
Abstract
Changes in microRNA (miRNA) expression levels that play important roles in regulation lead to many pathological events such as cancer. The miR-200 family is an important target in cancer therapy. The aim of this study is to equilibrate endogenous levels between cancer and noncancerous cells to prevent serious side effects of miR-200c- and miR-141-like metastatic colonization. For the first time, the characterization of miR-200c and miR-141 cluster containing chitosan nanoplexes was shown, and the optimization of miRNA expression levels by conducting dose studies in breast cancer cell lines was made. The mean diameter of chitosan/miR-141 and chitosan/miR-200c nanoplexes ranged from 296 to 355 nm and from 294 to 380 nm depending on the N/P ratio, respectively. The surface charge of nanoplexes was positive with zeta potential of +12 to +26 mV. While naked miRNA was degraded after 0 min in a 10% serum-containing medium, chitosan/miRNA nanoplexes were protected for 72 h. During the in vitro cellular uptake study, nanoplexes were observed to be accumulating in the cytoplasm or nucleus. After using different doses for miR-200c, the determined doses are 750, 100, and 750 ng in the MCF-7, MDA-MB-231, and MDA-MB-435 cell lines, respectively. Doses were determined as 100 ng for MDA-MB-231 and 150 ng for MDA-MB-435 to reach endogenous miR-141 levels of MCF-10A. Our results suggest that chitosan nanoplexes for miR-200c and miR-141 are an efficient delivery system in terms of formulation and transfection. As a conclusion, dose studies are important to provide effective treatment with miRNAs.
Introduction
B
Despite the important role of miRNAs in cancer therapy, the main challenges to miRNA therapeutics in in vivo applications remain the same as the other nucleic acid-based therapeutics. These include stability, cellular uptake, off-target effects, immunogenicity, and toxicity. As a result, appropriate delivery systems for miRNA should be developed. Among the nonviral gene delivery systems currently used, chitosan is a biodegradable, biocompatible, nontoxic, and low-immunogenic biopolymer. As a cationic polymer, chitosan is a particularly promising nonviral vector for nucleic acid-based therapeutics [10–13].
To date, very few articles related to a chitosan-based system for miRNA are present in the literature. Tekie et al. prepared vector-based miR-145 containing chitosan polyplex nanoparticles for transfection to MCF-7 cells [14]. They indicated that particle size, zeta potential, N/P ratio, time of incubation, and molecular weight of the polymer affect the gene transfection. Selecting formulations showed high transfection efficacy for miR-145 as an anticancer biological macromolecule [14]. Zhu et al. developed a probe using nanoparticles of miR-155 MB self-assembled with chitosan (CS) (CS-miR-155 MB) to image the expression of miR-155 in cancer cells [15]. Gaur et al. reported that chitosan nanoparticle-mediated delivery of miR-34a, a tumor suppressive miRNA that downregulates multiple gene products involved in PCa (prostate cancer) progression, and metastasis, inhibited prostate tumor growth and preserved bone integrity in a xenograft model representative of established PCa bone metastasis [16]. Santos-Carballal et al. investigated chitosan–hsa-miRNA-145 nanocomplexes and the biological responses of MCF-7 breast cancer cells cultured in vitro [17]. Self-assembled CS–miRNA nanocomplexes were produced with a range of (+/−) charges. Nanocomplexes with degrees of acetylation of 12% and 29% were biologically active, showing successful downregulation of target mRNA expression in MCF-7 cells [17]. Doxorubicin and miR-34a-encapsulated hyaluronic acid-chitosan nanoparticles were prepared and then investigated the therapeutic effects in vitro and in vivo in triple-negative breast cancer models. Codelivery of doxorubicin and miR-34a achieved synergistic effects on tumor suppression [18].
When miR-200s are given in high doses, it has been reported that they induce metastasis in breast cancer [19–21]. MiRNAs should be given in appropriate doses to avoid side and off-target effects. Therefore dose studies are needed; we tried to provide endogenous levels of miRNA by given miRNA 200c/141 and chitosan complex. To date, there have been no reports about the dose studies of formulations to provide effective treatment with miRNAs. The aims of this study are to characterize miR-200c and miR-141 containing chitosan nanoplexes and optimize miRNA expression levels by dose studies in various breast cancer cell lines, including MCF-7, MDA-MB-231, and MDA-MB-435, and to adjust levels of miR-200c and miR-141 levels according to the MCF-10A normal breast cell line.
Materials and Methods
Materials
Hsa-miR-200c, hsa-miR-141, cel-miR-67 as negative control, and Dy547-labelled cel-miR-67 for transfection studies were obtained from Dharmacon (miRIDIAN™, United Kingdom). Chitosan (75 kDa; 75%–85% deacetylation) was purchased from Sigma (St. Louis, MO). All the substances used in this study are of molecular grade.
Preparation and control of chitosan/miRNA nanoplexes
Chitosan stock solution (1% w/v) was prepared by dissolving chitosan in sterile 40 mM acetate buffer (pH 5.4) and filtering the solution under sterile conditions. Lyophilized miRNAs were dissolved in the siRNA buffer solution according to manufacturer's protocol. Chitosan/miRNA nanoplexes were prepared by the addition of miRNA to chitosan solution by pipetting. The mixed solution was stirred on a vortex mixer and incubated for 30 min at room temperature for forming nanoplexes completely. Chitosan/miRNA nanoplexes were prepared at different charge (N/P) ratios (from 0.1 to 8) using constant miRNA concentration. N/P ratios were calculated according to the previous article of Fernandez et al. [22].
The formation of nanoplexes was checked by agarose gel electrophoresis using 2% of gel at 80 mA. MiRNA was visualized under ultraviolet light as a result of reaction of miRNA with RedSafe™ (Intron Biotech, South Korea). Nanoplexes were prepared in triplicate (n = 3).
Stability of miRNA nanoplexes against serum
Stability of chitosan/miRNA nanoplexes against serum was studied by incubating the samples in 10% fetal bovine serum at 37°C. The integrity of miRNA was examined using the agarose gel retardation assay. Serum stability of free miRNA was studied at different intervals (0, 30, 60, 120, and 240 min; 24, 48, and 72 h) and analyzed with agarose gel electrophoresis [23].
Particle size and zeta potential of chitosan/miRNA nanoplexes
Particle size (diameter of particles) and zeta potential values of chitosan/miR-200c and chitosan/miR-141 nanoplexes were measured by using ZetaSizer NanoZS (Malvern Instruments, United Kingdom) after being dissolved in phosphate-buffered saline (PBS) pH 7.4. Each measurement was done in triplicate (n = 3).
Imaging of nanoplexes
The morphological examination of nanoplexes was made by transmission electron microscopy (TEM, JSM 5200; Jeol, Tokyo, Japan). Specimens for TEM measurements were prepared by spreading of one drop of dilute solution of nanoplexes on top of carbon-coated copper grids and then grids left to dry in air. The size and shape of nanoplexes were also checked.
