
Abstract
Single-stranded (ss) 2′-fluoro (2′-F)-modified oligonucleotides (ONs) with a full phosphorothioate (PS) backbone have been reported to be cytotoxic and cause DNA double-strand breaks (DSBs) when transfected into HeLa cells. However, the molecular determinants of these effects have not been fully explored. In this study, we investigated the impact of ON structure, chemistry, delivery method, and cell type on in vitro cytotoxicity and DSBs. We found that ss PS-ONs were more cytotoxic than double-stranded (ds) PS-ONs, irrespective of the 2′-ribose chemistry, inclusive of the 2′-F modification. Cytotoxicity of ss ONs was most affected by the total PS content, with an additional contribution of 2′-F substitutions in HeLa, but not HepG2, cells. The relatively mild cytotoxicity of ds ONs was most impacted by long contiguous PS stretches combined with 2′-F substitutions. None of the tested ds 2′-F-modified PS-ONs caused DSBs, while the previously reported DSBs caused by ss 2′-F-modified PS-ONs were PS dependent. HeLa cells were more sensitive to ON-mediated toxicity when transfected with Lipofectamine 2000 versus Lipofectamine RNAiMax. Importantly, asialoglycoprotein receptor-mediated uptake of N-acetylgalactosamine-conjugated ss or ds PS-ONs, even those with long PS stretches and high 2′-F content, was neither cytotoxic nor caused DSBs at transfection-equivalent exposures. These results suggest that in vitro cytotoxicity and DSBs associated with ONs are delivery method dependent and primarily determined by single-stranded nature and PS content of ONs.
Introduction
O
To confer optimal drug-like properties (pharmacokinetics, pharmacodynamics, minimal toxicity), many of these therapeutic modalities use common chemical modifications of the phosphodiester backbone and/or of the 2′-position of the ribose moiety [3,4]. Replacement of the phosphodiester linkage by a phosphorothioate (PS), where nonbridging oxygen is substituted with sulfur, is the most common backbone modification [5,6]. Due to charge–charge interactions and the greater polarity of a sulfur versus an oxygen atom [7], PS linkages promote protein binding [8,9] leading to increased plasma half-life, reduced renal excretion [10–12], and enhanced nuclear accumulation [13,14]. In addition, PS linkages increase resistance to endo- and exonucleases and hence enhance metabolic stability of ONs [15–17].
However, PS also introduces chirality to the phosphate linkage giving rise to a large number of diastereomers (2 N , where N = number of PS linkages) that may be difficult to characterize. Undesired protein binding mediated by the PS backbone may be associated with dose-dependent liver toxicity [18], degenerative changes due to accumulation in the kidney proximal tubules, antagonism of innate immunity receptors, immunostimulatory effects, coagulopathy, and complement activation [19]. In the context of improved pharmacokinetic and nuclease stability profiles, ss ONs typically have a high number of PS modifications, while ds ONs have fewer PS substitutions since PS may also decrease affinity for the target RNA and reduce thermal stability of ds ONs [20,21], which can negatively impact potency and overall metabolic stability, respectively.
The most common modifications at the 2′-position of the ribose are 2′-O-methyl (2′-OMe), 2′-O-methoxyethyl (2′-MOE), 2′-fluoro (2′-F), locked nucleic acid (LNA), and constrained ethyl (cEt) [22]. These modifications enhance metabolic stability over RNA and DNA, and, in contrast to PS substitutions, they increase binding affinity to the complementary target RNA by favoring formation of an A-form duplex with a C3′-endo sugar pucker, thereby improving potency [6,23,24].
An additional increase in binding affinity is observed specifically with 2′-F modifications as a result of enhanced base-pairing and stacking interactions with the electronegative fluorine [25]. In addition, 2′-F is less bulky compared with other 2′ modifications and thus tends to better preserve ON compatibility with endogenous enzymes, especially when incorporated into siRNAs [26]. However, the small size and relative hydrophobicity of 2′-F can promote undesired protein binding [27]. All 2′ modifications reduce Toll-like receptor-mediated innate immunostimulatory properties [28] and provide nuclease and thermal stability [29]. 2′-F and 2′-OMe nucleosides, as well as PS backbone modifications, have been successfully integrated to optimize drug-like properties, as evidenced by FDA-approved ON drugs, pegaptanib (Macugen; 2′-F and 2′-OMe), mipomersen (Kynamro; PS and 2′-MOE), and fomivirsen (Vitravene; PS).
Among RNA-targeting silencing ONs, ASOs and siRNAs are the most clinically advanced therapeutic platforms and the most widely used research tools. They differ in structure, combination of backbone and sugar modifications, and site and mechanism of action. ASOs are single-stranded oligomers generally with lengths of ≤20 nt (<7 kDa) with a full PS backbone and a central DNA gap region of 7–14 nt flanked on both sides by 2–6 nt wings typically containing 2′-MOE, cEt, or LNA nucleosides [23,30]. ASOs engage RNase H1 predominantly in the nucleus [31–33] to direct RNA cleavage. In contrast, siRNAs are double-stranded ∼19–23-mers (∼15 kDa) with very few PS linkages and sometimes containing 2′-F and 2′-OMe modifications. siRNAs direct RNA cleavage predominantly in the cytoplasm [34] by loading into Argonaute 2 (Ago2) in the RNA-induced silencing complex.
