and undergo the EMT. Intravasation and circulation: these cells enter the bloodstream with some of them eliminated by macrophages. The rest of CTCs will exit the blood in a process called extravasation. Secondary tumor formation: after they exit the blood, circulating cancer cells undergo a MET and start to invade distant organs forming a secondary tumor. CTC, circulating tumor cells; EMT, epithelial to mesenchymal transition; MET, mesenchymal to epithelial transition. Color images available online at www.liebertpub.com/nat
Metastasis and Formation of CTCs
Cancer is formed due to the accumulation of multiple mutations in the malignant cells, which lead to uncontrolled growth of the cells [7]. In general, mutant cells acquire the ability to independently synthesize or overexpress certain growth factors or proteins [7]. Moreover, the mutant cells are insensitive to inhibitory signals from signaling pathways [7]. Together, those mutations alter mitotic division resulting in abnormal cell division and elevated resistance to apoptosis (programmed cell death) leading to the formation of the solid tumor [7] (Fig. 1).
Cancer cells are heterogeneous and, thus, not all cancer cells are capable of initiating metastasis [8]. A small subset of CTCs gain stem cell-like properties (self-renewable, ability to initiate and sustain cancer growth) and thus have the ability to initiate metastasis through transformation from differentiated epithelial cancer cells into dedifferentiated cells that have more mesenchymal characteristics. This occurs via an epithelial to mesenchymal transition (EMT), which is considered to be a crucial event in the metastatic process [8,9] (Fig. 1). This facilitates their invasion into the bloodstream [7,8]. Downregulation of some of the cell adhesion proteins on the surface of the cancer cell (eg, E-cadherin), decreased expression of epithelial markers (eg, cytokeratin) [10], increased expression of mesenchymal markers (eg, vimentin), and change of the expression of the extracellular matrix (ECM) molecules (eg, integrin) are the most critical events in the EMT process [11–14]. The EMT cells break through the basement membrane, blood vessels, or interstitial spaces and start entering the circulation in a process called intravasation, allowing them to initiate distant or regional metastasis [15,16]. It is worth mentioning that intravasation could take place early in cancer development, even before the diagnosis of the primary tumor [17,18].
A solid tumor of one centimeter in diameter (∼109 cancer cells) could shed ∼1 million cancer cells into the circulation per day [19]; these cancer cells will be carried either by blood (CTCs) or by lymph [20]. CTCs do not spend long time circulating in the bloodstream. Indeed, most of them are collected by the first capillary bed that they encounter within minutes of entering the circulation, due their large size and inability to pass through capillaries [20]. It is well known that the majority of the secondary tumors are formed in organs that are not connected to lymph nodes (eg, brain, liver, lung, and bone) [21].
CTCs that are captured by the capillary beds may penetrate into the host organ through their parenchyma [22,23] and start to adapt to the new environment, obtaining nourishment by building new vessels through angiogenesis [24]. For those that succeed to adapt to the environment in the new organ, CTCs will undergo a mesenchymal epithelial transition in which the cancer cells will recover their epithelial markers [25,26]. The CTCs that make it to the new host organ and regain their proliferative properties are very few in number [27]; among the millions of cancer cells that shed every day from the primary tumor, only a small portion (∼0.1%) remain viable in the circulation after 24 h, and even fewer cells (0.01%) could initiate metastasis [28,29]. Most of the CTCs (85%) are lysed within a few hours of intravasation [30–32].
Current Methods of Detection of CTCs
Many methods have been established to detect CTCs in the blood of cancer patients. Some of them are based on the physical properties of CTCs, including size and density, along with immune and cell surface properties [33–35]. The above methods are not specific to capture CTCs and more specific techniques have been developed to detect CTCs with high selectivity and sensitivity [36–38].
Cellsearch® as the gold standard for the detection and quantification of CTCs
The Cellsearch® system was developed by Veridex and approved by the U.S. Food and Drug Administration as the only in vitro diagnostic system available in the clinic for detection of CTCs. Cellsearch provides sensitive detection and quantification of rare CTCs in human blood from breast, prostate, or colorectal cancer patients using fluorescence microscopy and immunology-based techniques [36–38].