Decomplexation control studies of nanoplexes
Decomplexation study of miRNA from nanoplexes was done according to previous article of Laroui et al. Nanoplexes were mixed with 0.1 M NaOH at a ratio of 1:1 and controlled with agarose gel electrophoresis within 1 min [24].
In vitro transfection studies
The human breast cell line MCF-10A and breast cancer cell lines MCF-7, MDA-MB-231, and MDA-MB-435 (ATCC, Rockville, MD) were used and cultured in Dulbecco's modified Eagle's medium (DMEM) containing 10% heat-inactivated fetal bovine serum, 2 mM glutamine, penicillin, and streptomycin antibiotic solution (Biological Industries), which was maintained in an incubator at 37°C with 5% CO2 in a humidified atmosphere (Sanyo CO2 Incubator, Japan). The MEGM Bullet kit (Lonza) was also used for MCF-10A cell line. The culture medium was replaced each 3 days and cells at logarithmic growth phase were used for following experiments. Cells were plated in a six-well plate at a cell density of 5 × 105 cells/well and incubated overnight. Transfection were performed on cells that were ∼70% confluent. Different doses of chitosan:miRNA complexes were used in the transfection study for dose studies. Nanoplexes were added to cells and incubated for 24–48 h for transfection.
In vitro cellular uptake studies
Cells were seeded on coverslips at densities of 5 × 105 cells/well. After 48 h from transfection of chitosan nanoplexes carrying Dy547-labeled cel-miR-67 mimic miRNAs (N/P = 4), cells were washed with PBS and fixed with 4% paraformaldehyde in PBS. Coverslips were mounted on glass slides and cells. Cellular uptake was evaluated by using a fluorescence microscope.
Determination of miR-200c and miR-141 levels of normal and cancerous breast cell lines by qRT-PCR
Endogenous miR-200c and miR-141 levels were determined to make dose optimization of miRNA levels in normal and cancerous breast cell lines. After 48 h from passage to six-well plate, miRNA isolation was performed by the mirVana miRNA Isolation kit, with phenol (Ambion) according to manufacturer's protocol. After total RNA isolation from cells, the controls of RNA were made spectrophotometrically and electrophoretically as mentioned above and the isolated RNA was stored at −80°C until used. For miRNA expression analysis, 10 ng of total RNA was applied for reverse transcription to synthesize cDNA of miRNA using the TaqMan® miRNA Reverse Transcription Kit (Applied Biosystems). Quantitative real-time polymerase chain reaction (qRT-PCR) analysis was performed using TaqMan Universal Master Mix (Applied Biosystems) and TaqMan microRNA assays (Applied Biosystems) for hsa-miR-200c and hsa-miR-141 according to manufacturer's instructions. Each sample was analyzed in triplicate. Relative expressions of miRNAs were calculated using the comparative CT method. All data were controlled for quantity of RNA input by performing measurements on an endogenous reference gene, RNU6B. In addition, results obtained on RNA from cancerous samples were normalized to results obtained on RNA from the control, MCF-10A. Results were expressed as relative quantitation (RQ).
Dose studies of miRNA
After determination of endogenous miRNA levels in normal and cancerous breast cell lines, downregulated miRNA levels in breast cancer cell lines have regulated miRNA levels to reach that of normal breast cell line by using mimic miRNAs. For this purpose, chitosan/miR-200c and chitosan/miR-141 nanoplexes (N/P = 4) in different doses (2.5, 1, 0.75, 0.5, 0.2, 0.15, and 0.1 μg) were transfected to breast cancer cell lines in six-well plates. Doses were determined to optimize miRNA levels of breast cancer cell lines according to MCF-10A, normal breast cell line. Prepared nanoplexes in the determined doses were transfected to cells.
Cell viability study
Cell viability or cytotoxicity was performed by using the WST-1 (4-[3-(4-lodophenyl)-2-(4-nitrophenyl)-2H-5-ltetrazolio]-1,3-benzene disulfonate) assay (Roche, Germany). The test is designed to be used for spectrophotometric quantification of cell proliferation, viability, and cytotoxicity. Cell viability assay was done by the WST-1 assay (Roche, Germany) according to the manufacturer's protocol. Cells were seeded at a concentration of 4 × 103 cells/well in 100 μL culture medium (1 × DMEM) into 96-well plates. Cells were incubated overnight at 37°C and 5% CO2. The medium was removed and prepared formulations were added with 50 μL serum-free medium for each cell. After 4 h of incubation, 50 μL medium was added with 10% serum to each well. Cells were incubated for 48 h at 37°C and 5% CO2. Ten microliters per well of Cell Proliferation Reagent WST-1 was added and cells were incubated for 4 h at 37°C and 5% CO2. Then, it was shaken thoroughly for 1 min on a shaker. Absorbance of the samples was measured against a background control as blank at 440 nm wavelength using a spectrometer.
Statistical analysis
Experiments were performed in triplicate. Data were presented as the mean ± standard deviation and were analyzed with the Paired t-test using SPSS 20.0 software (Chicago, IL). P < 0.05 was considered to indicate a statistically significant result.
Results
Preparation and in vitro characterization of nanoplexes
Nanoplexes were prepared by mixing chitosan with miRNAs at a certain N/P ratio (0.1–8). The complete nanoplexes with miR-200c and miR-141 were formed at an N/P = 4 ratio. As seen in Fig. 1, N/P ratios below this ratio partially complexed miRNA with chitosan due to the weak electrostatic interaction between miRNA and chitosan. The 2% agarose gel electrophoresis of chitosan/miRNA nanoplexes at various N/P ratios is shown in Fig. 1. Compared to the control, naked miRNA chitosan binding caused retarded mobility of miRNA. Nanoplexes forming full complexation were retained within the well.