It is generally accepted that synthetic therapeutic ONs are not genotoxic [35,36]. Importantly, all siRNA-N-acetylgalactosamine (GalNAc) conjugates in Alnylam's clinical development pipeline to date (10 programs) have tested negative in the in vitro and in vivo genotoxicity tests described in International Conference on Harmonisation (ICH) guidance S2(R1), consistent with other ONs tested to date [35,36]. However, recently a full PS ASO with 2′-F wings has been shown to be cytotoxic and cause DNA double-strand breaks (DSBs) when transfected into HeLa cells with Lipofectamine 2000 [27]. The authors speculated that ASO-induced degradation of nuclear paraspeckle proteins involved in DNA repair led to the observed toxicities.
In this study, we extended this in vitro assessment to 2′-F- and PS-modified ss and ds ONs after transfection with Lipofectamine 2000, Lipofectamine RNAiMax, or after asialoglycoprotein receptor (ASGR)-mediated uptake. We demonstrate that PS and 2′-F content in a single-stranded context is more cytotoxic than in a double-stranded context and that the magnitude of this cytotoxicity depends on the cell type and the delivery method. It is likely that DSBs reported with ss PS-ASOs were more strongly associated with the observed cytotoxicity than a genotoxic event.
Materials and Methods
Oligonucleotide synthesis
All oligonucleotides were synthesized as previously described by Nair et al. [37].
Cell culture and transfection
HeLa and HepG2 cells were cultured at 37°C, 5% CO2, in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin. Cells at 70% confluency were transfected with ONs at specified concentrations using 4 μg/mL Lipofectamine 2000 or Lipofectamine RNAiMax (Thermo Fisher Scientific) and harvested at 24 h after transfection for subsequent analyses. ON sequences are listed in Supplementary Tables S1 and S2 (Supplementary Data are available online at www.liebertpub.com/nat).
Stem-loop reverse transcription-quantitative polymerase chain reaction
To quantitate exposure to ONs, cell pellets were resuspended in phosphate-buffered saline (PBS) containing 0.25% Triton X-100, heated at 95°C for 10 min, centrifuged at 14,000 rpm at 4°C for 10 min, and reverse transcription was performed on the supernatants using the TaqMan MicroRNA Reverse Transcription Kit (Thermo Fisher Scientific) according to the manufacturer's instructions. Briefly, lysates were denatured at 95°C for 10 min, and 5 μL was immediately added to 10 μL of ice-cold master mix containing 0.5 μM RT primer. qPCR was performed on Roche LightCycler 480 II using LightCycler 480 Probes Master (Roche) according to the manufacturer's instructions. Primers (Integrated DNA Technologies) and probes (Thermo Fisher Scientific) are listed in Supplementary Table S1. Housekeeping U6 snRNA assay (Thermo Fisher Scientific) was used for normalization.
Cell survival assay
Percentage of viable cells was determined using the Cell Counting Kit-8 (Sigma) according to the manufacturer's instructions. Cells were incubated with 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt (WST-8) at 37°C, 5% CO2, for 1, 2, or 3 h; the percentage of viable cells was calculated based on absorbance at 450 nm and normalized to mock treatment.
Neutral comet assay
The neutral comet assay for detection of DNA DSBs was performed using the CometAssay kit (TREVIGEN) according to the manufacturer's instructions. Approximately 1,000 cells were plated per comet slide. DNA was stained using SYBR Gold (Thermo Fisher Scientific), imaged using a fluorescence microscope (Zeiss AxioImager.Z1), and scored using the Comet Assay IV Lite software (Perceptive Instruments).
Subcellular localization
Cells were seeded on coverslips, treated with CellLight Lysosomes-GFP, BacMam 2.0 (Thermo Fisher Scientific), and 8 h later transfected with siRNAs conjugated with Alexa647 at the 3′ end of the antisense strand using Lipofectamine 2000 or Lipofectamine RNAiMax, as described above. GalNAc-conjugated siRNAs were added to the media to allow uptake through ASGR. Cells were washed with PBS, fixed with 4% paraformaldehyde for 10 min at room temperature, washed with PBS, mounted on slides using ProLong Diamond Antifade Mountant with DAPI (Thermo Fisher Scientific), and imaged using a fluorescence microscope (Zeiss AxioImager.Z1).
Western blotting
Cell extracts were prepared in RIPA buffer (Cell Signaling Technology) supplemented with phenylmethanesulfonyl fluoride (PMSF; Cell Signaling Technology) and separated on 4%–12% Bis-Tris gradient sodium dodecyl sulfate–polyacrylamide gels (SDS-PAGE) in 1× NuPAGE MOPS SDS Running Buffer (Thermo Fisher Scientific). Proteins were transferred onto nitrocellulose membranes using the iBlot system (Thermo Fisher Scientific), blocked for 1 h at room temperature with the Odyssey Blocking Buffer (LI-COR), and incubated with primary antibodies overnight at 4°C. Anti-P54NRB (sc-376865) and anti-PSF (sc-374502) antibodies were purchased from Santa Cruz Biotechnology. Anti-PSPC1 (ab104238) was purchased from Abcam, and anti-GAPDH (D16H11) was purchased from Cell Signaling Technology. After washing with PBS containing 0.1% Tween and a 1-h incubation at room temperature with IRDye secondary antibodies (LI-COR), blots were scanned using the Odyssey imaging system (LI-COR).