Cellsearch is a semi-automated instrument. CTCs are enriched and separated from a 7.5 mL blood sample in the CellTracks AutoPrep system by antibody-mediated ferrofluid-based magnetic separation in which monoclonal antibodies against epithelial cell adhesion molecule (EpCAM) are conjugated to iron magnetic nanoparticles to specifically target CTCs. EpCAM is used as a marker to distinguish CTCs from blood cells. A second monoclonal antibody, anti-cytokeratin phycoerythrin (CK-PE), is used to stain CTCs followed by the DNA stain 4′,6-diamidino-2-phenylindole (DAPI). Blood leukocytes are separated from epithelial cells by using anti-CD45-allophycocyan (CD45-APC) [39]. Enriched CTCs are then incubated in the CellTracks Magnest where they will be analyzed using the CellTracks Analyzer II, a semi-automated fluorescence-based microscopy system that acquires images using a 10 × objective lens and filters corresponding to the different DAPI, PE, and APC stains [40]. A gallery of potential CTC images is then created. CTCs are selected by the user using qualitative analysis software and based on the differential staining mentioned earlier (CTCs are identified as EpCAM+, CK+, and CD45−) [39].
Challenges of Cellsearch in CTC detection
An evaluation of the morphology of CTCs can be obtained by the Cellsearch and quantification of the actual number of CTCs. However, the heterogeneity of CTCs, due to the diversity in their viability or apoptotic stage, make it difficult to set criteria for what counts as a CTC [41]. One of the limitations of the Cellsearch is the anti-EpCAM antibody-based enrichment strategy; as mentioned earlier, the expression of the epithelial markers decrease during the EMT process; therefore the expression of EpCAM is not the same among all CTCs. It has been reported that EpCAM expression changes in mammary carcinomas [42], and is downregulated in CTCs in peripheral blood [43]. Cellsearch results can carry the risk of false positives due to the use of epithelial markers, like cytokeratins, which are not tissue type-specific nor have the capability to distinguish the origin of epithelial cells. For example, some leukocytes can express epithelial markers in response to cytokine activation [44]. It has been reported that Cellsearch may undercount the number of CTCs [45]. CTCs are rare events in blood and 7.5 mL of blood, as a sample, may not reflect the entire cell population [46].
Recently, with the development of more sensitive and accurate detection methods, the CTC counts might be evaluated using smaller blood volumes (1 mL of whole blood instead of 7.5 mL). For example, most microfluidic systems require small blood samples (ranging between 1 and 4 mL, depending on the type of micro device) and show better CTCs recovery and enrichment rates than the Cellsearch method [47–51].
Relevance of Aptamers as a Detection Probe to Detect CTCs
Antibodies are well known as recognition elements, and therefore they are the main ligands in use for the detection of CTCs. However, the availability and the cross-reactivity of antibodies may limit their use in CTC detection [52]. Peptides tend to be advantageous compared to whole antibodies, due to their small size, and the ability to be conjugated to oligonucleotides or other functional group by chemical synthesis. Therefore, peptides have also been used as targeting ligands in the detection of CTCs [53,54]. Aptamers have shown potential as alternative ligands over both antibodies and peptides in the detection of CTCs, due to their lower cost and stability, allowing them to be used to detect CTCs under a wide range of conditions [52,55,56].
Herein, aptamers are being reviewed as powerful tools for the detection of CTCs. The different methods by which aptamers have been selected and integrated in the detection of CTCs are described. Finally, the future prospects of utilizing aptamers in the detection of CTCs are addressed.
Aptamers have first been described in 1990 as a new class of receptor molecules [57]. They are single stranded, short, DNA or RNA oligonucleotides (15–100 nt) that bind to their targets with high affinity and selectivity with a unique three dimensional interaction. Their discovery is facilitated by an in vitro screening method called SELEX (Systematic Evolution of Ligands by EXponential enrichment). Aptamers, unlike antibodies, are not limited to immunogenic or nontoxic targets. They can be made against small molecules (eg, amino acid, toxins), proteins, bacteria, and viruses. Aptamers are more stable in a variety of environmental conditions such as change in pH, temperatures, redox, and under nonphysiological conditions [58]. Aptamers can be easily synthesized and highly purified in the lab without the need for animals and without batch-to-batch variations [58]. Modification of aptamers can be done rationally and homogeneously, without a loss of affinity to the target, and thus can be used in a variety of diagnostic and therapeutic applications for CTCs [59].