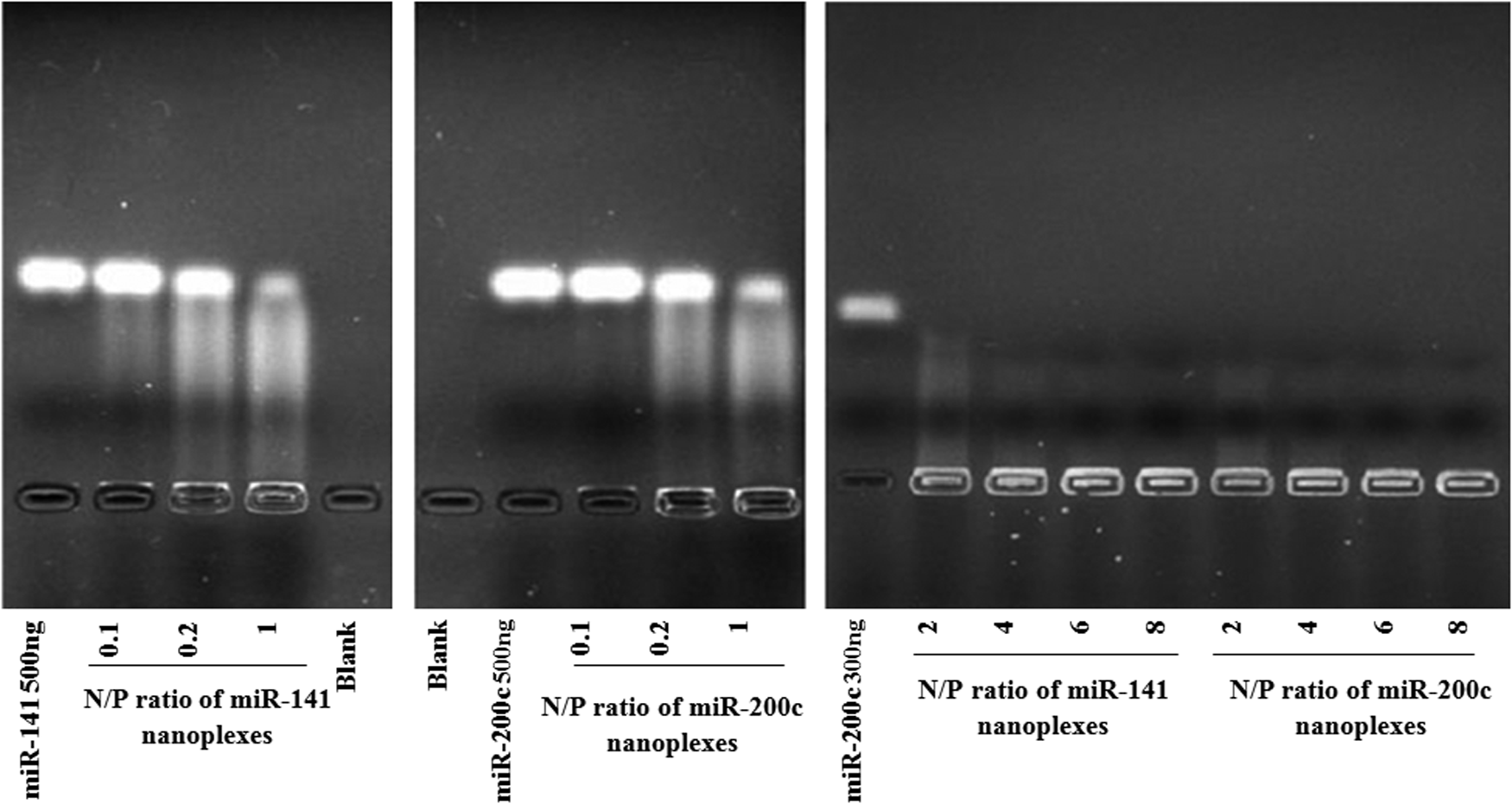
Agarose gel electrophoresis of chitosan/miRNA nanoplexes in the different N/P ratios. miRNA, microRNA.
Nanoplex shape and size were controlled with TEM (Fig. 2). TEM revealed that chitosan nanoplexes had a relatively spherical shape. Particle size and zeta potential are very important variables for cellular uptake of nanoplexes and these parameters may also affect particle stability. The particle size of chitosan/miR-141 nanoplexes changed between 296.1 ± 1.0 and 355.8 ± 0.2 nm, and the sizes of chitosan/miR-200c were between 294.8 ± 5.0 and 380.4 ± 4 nm depending on (+/−) the charge ratio (Table 1).

Transmission electron microscopy photographs of chitosan nanoplexes with miR-200c and miR-141, respectively
SD, standard deviation.
The positive surface charge of chitosan/miRNA nanoplexes increased with an increasing ratio of chitosan at a constant miRNA ratio. The mean zeta potential values of nanoplexes varied from +12.4 ± 0.6 to +25.8 ± 0.8 mV. Zeta potential values increased with an increase in the chitosan/miRNA ratio (P < 0.05).
The serum stability of chitosan/miRNA nanoplexes was studied with 10% serum-containing media for 72 h (Fig. 3). The chitosan/miR-141 and chitosan/miR-200c nanoplexes were protected from degradation miRNA until 72 h, while rapid degradation of naked miR-141 was observed as early as time point of 30 min and naked miR-200c was fully degraded after 24 h. Chitosan/miRNA nanoplexes significantly protected miRNAs against the serum proteins.
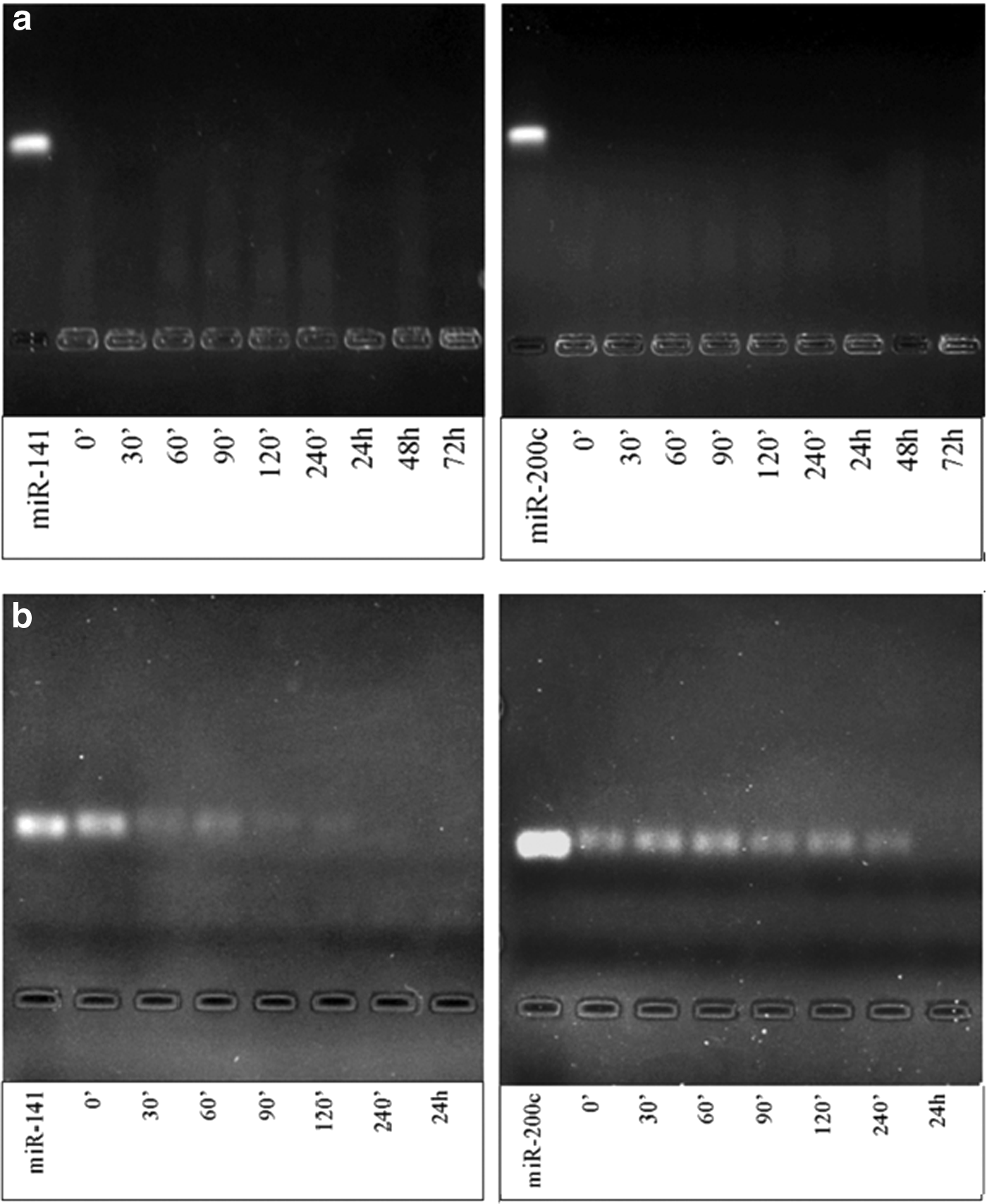
Serum stabilities of chitosan/miRNA (N/P = 4) nanoplexes
The effect of nanoplex formation on miRNA structure was investigated by the decomplexation study, and it was seen that miRNA has been decomplexed from nanoplexes without damage (Fig. 4).