Results
In vitro exposure to siRNAs, ssRNAs, and ASOs
PS and 2′-F substitutions were titrated using previously described phosphatase and tensin homolog (PTEN)-targeting ONs: 21/21-mer siRNA (siPTEN), 21-mer ssRNA, and 20-mer ASO [27], as well as an additional scrambled 21/23-mer siRNA (siSCR) (Fig. 1 and Supplementary Tables S2 and S3). siPTEN had 18 (hiF) alternating 2′-F modifications and either four (loPS) or 28 (hiPS) PS linkages. siSCR had either zero (noF) or 21 (hiF) alternating 2′-F modifications and eight (loPS), 16, or 18 (hiPS) PS linkages. ssRNA had nine (hiF) alternating 2′-F modifications, and either four (loPS) or 12 (hiPS) alternating PS linkages. ASO had either zero (noF) or 10 (hiF) 2′-F modifications and either 10 (loPS) or 19 (hiPS) PS linkages. These previously described ASOs [27] are 20-mers reminiscent of mipomersen, but shorter 16-mer cEt-modified ASOs are currently being introduced into the clinic [38,39].

Chemical composition of siRNAs, ssRNAs, and ASOs used in these studies. siSCR is a nontargeting scrambled sequence; siPTEN, ssRNA, and ASO are targeting PTEN. 2′-F, 2′-fluoro ribose modification; 2′-MOE, 2′-O-methoxyethyl; 2′-OMe, 2′-O-methyl; DNA, deoxyribonucleotide; ASO, antisense oligonucleotide; c, >10 contiguous PS linkages; PS, phosphorothioate; ssRNA, single-stranded RNA.
Intracellular exposure achieved with Lipofectamine 2000 transfection into HeLa cells or through ASGR-mediated uptake of GalNAc-conjugated ONs into HepG2 cells was quantified by stem-loop RT-qPCR at 24 h post-treatment (Fig. 2). HeLa cells were chosen to recapitulate previous findings [27], while HepG2 cells were chosen because of the highest in vitro ASGR1 surface expression among liver cell lines or hepatocytes [40]. After transfection, exposures in HeLa cells were two to fivefold higher than in HepG2 cells across ONs. Notably, in both cell lines, siRNA levels were ∼10-fold higher than ssRNA or ASO levels. In contrast to transfection, exposures resulting from ASGR-mediated uptake were equivalent for ds and ss ONs and were 500–2,500-fold lower for siRNAs and 50–100-fold lower for ssRNAs and ASOs when normalized to dose (30 nM transfection vs. 1 μM ASGR-mediated uptake). Importantly, altering the 2′-F or PS content of ONs did not affect transfection or uptake efficiency at the 24-h time point (the time point used for all subsequent experiments), verifying that any differences in cytotoxicity or DSBs for each titration series (described below) were not due to differences in exposure to parent ON.
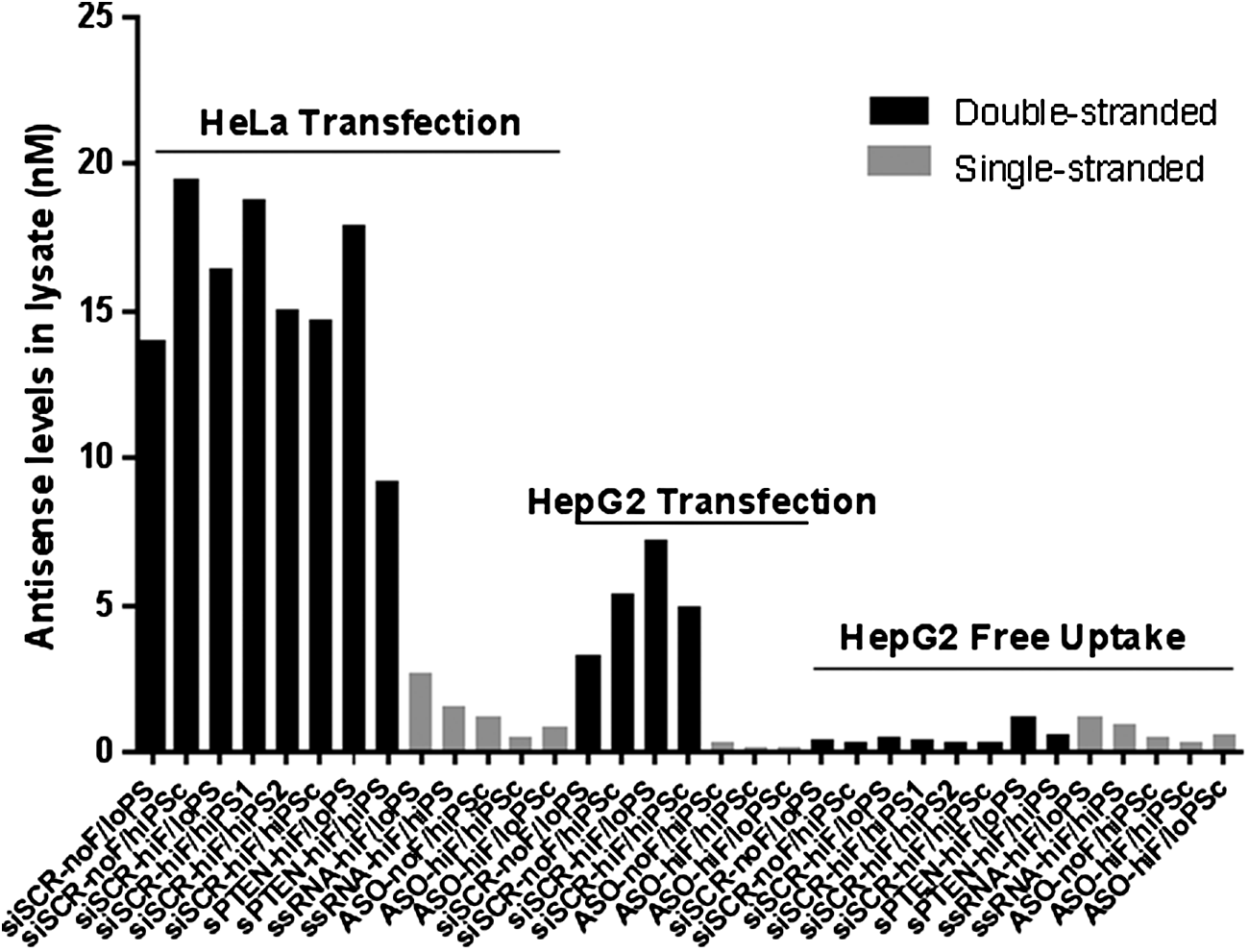
Exposure to siRNAs, ssRNAs, and ASOs after transfection into HeLa cells and HepG2 cells, or ASGR-mediated free uptake into HepG2 cells. Unconjugated siRNAs, ssRNAs, and ASOs were transfected with Lipofectamine 2000 at 30 nM, while their GalNAc-conjugated versions were delivered through ASGR at 1 μM. Exposure was assessed by stem-loop RT-qPCR at 24 h post-treatment. ASGR, asialoglycoprotein receptor; RT-qPCR, reverse transcription–quantitative polymerase chain reaction.