Current Methods of Aptamer Selection
The overexpression of the surface biomarkers is often related to tumorigenesis [60]. Thus, about 60% of cancer-targeting drugs, including therapeutic antibodies and small molecule inhibitors, target cell surface biomarkers, making them the most attractive targets for cancer diagnosis and treatment [60,61]. Aptamers are generated by a variety of methods based on SELEX [62]. The most relevant approach to aptamer selection in the field of cancer research, however, is cell-SELEX as it is considered to be a powerful tool for the identification of new biomarkers [61]. In cell-SELEX, the selection is based on the native conformation of the surface proteins under physiological conditions, improving the chances that the aptamers will bind under physiological conditions (pH, temperature, etc.) [63]. Moreover, multiple aptamers can be generated for several protein receptors on the surface of the cell, enabling the identification of the molecular profile for the cancer cells, which is vital for diagnosis and treatment [60–63].
Whole Cell-SELEX using cell cultures and tissue as targets
Cell-SELEX starts with a large (1013–1015 ssDNA or ssRNA) randomized oligonucleotide library. The library is incubated with target cancer cells (positive selection) in the first step. Unbound DNA is washed away in the partitioning step, while bound DNA is eluted, then a counter selection step takes place with the eluted DNA being incubated with a similar noncancerous cell type as a counter target (eg, breast cancer cells vs. normal healthy breast cells [62]) followed by amplification of the unbound counter selection DNA. The cycle of selection is then repeated (Fig. 2). Changing the conditions of selection throughout the cycles (such as ionic strength and number of washes to remove the weak binders) is crucial to increase stringency and enrichment, and to yield selective, high affinity binders to the target [64]. The same procedure can be applied in situ on clinical tissue samples, in which the DNA library is incubated with a tissue slide as the target and the rest of the steps applied in the same way as mentioned above [65].
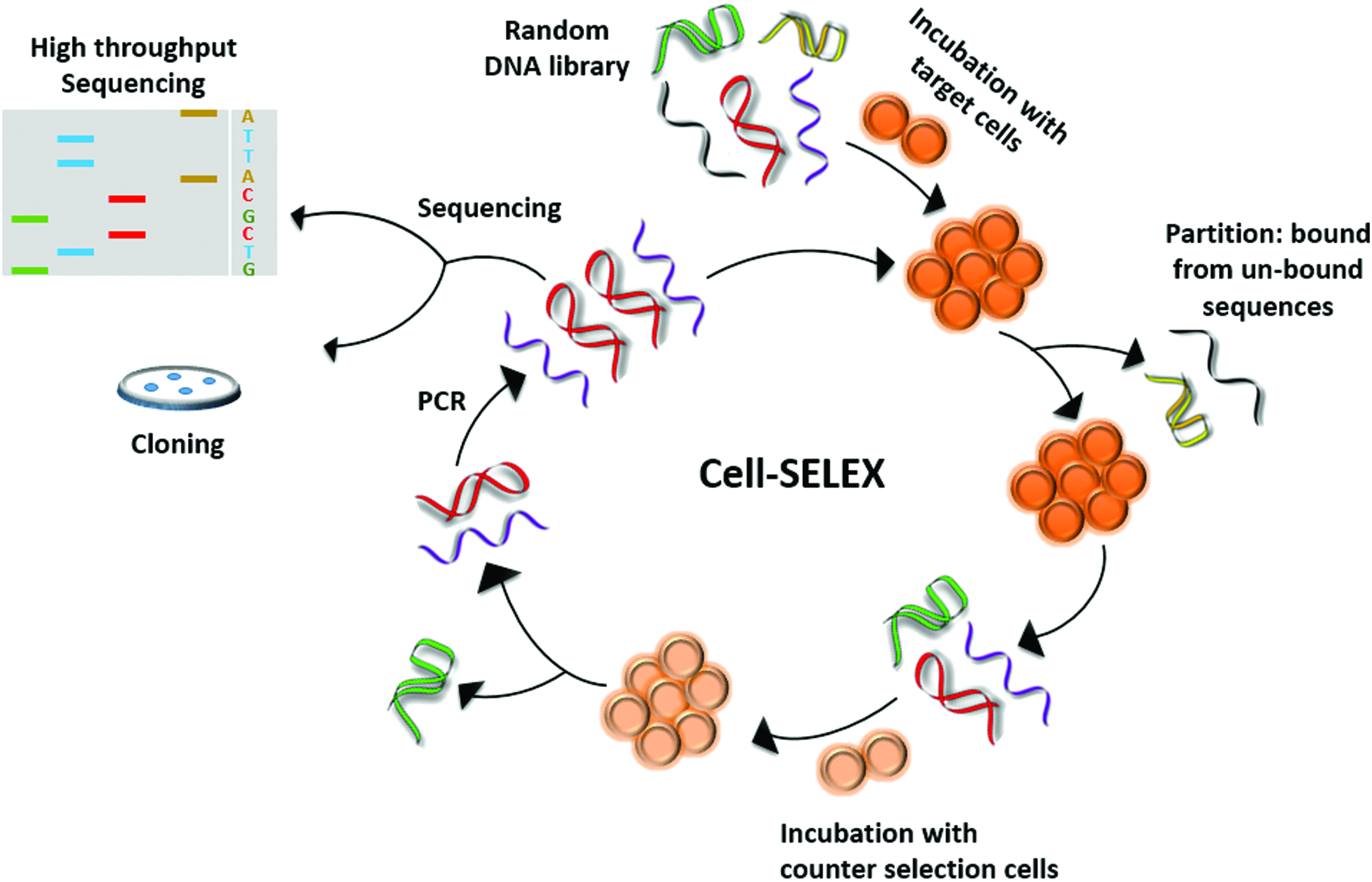