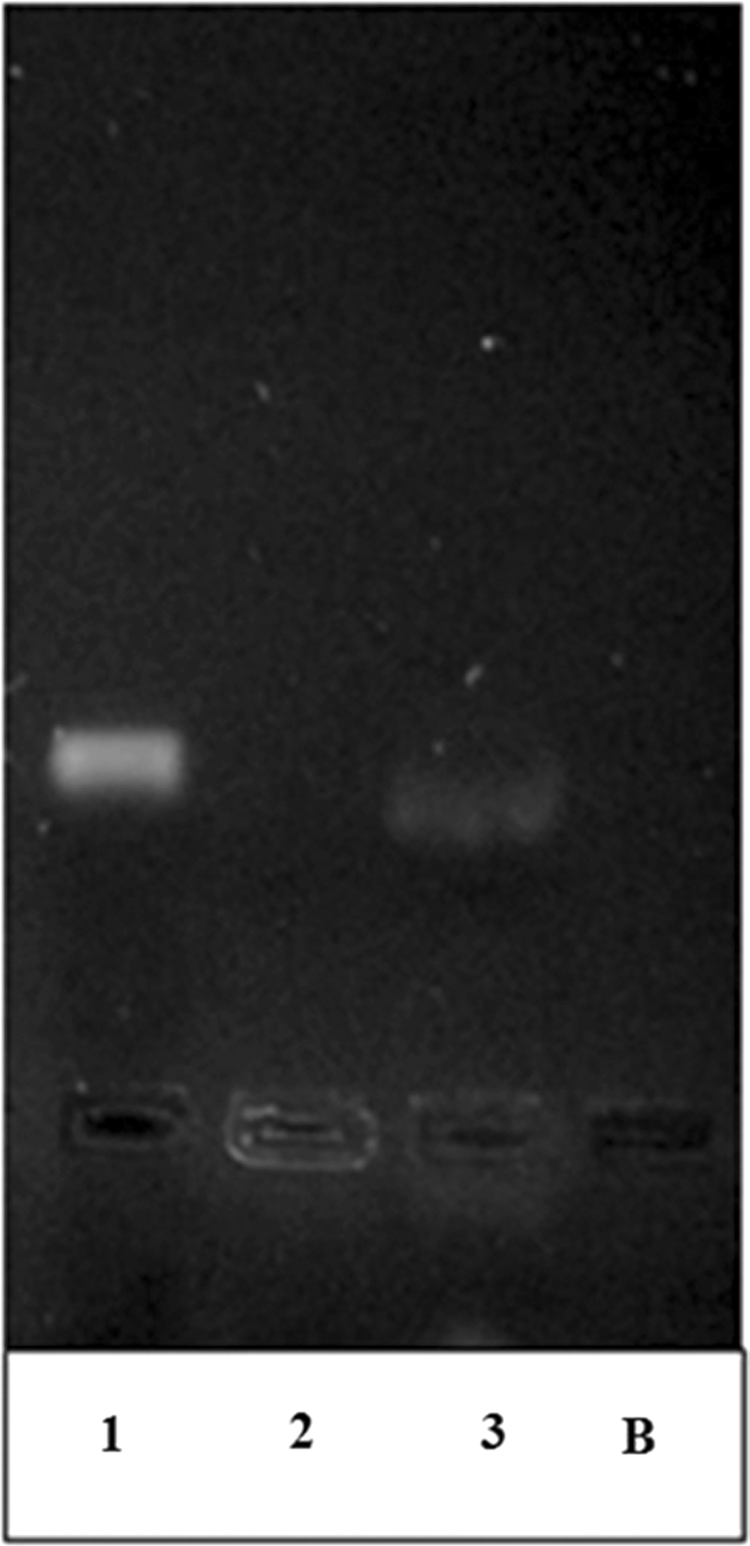
Agarose gel electrophoresis of free miRNA, chitosan/miRNA, and decomplexed chitosan/miRNA nanoplexes (N/P = 4). 1, Free miRNA; 2, complexed miRNA; 3, decomplexed miRNA; B, blank.
In vitro cellular uptake studies
For control of transfection efficiency of chitosan/miR-141 and chitosan/miR-200c nanoplexes, Dy547-labeled miRNA mimic was used and complexed with chitosan at an N/P ratio = 4. Cellular uptake and localization were observed using a fluorescence microscope. As seen in Fig. 5, transfection efficiencies were about 100% in the MCF-7, MDA-MB-231, and MDA-MB-435 cell lines.

Cellular localization of chitosan/Dy547-miR-67 mimic (N/P = 4) nanoplexes in MCF-7, MDA-MB-231, and MDA-MB-435 cell lines at 4th
After transfection, cellular uptake was observed after 4, 24, and 48 h. As seen in Fig. 5, most complexes were not internalized by the cells; they were localized in the medium of all three cell lines. However, after 24 h of incubation, some complexes were outside the cells, while most of them were localized in the cytoplasm (Fig. 5d–f). However, after 48 h of transfection, all the nanoplexes were internalized into the breast cancer cells (Fig. 5g–i).
In vitro endogenous miR-141 and miR-200c expression profile studies
For the determination of endogenous miRNA expression levels, healthy normal and cancerous breast cells were used. Endogenous miR-141 level was not changed in the MCF-7 cell, but it is seen that miRNAs were significantly downregulated in MDA-MB-231 and MDA-MB-435 cell lines (Fig. 6). Markedly downregulated miR-200c level was determined in all the breast cancer lines when compared to the normal breast cell line (Fig. 6).

Endogenous miR-200c and miR-141 expression profiles in healthy normal and cancerous breast cells (n = 3).