Effects of the 2′-F and PS content on siRNA cytotoxicity
We first examined the impact of the 2′-F modifications on siRNA-mediated cytotoxicity in the context of low PS content. siSCR-noF/loPS (zero 2′-F), siSCR-hiF/loPS (21 2′-F), and siPTEN-hiF/loPS (18 2′-F) were transfected into HeLa or HepG2 cells with Lipofectamine 2000. In both cell lines, reductions in cell viability were statistically indistinguishable between these siRNAs (Fig. 3). In HeLa cells, viability was reduced in a dose-dependent manner with LD50s ranging from 98 to 158 nM, plateauing at ∼40% reduction at ≥60 nM with all three compounds (Fig. 3A). In HepG2 cells, we observed ∼20% maximum reduction in viability for the three siRNAs at the highest dose tested (120 nM), with LD50s ranging from 375 to 655 nM (Fig. 3B), potentially due to lower exposure after transfection (Fig. 2). The top dose of 120 nM was chosen because the capacity of Lipofectamine is 50–100 nM in our hands. Because cytotoxicity and exposure to low- and high-2′-F siRNAs were comparable in each cell system (Fig. 2), these data suggest that 2′-F substitutions do not contribute to the cytotoxicity of ds ONs with a limited PS content. Cytotoxicity was not due to PTEN knockdown since the same trends were observed for siPTEN and siSCR. These findings are consistent with previous studies of 2′-F-substituted siRNAs or aptamers [41–44].
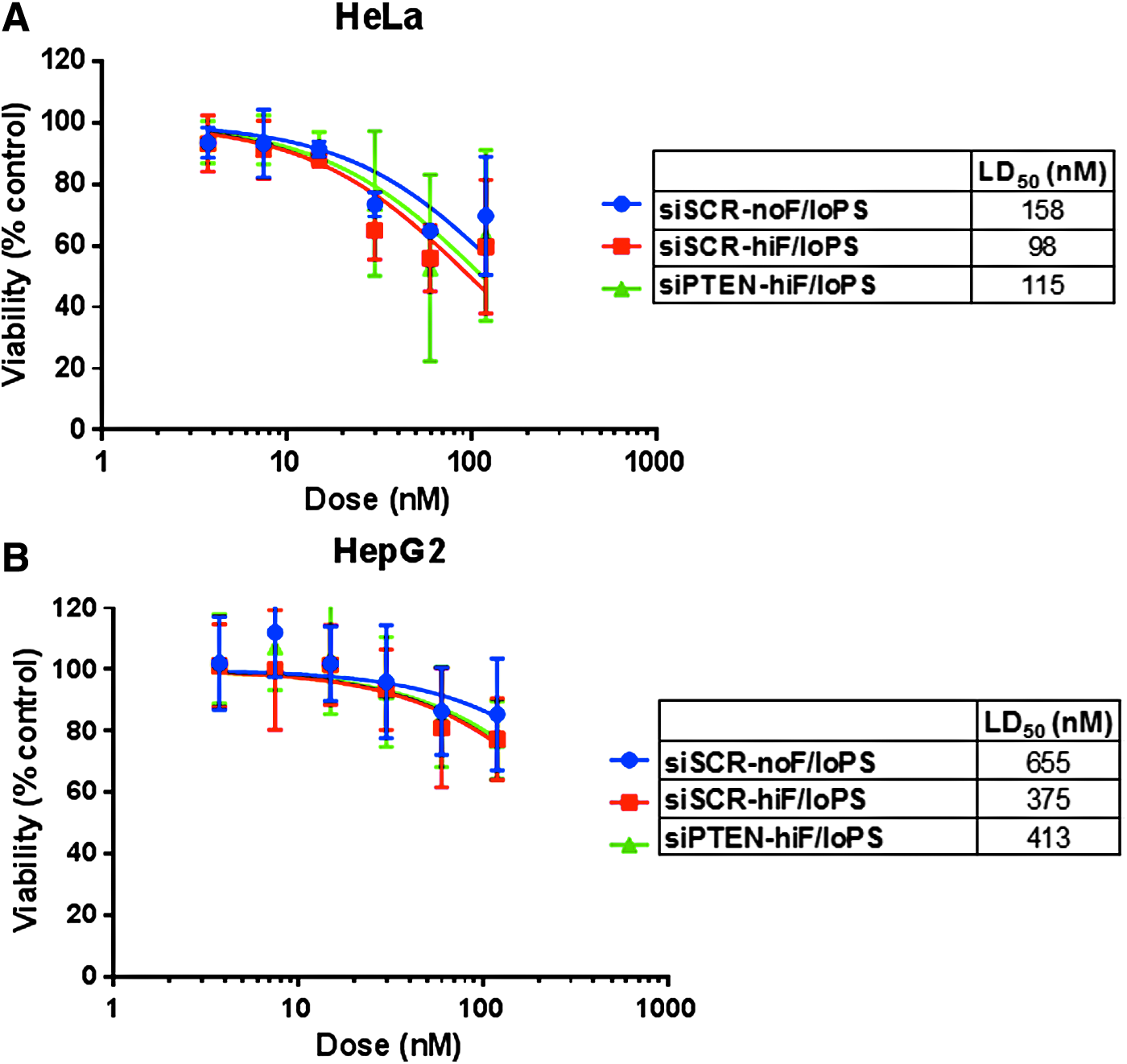
Effects of the 2′-F content on siRNA cytotoxicity in vitro. Low-PS siRNAs were transfected into HeLa
Next, we tested the effects of PS content on siRNA-mediated cytotoxicity using siRNAs containing up to 28 total and up to 10 contiguous PS linkages in the presence or absence of 2′-F modifications. Similar to the 2′-F titration experiment, we did not observe any statistically significant differences between these siRNAs. Up to 40% reduction in viability was observed in HeLa cells at ≥60 nM with LD50s, ranging from 68 to 158 nM (Fig. 4A), and up to 20% reduction in viability was observed in HepG2 cells at ≥60 nM with LD50s ranging from 171 to 655 nM (Fig. 4B). Increased cytotoxicity (by 10%–20% or approximately twofold decrease in LD50 in both cell lines) was observed only for siSCR-hiF/hiPSc, the only 2′-F-modified siRNA with 10 contiguous PS linkages, although the difference did not reach statistical significance. The PS content of these high-PS siRNAs exceeds the number of PS linkages in clinical siRNA candidates, all of which resemble the low-PS siRNAs described above.