Cell-SELEX: Cell-SELEX starts with the incubation of a particular target cell with a random DNA library (positive selection). Unbound sequences are washed off and what remains is eluted off the cells (eg, by boiling). The eluted sequences are exposed to the counter selection cell line (to ensure that the resulting aptamers are selective). Sequences without affinity to the counter cells are amplified by PCR and cycle is repeated between 5 and 15 times. The final library is sequenced to identify aptamers candidates either by traditional cloning or high-throughput sequencing. PCR, polymerase chain reaction. Color images available online at www.liebertpub.com/nat
Whole cell-SELEX is of great benefit as the selection takes place on the native structure of the target proteins [66]. However, the presence of large numbers of different cell surface molecules and target biomarker(s) may lead to the selection of many unwanted aptamers [65]. Applying in situ SELEX is closer to the real environment of the disease (especially in the case of cancer) as the cells are still intact and not affected by in vitro factors as is the case of cell culture SELEX [65]. Moreover, the identification and characterization of new aptamers using the tissue as the target provides more information about the molecular mechanisms of cancer that might be different among individuals, thus using tissue as target is a step toward personalized medicine to improve the outcome of cancer patients [65].
Hybrid SELEX using recombinant proteins and cells as targets
Purified recombinant proteins can be used as targets for SELEX [58]. Post-translational modifications, however, add to the complexity of the use of recombinant proteins as aptamer targets, especially in the case of highly glycosylated proteins. Recombinant proteins or peptides often cannot fold into the correct three-dimensional structure that is formed under physiological conditions [67], and therefore the selected aptamers may not be able to bind specifically to their corresponding targets on the surface of the cell [67]. Since many unwanted aptamers are often generated using whole cell-SELEX, a hybrid (or cross-over) SELEX was developed using both recombinant proteins and cells as targets [68]. The SELEX selection alternates between the cells and the recombinant protein of interest. This will improve the affinity of the library of aptamers for the desired target and discern ones with higher selectivity toward a specific cell surface protein compared to whole cell-SELEX [68].