When we compared miR-200c and miR-141 expression levels in breast cancer cell lines, we observed different expression levels in different cell lines. Although the miR-141 level has not downregulated in the MCF-7 cell, the miR-200c level was downregulated (60% ± 4.2%). miR-141 and miR-200c levels are in downregulated levels in the MDA-MB-231 cell, respectively, by 70% ± 2.9% and 99% ± 0.2%. When miRNA levels were compared in MDA-MB-435 and MCF-10A cells, downregulation levels of miR-141 and miR-200c were 90% ± 0.2% and 99% ± 0.1%, respectively, (P < 0.05).
In vitro dose studies with chitosan/miRNA nanoplexes
The purpose of a dose study is to eliminate the expression differences between normal and tumor groups. According to endogenous expression levels in normal breast cells, the rearrangement of miR-200c and miR-141 levels in breast cancer cells is important in terms of breast cancer therapy. Therefore, transfection and dose studies were made using qRT-PCR for reaching endogenous miRNA expression levels of normal breast cell line to the downregulated miRNA levels in breast cancer cell lines.
In this study, miRNA containing chitosan nanoplexes (N/P = 4) at different doses of miRNA (5, 2.5, 1, 0.75, 0.5, 0.2, and 0.1 μg) were used for dose studies (Figs. 7–11). According to the result of the dose studies, the optimal dose for miR-200c containing chitosan nanoplexes was determined as 750 ng in the MCF-7 cell line (Fig. 7). In the MDA-MB-231 cell line, 100 ng miR-141 and miR-200c containing chitosan/miRNA nanoplexes were required for an appropriate dose (Figs. 8 and 9). In the MDA-MB-435 cell line, 750 ng miR-200c and 150 ng miR-141 containing chitosan nanoplexes were found to be necessary for the upregulation of miRNAs levels to endogenous levels (Figs. 10 and 11).
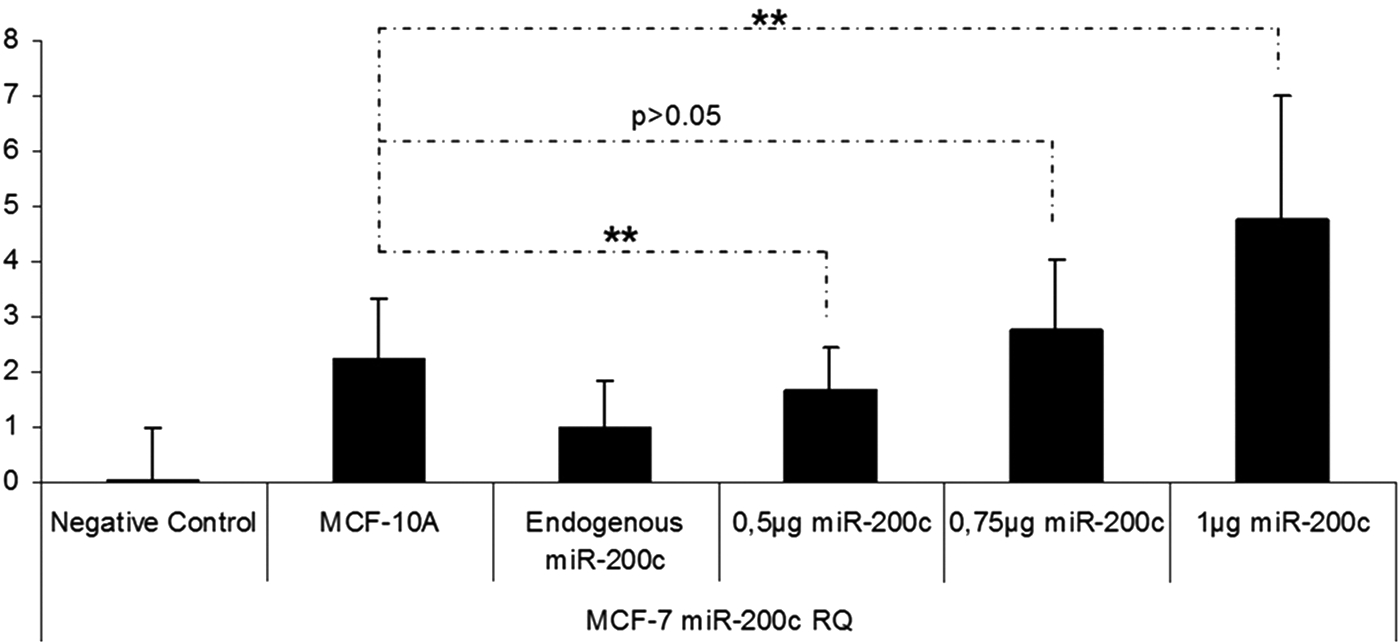
miR-200c expression profiling after transfection of miR-200c containing chitosan nanoplexes in the different doses in MCF-7 cells. **P < 0.01.
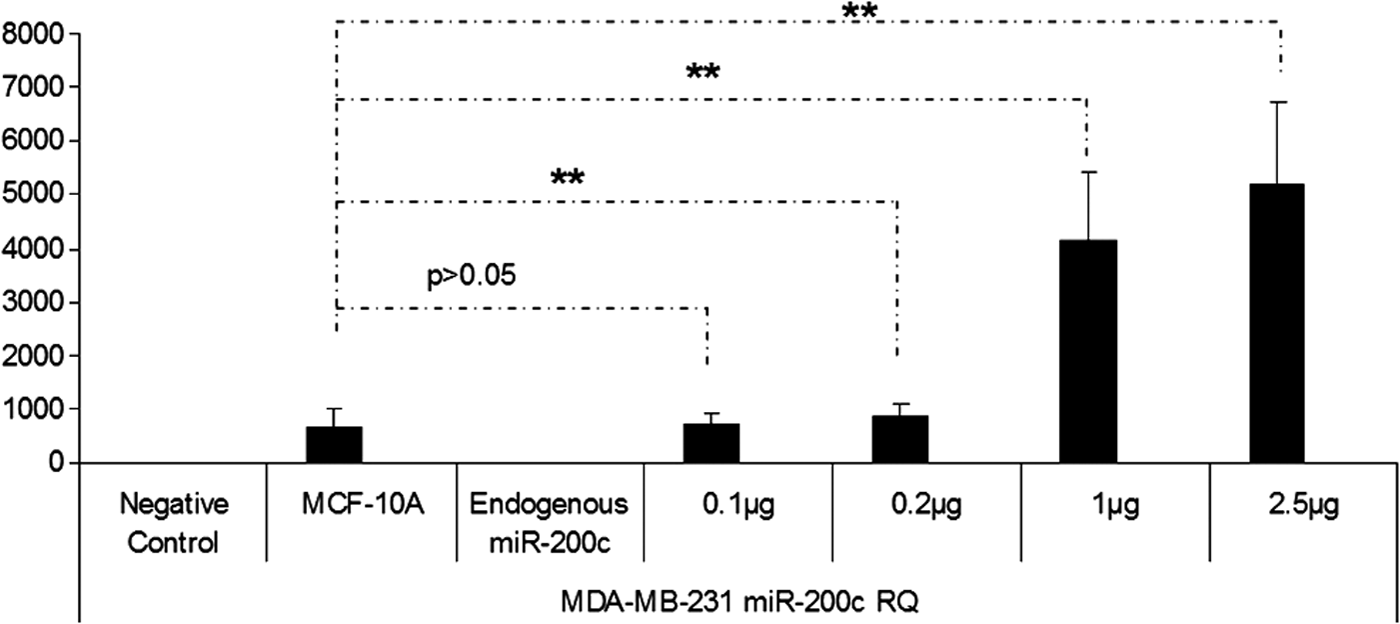
miR-200c expression profiling after transfection of miR-200c containing chitosan nanoplexes in the different doses in MDA-MB-231 cells. **P < 0.01.