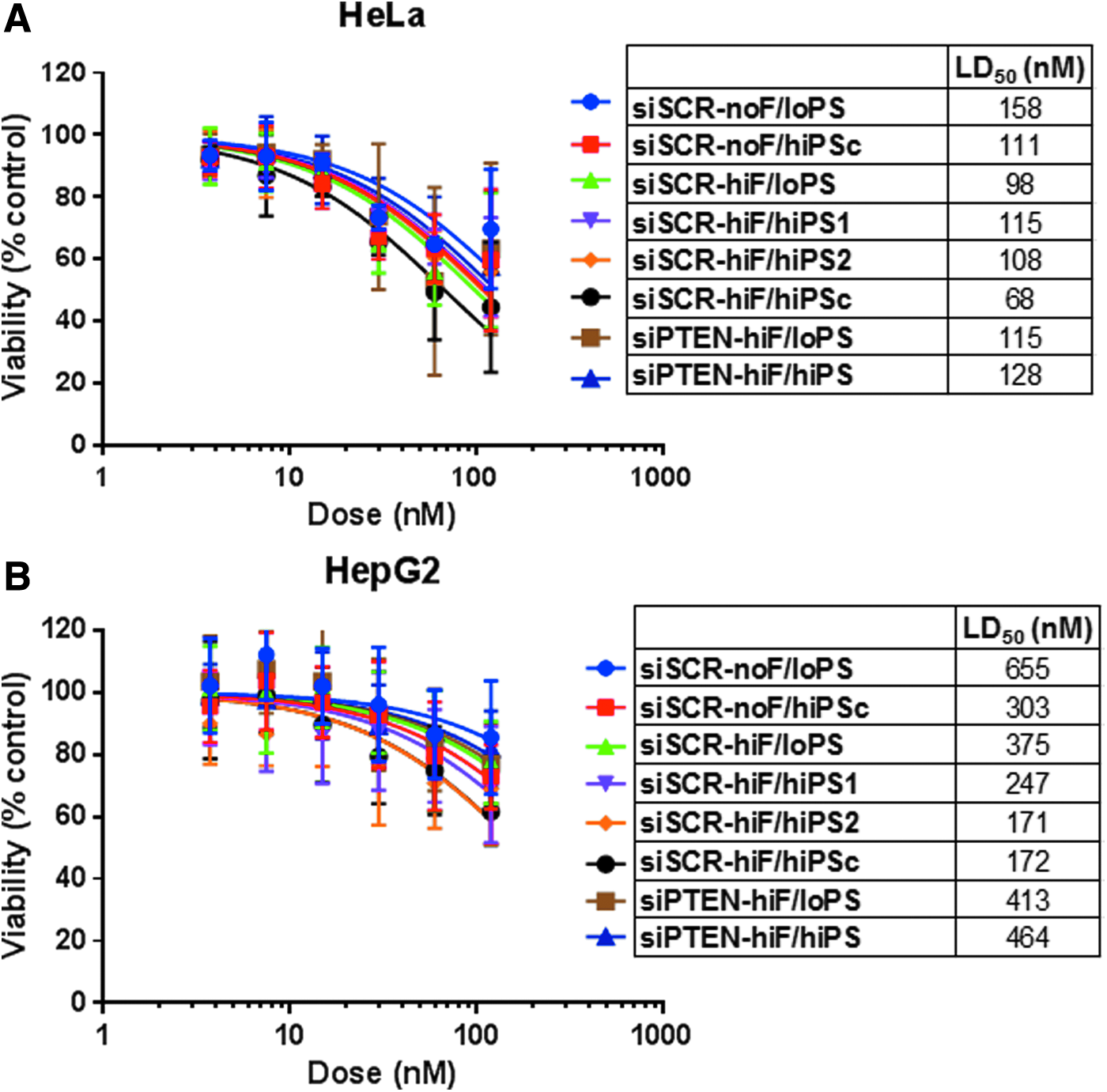
Effects of the PS content on siRNA cytotoxicity in vitro. siRNAs were transfected into HeLa
Effects of the 2′-F and PS content on ssRNA and ASO cytotoxicity
Lipofectamine 2000 transfection experiments in HeLa and HepG2 cells were also conducted with ssRNAs (Fig. 5) and ASOs (Fig. 6). The majority of these ss ONs were more cytotoxic in both cell lines compared with siRNAs, even though ∼10-fold lower exposures were achieved relative to siRNAs (Fig. 1). In contrast to 2′-F siRNAs where increased noncontiguous PS content (up to 28 PS) did not lead to increased reductions in viability, increasing PS content of a 2′-F ssRNA from 4 to 12 significantly increased cytotoxicity by 20%–30% in both HeLa and HepG2 cells, corresponding to a 2.5–4-fold decrease in LD50s (Fig. 5). These data suggest that 2′-F substitutions are not well tolerated in PS-modified ssRNAs, as reported previously for ASOs [27], whereas 2′F modifications in noncontiguous PS siRNA configurations are well tolerated and not associated with increased cytotoxicity.

Effects of the PS content on ssRNA cytotoxicity in vitro. ssRNAs were transfected into HeLa

Effects of the 2′-F and PS content on ASO cytotoxicity in vitro. ASOs were transfected into HeLa
In addition, consistent with previously published data [27], we observed increased cytotoxicity in HeLa cells for a full PS ASO when 2′-MOE wings were replaced with 2′-F wings (Fig. 6A). The viability was reduced by 60% with ASO-noF/hiPSc (LD50 of 90 nM) versus 80% with ASO-hiF/hiPSc (LD50 of 39 nM). Importantly, reducing the PS content of ASO-hiF/hiPSc from 19 to 12 (ASO-hiF/loPSc) eliminated this difference (LD50 of 74 nM) and the same transfection into HepG2 did not statistically differentiate between 2′-MOE and 2′-F PS-ASO, with LD50s ranging from 111 to 262 nM for the three ASOs (Fig. 6B). These data suggest that 2′-F- and PS-driven cytotoxicity of ASOs is cell-type dependent. While HeLa cells were sensitive to both 2′-F and PS content of ASOs, liver HepG2 cells were sensitive only to the PS content. Consistent with the ssRNA data, all ASOs were more cytotoxic than siRNAs, which had a higher overall 2′-F and PS content and a higher exposure.
Effects of the delivery method on cytotoxicity