Ligand guided selection using cells as target and antibodies as epitope predetermination
Recently, ligand-guided selection, a modified cell-SELEX method, has been developed [69]. In this method, a strong secondary high affinity ligand (in this case, an antibody) is introduced in the partitioning step to compete with the enriched library on the same target [69]. Aptamers that have low affinity to the target will be eliminated from the selection (by being out competed) resulting in highly specific aptamers against the target [69]. In this particular study, aptamers against membrane-bound Immunoglobulin M (mIgM), expressed on Burkitt's lymphoma cells, were selected by adding an antibody for the same target to outcompete the low affinity aptamers in the library [69]. This new method allows for the selection of predetermined targets (epitopes), resulting in the generation of very specific aptamers against those epitopes [69]. This method will have a great benefit in the selection of new biomarkers for CTCs, thus leading to improve the detection of CTCs by creating very specific aptamers against predetermined targets.
Most of the aptamers that target tumor biomarkers are used in the diagnosis and therapy of solid tumors, and in the detection of CTCs. These are selected using purified recombinant proteins, or cell lysates that contain overexpressed recombinant target proteins, as targets [70–95]. The integration of the different types of cell-SELEX in the selection process will increase the specificity of CTC detection and therapy using aptamers. Table 1 summarizes the aptamers that have been currently selected against tumor biomarkers, along with their applications and methods of selection.
CEA, carcinoembryonic antigen; CTC, circulating tumor cell; ECM, extracellular matrix; EGFR, epidermal growth factor receptor; EpCAM, epithelial cell adhesion molecule; hnRNP A1, heterogeneous nuclear ribonucleoprotein A1; MUC-1, Mucin-1; OPN, osteopontin; PSMA, prostate-specific membrane antigen.
As seen in Table 1, EpCAM [70–72] and epidermal growth factor receptor (EGFR) [78–80] are the most common biomarkers for which aptamers have been employed for targeting CTCs. There are a number of aptamers selected against these two biomarkers, and most research groups use them as ligands for detection [70–72,78–80]. In a tissue slide-based SELEX method, Shaohua et al. developed an aptamer against heterogeneous nuclear ribo-nucleoprotein A1 (hnRNP A1) in which they prepared slides coated with cancer tissues and used it as target for SELEX [65]. Later, the same group used their system for the detection of CTCs from pancreatic cancer patients [85]. Another CTC biomarker is human periostin ECM protein, which is found in different types of metastatic cancers such as breast and gastric. For this reason, an aptamer was developed to detect CTCs in blood by recognizing this biomarker [91–95].
Detection of CTCs Using Single-Target Aptamer-Based Prototypes
Because of their small physical dimensions, microfluidic systems have shown great promise for establishing inexpensive, simple, and rapid tests for many different biological applications; one of which is capturing CTCs from the peripheral blood of cancer patients [96,97]. Since these devices are easily manufactured and portable, they can be widely utilized for clinical purposes. Furthermore, small samples and reagent volumes are utilized, and short sample preparation times allow for rapid CTC capture [98–100].
The design of such systems mainly depends on cell size, fluorescent or magnetic tagging of cells, and surface adhesion [101]. The ligand for a specific cell receptor is immobilized onto the surface of a channel to achieve cell capture from a flowing suspension [99]. Many different cell types have been captured using this strategy, such as lymphocyte subpopulations, breast cancer cells, lung cancer cells, and endothelial cells [98,102].
Several biomarkers have been used for capturing CTCs, among which EpCAM and different subtypes of cytokeratins (CKs) are, by far, the most frequently used markers. The EpCAM antibody has been used in many microfluidic device systems [103–108] as the ligand for capturing CTCs. The methods utilized vary in terms of physical dimension of the device, assay performance characteristics such as sample volume, sample processing time, assay preparation time, clinical sensitivity, clinical specificity, purity, recovery of the cells and other factors. The antibody-based micro devices typically require longer sample processing times but show higher purity than the CellSearch system [109].
Aptamers can rival antibodies for specificity and affinity for the target [110]. Therefore, they are good candidate probes for capturing CTCs in the bloodstream [69,111]. In microfluidic chip-based systems, aptamers have advantages over antibodies in that they are more easily labeled and more stable for longer periods under various conditions (pH, urea, organic solvents, and detergents) [112]. Aptamers that have been recently used in microfluidic devices are listed in Table 2.