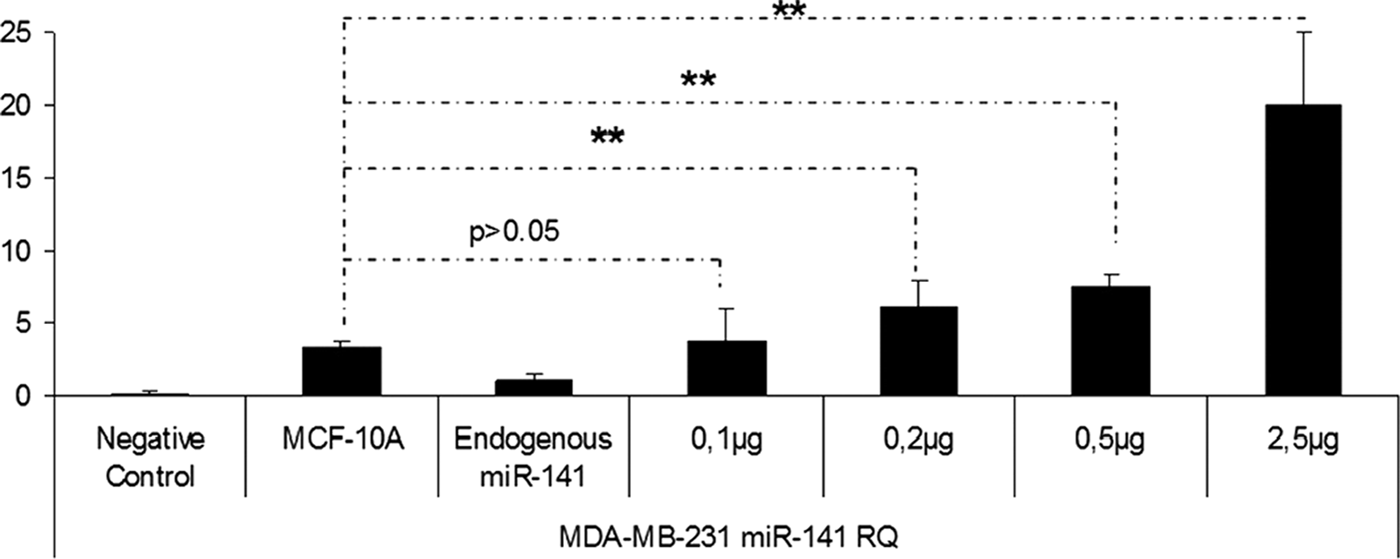
miR-141 expression profiling after transfection of miR-141 containing chitosan nanoplexes in the different doses in MDA-MB-231 cells. **P < 0.01.

miR-200c expression profiling after transfection of miR-200c containing chitosan nanoplexes in the different doses in MDA-MB-435 cells. *P < 0.05, **P < 0.01.
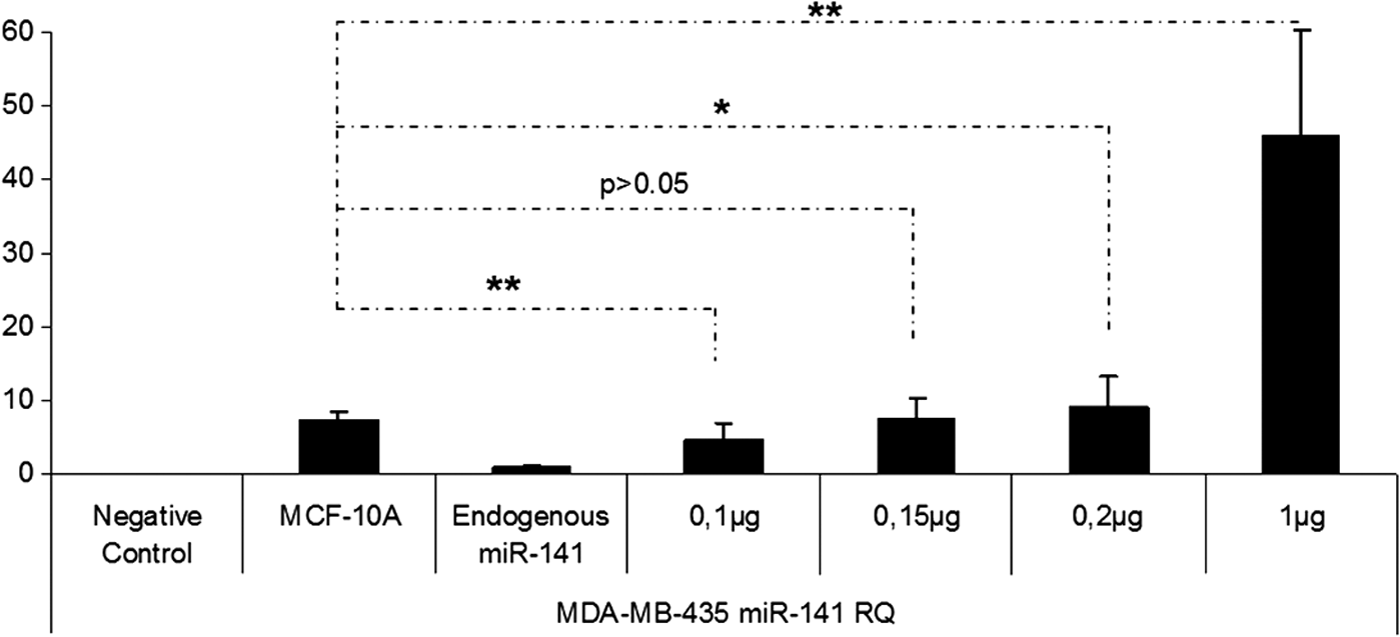
miR-141 expression profiling after transfection of miR-141 containing chitosan nanoplexes in the different doses in MDA-MB-435 cells. *P < 0.05, **P < 0.01.