Lipofectamine 2000 was developed for efficient intracellular delivery of DNA for protein expression and thus this cationic lipid formulation likely promotes nuclear accumulation of its payload. Indeed, siRNAs and PS-ASOs transfected with Lipofectamine 2000 do accumulate in the nucleus [45–51]. In contrast, Lipofectamine RNAiMax has been reported to be superior to Lipofectamine 2000 at siRNA transfection [52], suggesting accumulation of the payload in the cytosol where the RNAi machinery is predominantly localized (Lipofectamine RNAiMax is not recommended for DNA transfection). To visualize subcellular localization after transfection with these two formulations, we labeled siSCR-hiF/loPS with Alexa647 at the 3′ end of antisense strand (Supplementary Fig. S1). Indeed, Lipofectamine 2000 transfection led to punctate accumulation in the nucleus, while Lipofectamine RNAiMax led to a more diffuse staining throughout the cell.
We therefore considered the possibility that artificial targeting of ONs to the nucleus with Lipofectamine 2000 might explain the observed cytotoxicity of 2′-F- and PS-modified ONs. When HeLa cells were transfected with Lipofectamine RNAiMax, cytotoxicity was reduced to 10%–20% for all siRNAs (Fig. 7A) and all ssRNAs (Fig. 7B), regardless of the total 2-′F content and total or contiguous PS content, which corresponded to a 5–100-fold increase in LD50s (Supplementary Table S3). There was still significant, although reduced, cytotoxicity with ASOs: ASO-noF/hiPSc and ASO-hiF/loPSc reduced viability by 40% compared with 60% with Lipofectamine 2000, which corresponded to a 3–5-fold increase in LD50s (Supplementary Table S3). Interestingly, the 80% cytotoxicity observed with ASO-hiF/hiPSc with Lipofectamine 2000 (LD50 of 39 nM) was not reduced with Lipofectamine RNAiMax (LD50 of 48 nM), possibly because this relatively more hydrophobic ASO shuttles to the nucleus more efficiently due to enhanced protein binding relative to 2′-MOE or low-PS versions [53] even when initially delivered to the cytosol. Importantly, reduced cytotoxicity was not due to reduced exposure, which was in fact greater with Lipofectamine RNAiMax compared with Lipofectamine 2000 in most cases (Supplementary Fig. S2). These data suggest that nuclear accumulation of ONs with high 2′-F and PS content contributes to the in vitro cytotoxicity.