ALL, acute lymphoblastic leukemia; GBM, glioblastoma; NSCLC, nonsmall cell lung cancer.
As Table 2 shows, one of the first aptamers that were integrated into a microfluidic device was an RNA aptamer against prostate-specific membrane antigen (PMSA), which showed highly efficient isolation and enumeration of rare circulating prostate tumor cells from whole blood samples with high recovery rate of CTCs [113]. Enrichment and detection of several rare cancer cells was established in a poly dimethylsiloxane (PDMS) microfluidic channel mentioned in Table 3 [116]. Briefly, DNA aptamers against different leukemia cell lines were immobilized on the surface of the micro device and CTCs were enriched and captured with high purity (83% ± 9%, 61% ± 14%, and 50% ± 10% according to the type of the cancer cell). Moreover, CTCs were released and cultured for further studies [116]. In an interesting example of maximal capture efficiency of CTCs, a three-dimensional multivalent DNA aptamer-based microfluidic device system was established by Zhao et al. (2012), in which a DNA aptamer against protein tyrosine kinase 7 (PTK7) was encoded into a circular template to produce single repeating adhesive aptamer domains by rolling circle amplification (Fig. 3). This ssDNA aptamer extended over tens of micrometers into the solution. Unlike monovalent microfluidic devices that tend to capture cells in low shear stress, the multivalent device captured cells at higher shear stress, permitting them to process higher volumes of blood more rapidly than monovalent devices. Moreover, the aptamer could be cleaved by restriction enzymes, allowing the captured cells to be easily released for molecular analysis [120].

Schematic of a multivalent DNA aptamer-based microfluidic device used to detect and isolate CTCs. The target CTC is captured by binding to a long DNA polymer containing multiple aptamer domains. Restriction enzymes are used to cleave the long DNA aptamers to release CTCs for further analysis. (Adapted from 107). Color images available online at www.liebertpub.com/nat
ANSB, Aptamer-gold nanoparticle strip biosensor; LDI-MS, laser desorption/ionization mass spectrometry; UCNP, upconversion nanoparticles.
Due to several advantageous properties, nanoparticles (NPs) have been used extensively in the detection of solid tumors and CTCs [121–124]. NPs are small in size (1–100 nm) so they tend not to trigger an immune response (useful for in vivo applications) [125]. Since functional groups can be attached to them, NPs can be coated with different types of ligands, such as antibodies, enzymes, and DNA to improve their ability to bind to a specific marker and increase the binding signal. Moreover, NPs have high surface-to-volume ratio, resulting in higher binding capacity and capture efficiency than other types of scaffolds [125].
Aptamers can be incorporated into NP-based detection systems for capturing CTCs. Aptamers have low molecular weight, which makes it easy to conjugate multiple aptamers on the surface of NPs. Also, aptamers are more easily modified than antibodies with different functional groups for conjugation purposes. Lastly, aptamers are cheaper than antibodies, which reduce the overall cost of the detection system [126]. Table 3 summarizes some of the aptamer-conjugated NP systems and their advantages and disadvantages in capturing CTCs.
Recently, a new class of nanoparticles have been utilized for the capture and detection of CTCs. Upconversion nanoparticles (UCNPs) carry the potential to increase the sensitivity of tracking cells in vivo to the single cell level. UCNPs have advantages over other fluorophores (eg, quantum dots) because of their ability to absorb light in the low-energy near-infrared and emit light at a shorter wavelength [127]. Another major advantage over other fluorescent particles is that UCNPs do not suffer from problems with background autofluorescence when they are used for imaging or integrated in a platform for detection of cancer cells [128].
Similar to the findings of Zhao et al., the Fan group succeeded in the detection and isolation of human acute leukemia cells from a cell mixture in buffer using up to 95 copies of aptamers attached to gold nanoparticles (AuNP) (DNA nanospheres). The aptamers that target leukemia cells were covalently bound to gold nanoparticles onto microfluidic devices. The AuNP-aptamers conjugate increased the capture efficiency of human acute leukemia cells from a cell mixture in buffer from 49% using aptamer alone to 92% using AuNP-aptamer [129]. As Table 3 also shows, Fang et al. were the first group to use UCNPs as nanoprobes to detect tumor cells [132]. Here, an aptamer against PTK7 was conjugated to UCNPs. The limit of the detection of CTCs was as low as 10 tumor cells spiked in 0.5 mL of whole blood. Notably, this nanoprobe could bind to the tumor cells in the presence of many nontumor cells, which reflect high selectivity [132].