Cell viability study
Cell viability or cytotoxicity studies were conducted using Cell Proliferation Reagent WST-1 in mouse fibroblast cell lines (L-929) and breast cancer cell lines (Fig. 12). Our cell viability findings show that there is no difference between the control and chitosan/miR-200c nanoplex (N/P = 4) groups in the L-929 cell line. We observed that cell proliferation did not change after transfection of chitosan/miR-200c and chitosan/miR-141 nanoplexes compared to the control in MCF-7, MDA-MB-231, and MDA-MB-435 cell lines (P > 0.05).
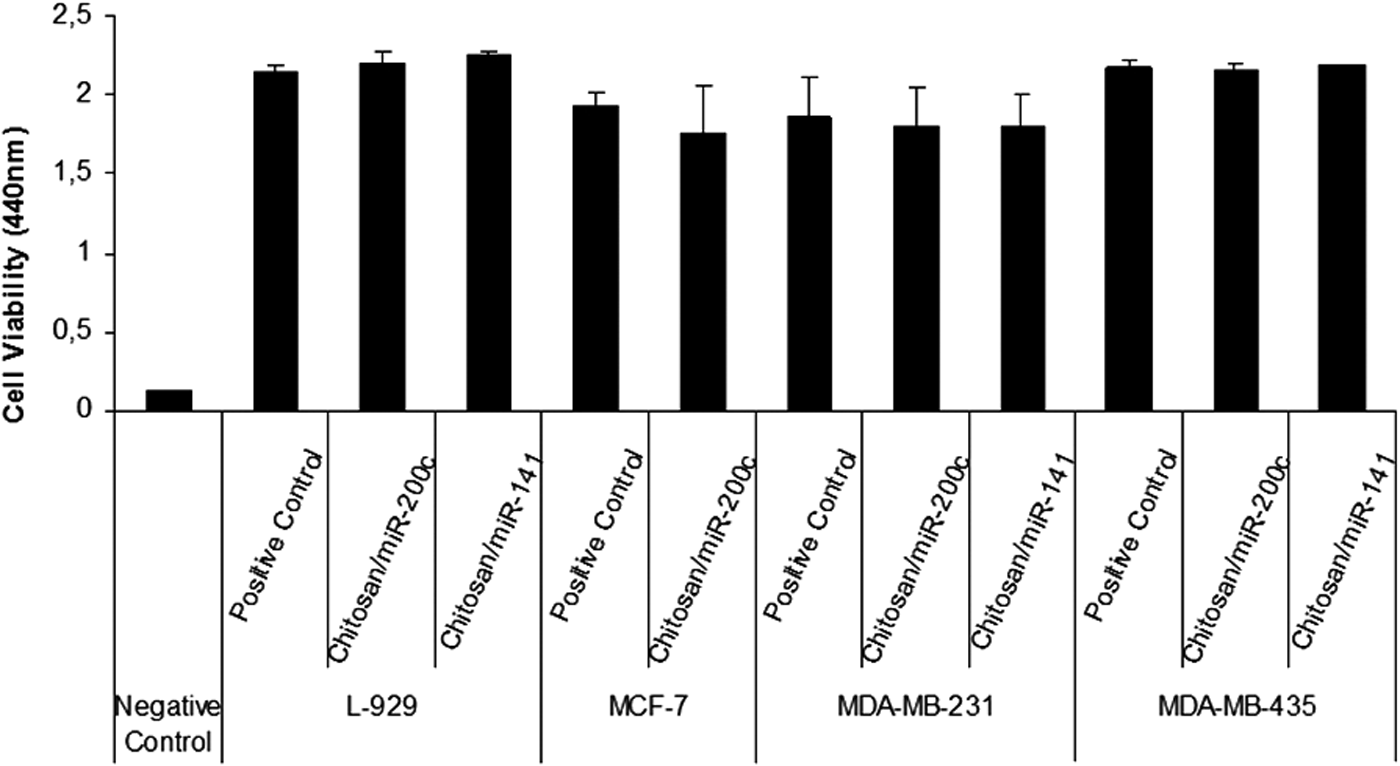
Cell viability studies of chitosan/miRNA nanoplexes in the L-929 and breast cancer cell lines (n = 3).
Discussion
Recently, miRNA-based cancer therapy has come to represent new and disease-specific approaches to cancer gene therapy. miRNAs can function as tumor suppressors or oncogenes during tumor initiation and progression. The expression of tumor-suppressive miRNAs lowered in tumor cells compared to normal cells. miR-200s have been shown to be remarkably downregulated in a variety of cancers [25,26]. Lower level expressions of miR-200c and miR-141 have been reported in breast cancer [27–29] as we determined in this study (Fig. 6).
The most important therapeutic challenge for miRNAs is failing to reach the target cell and tissue, cellular uptake, because of a negative charged backbone, leading to electrostatic repulsion [30]. The other limitations are inappropriate distribution, disruption, poor in vivo stability and saturation of endogenous RNAi pathway, and unwanted side effects [8]. The other important point is the metastasis-enhancing effect of miR-200s in breast cancer. miR-200s enhance the colonization of cancer cells and metastasis [8,19,20]. Besides that, dose of miR-200s are very important in cancer treatment. Safe, stable, effective, and specific delivery systems are required for the translation of miRNA-based therapeutics into successful clinical applications. In this study, we used chitosan biopolymer as the carrier for the delivery of miRNA to the breast cancer cell lines and investigated appropriate miR-200c and miR-141 levels in breast cancer cell lines by conducting dose studies with mimic miRNAs to reach endogenous levels in noncancerous breast cells.
Nanoplex formulations were prepared by the simple complexation method as described in our previous article [23]. The formation of nanoplexes by this method is dependent on the chitosan/miRNA N/P ratio [14]. In our study, miR-200c and miR-141 easily formed full complexes with low Mw chitosan at an N/P ratio = 4. The different ratios of chitosan to miRNA were prepared (0.1–8) and complexation was controlled. While partial complexation was formed at an N/P ratio = 2, full complexation was obtained after an N/P ratio = 4 (Fig. 1). siRNA and miRNA required at least an N/P ratio above 2 to initiate nanoplex formation. Since siRNA and miRNA are small molecules, they may need much more chitosan charge density to initiate nanoplex formation [31]. The short length of siRNA (21–25 mer) and its linearity are like properties with miRNA structure [32].
In the study of Tekie et al., they used chitosan having low molecular weight and low deacetylation degree (1.3–40 kDa and <67%) and evaluated chitosan/plasmid miR-145 polyplexes [14]. They suggested that it is essential to determine the best formulation for delivery of miRNA plasmid. For the optimal design of polyplexes and the formation of chitosan/plasmid miR-145 polyplexes, miRNA plasmid vector concentration, stirring time, and incubation time are important parameters [14].