Effects of transfection reagents on in vitro cytotoxicity. siRNAs
In addition to different transfection agents, we also investigated cytotoxicity after ASGR-mediated uptake into HepG2 cells using GalNAc-conjugated versions of ONs shown in Figure 1. The high-affinity interaction between a GalNAc ligand and the abundant, fast-turnover, hepatocyte-specific ASGR receptor facilitates receptor-mediated endocytosis of ONs [37]. Given that ASGR-mediated uptake in vitro is much less efficient than transfection (Fig. 2), doses were increased by a factor of 67, resulting in exposures approaching those after transfection. We did not observe any cytotoxicity when ASGR-expressing HepG2 cells were treated with 2′-F- and PS-modified siRNAs (Supplementary Fig. S3). We also did not observe any cytotoxicity with ASGR-mediated uptake of ASO-noF/hiPSc or ASO-hiF/hiPSc (Supplementary Fig. S4), the two ONs that were most cytotoxic after transfection. These data support the hypothesis that intracellular trafficking can determine cytotoxicity outcomes.
DNA DSBs
A previous report suggested that increased cytotoxicity of 2′-F PS-ASOs was due to increased affinity of these ONs for nuclear paraspeckle proteins (PSF, P54NRB, and PSPC1), leading to degradation of these proteins by the proteasome [27]. Since paraspeckle proteins have roles in DNA repair, the authors proposed that their decrease could result in an increase in DSBs. However, we did not observe paraspeckle protein reductions with any 2′-F and PS-modified ss or ds ONs that were transfected at the same or higher concentrations into HeLa or HepG2 cells (Fig. 8 and Supplementary Fig. S5), indicating that paraspeckle protein reductions are not necessary to cause DSBs or cytotoxicity.
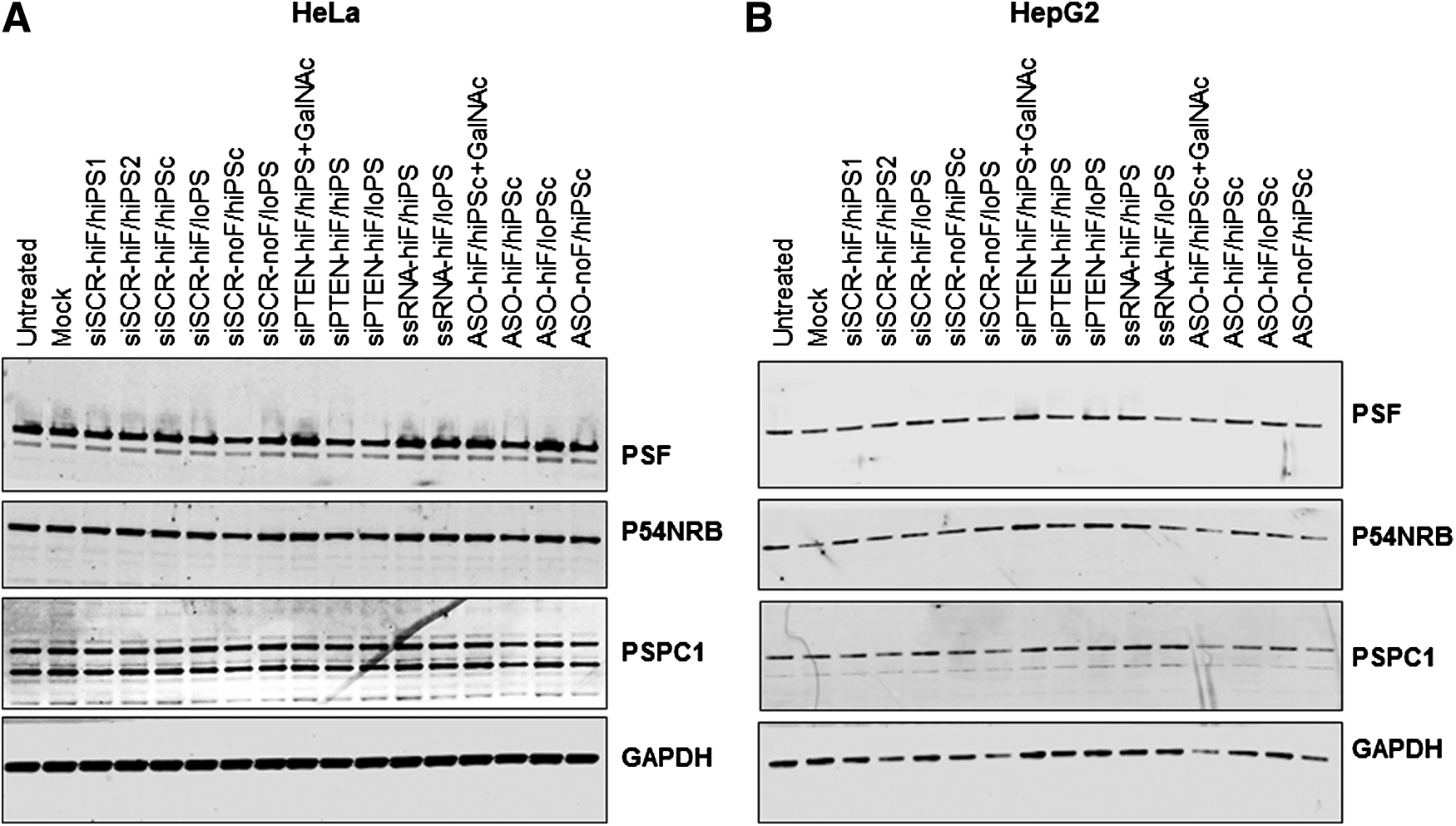
Effects of siRNA, ssRNA, and ASO transfection on paraspeckle protein levels. siRNAs, ssRNAs, and ASOs were transfected into HeLa cells at 30 nM
However, consistent with a previous report [27], we did observe increased DSBs with ASO-hiF/hiPSc transfected with Lipofectamine 2000 into HeLa cells, which manifested in increased tail intensity and tail moment in a neutral comet assay (Fig. 9). Importantly, reducing the PS content from 19 to 12 mitigated the increased incidence of DSBs, consistent with reduced cytotoxicity of ASO-hiF/loPSc. Transfection with ASO-noF/hiPSc did not result in DSBs, as expected [27]. Thus, lowering the PS content of a 2′-F-modified ASO eliminated DSBs, which can occur even in the absence of paraspeckle protein degradation.