Detection of CTCs Using Multiple-Target Aptamer-Based Prototypes
As mentioned previously, tumor cells could decrease the expression of some of their epithelial markers during the EMT process, or even lose them completely, causing significant heterogeneity of epithelial markers for example, EpCAM [12,13,134]. Therefore, a single CTC marker will be insufficient to identify the actual or entire CTC population in the blood. Use of a multivalent aptamer platform for capturing CTCs allows for the binding with multiple affinities to the target cells leading to the identification of the molecular profile and the phenotype of the CTCs more thoroughly than using single-target platforms. A study from Viraka Nellore et al. showed an aptamer-modified porous graphene oxide membranes for the capture and identification of multiple types of CTCs spiked in rabbit blood [135]. Three aptamers S6, A9, and YJ-1 for breast, prostate, and colon cancer cells, respectively, were attached to porous graphene oxide membranes. The three aptamers were capable of capturing SKBR3 breast cancer cells, LNCaP prostate cancer cells, and SW-948 colon cancer cells selectively and simultaneously when 100 cells were spiked in 1 mL of rabbit blood with a capture efficiency as high as 98% [135].
In another interesting example, as mentioned in Table 3 and illustrated in Fig. 4, three different aptamers (TD05, Sgc8, and Sgd5) against Ramos cells from a human lymphoma cell line and CCRF-CEM cells originating from human leukemia were immobilized on the surface of barcode nanoparticles (Fig. 4). This system, when integrated in a microfluidic, succeeds in the capture, detection, and release of multiple types of CTCs, and furthermore allows the culturing of CTCs for further studies [133].

Schematic of barcode particles used to capture different types of CTCs: aptamers were immobilized on the surface of three types of barcode particles (reflecting red, green, and blue) using a PAMAM dendrimer. The aptamers were TD05 that bind to Ramos cells, while Sgc8 and Sgd5 both bind to lymphoma cells with high specificity. Captured CTCs were released from aptamers using exonuclease I enzyme. Released CTCs could then be cultured and analyzed by microscopy. (Adapted from 116). Color images available online at www.liebertpub.com/nat
To be used in a clinical application, an assay for the detection of CTCs should be high-throughput (minimum number of steps) and highly sensitive (utilizing a minimum volume of the blood). In a study by Zeng et al., a one-step prototype was established for the detection of CTCs in which they end-modified an aptamer against CD30 protein with a fluorochrome-quencher system to form an aptamer reporter system [136]. The interaction of the aptamer and the CD30 protein on the cell surface allows specific uptake in tumor cells through endocytosis and, subsequently, into cell lysosomes. The lysosome-activated degradation of the system leads to the separation of the fluorochrome and quencher molecules, which restores the fluorescent signal within the tumor cell with low background noise [136]. The advantage of this method is the rapid detection of CTCs (30 min) and the high sensitivity due to the absence of background signal inside the tumor cells.
Antibody and Peptide-Based Platforms for the Detection and Isolation of CTCs
To advance aptamer-based CTC detection and isolation technologies, it may be valuable to examine innovative systems prepared from other molecular recognition elements for comparison. Antibodies, for example, have been used widely as a ligand for capturing CTCs. Many studies have employed several types of NPs conjugated to antibodies (mainly EpCAM) as a scaffold for the capture CTCs. Systems have been explored in which NP-antibody conjugates were used alone [137–139], or integrated into a micro device [140].
In an interesting study by Lee et al. [141], a hybrid nanoparticle system was established. The system consists of antibodies that bind to specific surface proteins on CTCs, a quantum dot used for fluorescence detection, and biotinylated DNA for binding CTC-capturing particles to a streptavidin-coated chip. As the DNA can be cleaved by restriction enzymes after the binding of antibodies to CTCs, the CTCs can be released (86.1% efficiency) for further analysis. Three different types of antibodies were used (EpCAM, EGFR, and HER2) to increase the selectivity for CTCs and increase the sensitivity to 92.4% [141].