Nanoplexes were analyzed in terms of particle morphology, size, and surface topology. From the TEM images, the chitosan/miRNA nanoplexes showed different topological structures, including spherical and toroidal morphologies. The sizes determined by TEM analysis correlated with those determined by the particle sizer (Fig. 2). The mean diameter of chitosan/miR-141 nanoplexes ranged from 296 to 355 nm, while chitosan/miR-200c nanoplexes changed from 294 to 380 nm depending on the N/P ratio (Table 1). The obtained particle size ranges are suitable for efficient endocytosis by cells. The size of nanoplexes increased with an increase in N/P ratios. In particular, the size of nanoplexes significantly increased above the N/P ratio of 8. These data are in accordance with earlier reports [14]. The surface charge of nanoplexes is positive and zeta potential changed from +12 to +26 mV depending on the chitosan miRNA ratio (Table 1). When nanoplexes could not be formed completely, the zeta potential was negative, but with increasing N/P ratios, the zeta potential increased to a positive charge. The reason for this situation was the increase in the number of positive charges of chitosan that interact with negatively charged miRNA as the amount of miRNA was constant. Thus, the positive charge of nanoplexes promotes electrostatic interaction with the negative charge of the cell membrane.
The effect of the serum nuclease on degradation of nucleic acid therapeutics is a potential obstacle to functional delivery to the target cells [33]. miRNAs must be stable against the degradation effect of nucleases. As seen in Fig. 3, chitosan/miRNA nanoplexes protected miRNA until 72 h against serum attack. Chitosan protected the condensed siRNA and miRNA from enzymatic digestion and serum components [31,34]. After the internalization of nanoplexes, it is also important that high complex stability does not prohibit intracellular decomplexation of RNA [35]. As seen in Fig. 4, miRNAs were completely released from the chitosan nanoplexes with decomplexation. The enhanced stability of nucleic acid therapeutics with chitosan nanoplexes can be important for increased in vitro transfection efficiency [36].
An in vitro cellular uptake study with chitosan/Dy547-labeled mimic miRNA nanoplexes (20/1) was prepared for the control of transfection and cellular internalization of the nanoplexes as shown in Fig. 5. Nanoplexes were taken up into the cell after transfection. Similar to our research, Jayakumar et al. studied the transfection mechanism of chitosan/nucleic acid nanocomplexes and indicated that chitosan forms self-aggregation with nucleic acids [33]. Then, they are taken up into cells by endocytosis and possibly released from endosomes because of swelling endolysosomes, causing the endosome to rupture. Finally, these nanocomplexes were observed to be accumulating in the cytoplasm or nucleus (Fig. 5).
The second goal of this study was to determine the suitable dose for miR-200c and miR-141 therapy. It is known that miRNA expression levels of the cell are important, since high miRNA expression may have a negative effect on cellular viability and may be cytotoxic [37]. Recent in vivo studies have shown that the toxic effect of small RNAs appears to be related to saturation of the endogenous micro RNA pathway [38,39]. Previous studies suggested that siRNAs and shRNAs may enter the miRNA pathway either before or after microprocessor complex formation, so the similar pathways are used by miRNAs and shRNAs, and toxicity can be explained by the saturation of endogenous RNAi mechanism [38]. In another article, Grimm et al. reported less clear correlation between toxicity and premature miRNAs [39]. Therefore dose of miRNA may be important in the miRNA-based treatment.
On the other hand, when we design miRNA approaches, miRNA must be used at the lowest concentration possible as to interfere with tumor growth because of potential toxicity due to off-target effects [37,38]. In our study, before the in vitro transfection and efficiency studies of chitosan/miRNA nanoplexes, endogenous miR-141 and miR-200c levels were determined in MCF-7, MDA-MB-231, and MDA-MB-435 breast cancer cell lines compared to the MCF-10A noncancerous breast cell line. While the miR-141 level did not change in the MCF-7 cell line (2%) compared to the MCF-10A cell line, the miR-141 level downregulated in MDA-MB-231 (70%) and MDA-MB-435 (90%) (Fig. 6). As seen in Fig. 6, the miR-200c level decreased to about 60%, 90%, and 99%, in MCF-7, MDA-MB-231, and MDA-MB-435 cells, respectively. These data showed similarity to previous articles related to downregulation of miR-200c and miR-141 in cancer cells [18,40].
Recent studies reported the role in inhibiting cell malignant transformation and preventing tumor initiation of miR-200s that act as tumor suppressors [7]. However, contraindicated results are present in the literature about the dysregulation of miR-200 family in breast tumors [9,19–21]. Dykxhoorn et al. showed that upregulation of the miR-200 family has been associated with enhanced tumorigenesis and metastasis [19]. In accordance with above-mentioned facts, increased miR-200s expression levels may benefit in cancer therapy, but upregulation of miRNA levels may reveal unwanted effects. For this purpose, dose studies were conducted in this study.
After expression profiling studies of miR-200c and miR-141 in normal breast and cancer cells, because of upregulation or high expression levels of miRNAs in cells, miRNAs can be toxic [37] and promote metastasis in breast cancer [20,21]; dose studies have been conducted to reach miRNA levels in the noncancerous MCF-10A cells. After comparing different doses for miR-200c, the determined doses are 750, 100, and 750 ng in the MCF-7 (Fig. 7), MDA-MB-231 (Fig. 8), and MDA-MB-435 (Fig. 10) cell lines, respectively. For miR-141, doses were determined as 100 ng for MDA-MB-231 (Fig. 9) and 150 ng for MDA-MB-435 (Fig. 11). The miR-141 level is not changed or downregulated in the MCF-7 cell, therefore a dose study was not made in the MCF-7 cell line for miR-141.
In the study of Korpal et al., they showed that miR-200s promote metastatic colonization through direct targeting of Sec23a, which mediates secretion of metastatic suppressive proteins, including Igfbp4 and Tinagl1 [20]. After the dose studies, when we controlled metastatic faces of miR-200c and miR-141 on Sec23a with determined levels of miRNAs, we showed that the Igfbp-4 levels of the cells did not change, or slightly changed dependent on the cell types and the level of Tinagl1 increased in all the cells after being given chitosan/miR-200c nanoplexes [41] and chitosan/miR-141 nanoplexes (data not shown), when compared to the control group in another research.
We also assessed the effect of complexation on cell viability of chitosan/miRNA nanoplexes. As shown in Fig. 12, the viability of the mouse fibroblast cell line and breast cancer cell lines were not affected by miR-200c and miR-141 containing chitosan nanoplexes. These data are in accordance with other studies [17].
In conclusion, our study showed that dose studies are important in providing an effective treatment with miRNAs because of serious two-sided effects of high dose of miRNAs. This article will provide a new point of view with regard to rearranging the tumor suppressor as well as the oncogenic behavior of the same miRNAs. In addition, the delivery of miRNAs to breast cancer cells still remains a challenge. The findings of this study highlight the pivotal role of chitosan as a safe and efficient delivery system for miR-200c and miR-141 in terms of formulation and transfection.
Footnotes
Acknowledgments
This study was supported by the Marmara University Scientific Research Projects Association (SAG-C-YLP-120514-0137). We would like to thank to Prof. Dr. Oguz Ozturk for kindly providing the MCF-10A cell line.
Author Disclosure Statement
No competing financial interests exist.