In a proof-of-concept study by Sha et al. [142], surface enhanced Raman spectroscopy (SERS) was used to increase the detection sensitivity of CTCs from whole blood, using two conjugated antibodies (anti-EpCAM and anti Her2) to magnetic beads and SERS tags. The detection limit of this method was ∼50 tumor cells/mL of whole blood in less than 1 h [142]. The most important advantage of SERS is the sharp spectral signature that it gives, which eliminates the need for further separation techniques for the sample [142]. To improve the lower detection limit using the same system, another study by Wang et al. used a peptide against epidermal growth factor as the ligand (to increase specificity of binding to CTCs) conjugated to SERS nanoparticles. In this case, the lower limit of detection was improved to 5–50 tumor cells/mL of whole blood [143].
In an experiment to detect and isolate live CTCs in real time, Wang et al. used anti-EpCAM antibody conjugated to magnetic UCNPs as the probe, which was incorporated into a hybrid microfluidic device with two large symmetric chambers made by polydimethylsiloxane (PDMS) bonded onto the substrate containing arrays of silicon nanowires (SiNW) [144]. The capture efficiency of this platform was around 90%. CTCs could be released from the microfluidic chamber using an external magnet with 80% efficiency. Interestingly, CTCs were still viable after release from the chamber, which allowed further analysis of their culture, and is ideal for cancer prognosis [144].
Aptamers can rival antibodies and peptides in all of the systems mentioned above due to their practical advantages and superior properties in biomolecular recognition [55,56,58]. Aptamers are smaller and easier to modify (add functional groups, or covalently tether to different surfaces, including nanoparticles) than antibodies or peptides. Therefore, the sensitivity and stability of such systems will increase. Moreover, the release of CTCs from aptamers can be done easily by using a complementary DNA. This would have little or no impact of the viability of the cells, which could improve the analysis process afterward.
Conclusions and Future Prospects
CTCs play an important role in cancer metastasis. Therefore, their enumeration and detection may be used to understand the mechanism by which they initiate metastasis, thus leading to the development of more effective therapies, more accurate cancer diagnoses and prognoses, and improved cancer treatment. There is no ideal marker that can reliably and efficiently differentiate all CTCs from other blood cells. Therefore, new markers are still needed to capture and isolate these cells. Moreover, increasing the sensitivity and specificity by investigating new ligands is required. Aptamers have great potential for the future of CTC detection as they bind to their targets with high affinity and selectivity. Cell-SELEX produces aptamers that detect CTCs, and identify new biomarkers. Aptamers are preferable to use in CTC detection platforms as they are more stable than antibodies and they bind to their targets with high specificity and selectivity. Aptamers also can be used with a wider range of experimental conditions.
Aptamer detection platforms show great potential in the detection of CTCs. Some of them show improvement of the sensitivity when using multiple aptamers to capture CTCs, for example, micro-devices. For these platforms to be used in a clinical environment, they should provide high purity and higher throughput. Also, most of the platforms mentioned above have only been tested using spiked cells in a diluted buffer or blood sample, which does not reflect the true complexity of a real clinical sample. Some of the reviewed platforms have the ability to recover viable CTCs from a real blood sample with high efficiency and thus allow further analysis of the molecular profile and the phenotype of the captured CTCs at the single cell level. Although antibodies and peptides were used in some of these platforms as targeting ligands to capture CTCs, aptamers could increase the sensitivity, selectivity, and stability of these platforms, thus improving their chances of success in the clinic.
The data regarding the sensitivity of most of the devices reviewed are lacking; indeed most studies focused more on the specificity of their ligands over the sensitivity. Therefore, the sensitivity of these systems should be addressed with more real data and should be compared to the sensitivity achieved using other ligands, for example aptamers versus antibodies, in capturing CTCs.
Footnotes
Acknowledgment
M.C.D. and W.G.W. acknowledge the Natural Sciences and Engineering Research Council of Canada (NSERC) for funding.
Author Disclosure Statement
No competing financial interests exist.