In vitro assessment of DNA DSBs. siRNA or ASOs were transfected into HeLa cells using Lipofectamine 2000 at 30 nM, and the neutral comet assay was performed at 24 h post-transfection to assess DSBs.
In contrast to 2′-F ss ONs, transfection with siSCR-hiF/hiPSc containing 21 2′-F modifications and 16 PS linkages (including 10 contiguous) did not cause any significant increases in comet tail intensity or tail moment (Fig. 9), consistent with mild cytotoxicity compared with ASOs. Therefore, the extent of DSBs correlated well with the cytotoxicity data, was increased only for a 2′-F-modified ASO with a high PS content, and appeared unrelated to paraspeckle protein levels.
Discussion
To better understand the impact of PS linkages and 2′-sugar modifications on in vitro toxicities of siRNAs, ssRNAs, and ASOs, we evaluated the effect of PS and 2′-F content on cytotoxicity and DSBs. Our Lipofectamine 2000 transfection data (LD50 summary in Supplementary Table S3) together with the exposure data (Fig. 2) support the following cytotoxicity rank order of chemically modified ONs: hiF/hiPS ss ON > hiF/loPS ss ON = loF/hiPS ss ON > hiF/PSc siRNA > hiF/hiPS siRNA = hiF/loPS siRNA = loF/hiPS siRNA > loF/loPS siRNA. Single-stranded ONs with a moderate 2′-F content (0–10) and a moderate PS content (4–19) were more toxic than siRNAs with a higher 2′-F content (0–21) and a higher PS content (0–28).
It is possible that the relatively more polar and hydrophobic PS and 2′-F substitutions can adapt to protein structures more easily in flexible ss regions than when present in a preorganized perfectly complementary double helix, thereby facilitating and enhancing undesired protein binding. In addition, hydrophobic, charge–charge, or hydrogen bond interactions are also more likely to occur between proteins and the nucleobases and/or ribose moieties of ss ONs [54]. RNAfold analysis (http://rna.tbi.univie.ac.at/cgi-bin/RNAfold.cgi) of ssRNAs and ASOs used in these studies indicates that they can form hairpins with ss overhangs (not shown).
Increased ss ON affinity for proteins can either directly alter protein function or promote ON nuclear localization [55], possibly explaining increased DSBs, resulting in increased cytotoxicity. It is conceivable that the protein interactome differs between cell types because 2′-F PS-ASO was more cytotoxic than 2′-MOE PS-ASO in HeLa, but not in HepG2, cells. The cell-type-dependent increase in cytotoxicity is consistent with a recent report where ssRNAs, but not siRNAs, were found to be cytotoxic in HEK293 cells, but not in MCF7 or MRC-5 cells [56]. This cell-type dependency warrants further mechanistic studies as it may have important implications for systemically delivered ON therapeutics.
Although for siRNAs, both 2′-modifications (which point toward the minor groove of nucleic acid duplexes [24,57,58]) and PS linkages are exposed on the outside surface of the double helix, they may be less adaptable to protein binding. Indeed, increasing PS content (from 8 to 28) and 2′-F content (from 0 to 21) of siRNAs did not increase cytotoxicity, with a possible (not statistically significant) exception of an siRNA with 10 contiguous PS linkages in a 2′-F-substituted, but not 2′-OMe-substituted, siRNA. A stretch of PS in conjunction with 2′-F modifications may create a hydrophobic surface that promotes undesired protein binding and possibly facilitates nuclear import. Interestingly, there was no difference in cytotoxicity between siSCR-hiF/hiPS1 with 16 PS linkages mostly on 2′-F-nucleotides and siSCR-hiF/hiPS2 with 18 PS linkages mostly on 2′-OMe nucleotides, suggesting that the exact combination of a PS linkage and a 2′-modification on each monomer does not impact cytotoxicity. These high-PS designs do not resemble clinical siRNA candidates, which are characterized by low-PS content.
Effects of 2′-F- and PS-siRNAs on cell viability have been reported previously, but not in a systematic manner. siRNAs with 50% alternating PS content and no 2′-F substitution induced cytotoxicity in HeLa and mouse SW3T3 cells after transfection, while terminal PS linkages did not [59]. Importantly, siRNAs with 18 2′-F modifications (up to eight contiguous) containing zero or six PS linkages did not induce cytotoxicity in that study. Cytotoxicity was also not reported for 2′-F-modified siRNAs transfected into 293T and CD4+ T cells [41]. In addition, the RNA aptamer field has a history of safely using the 2′-F modification, suggesting that this substitution is generally well tolerated [42]. These reports are in concordance with our data that the 2′-F modification of siRNAs has a negative impact on cell viability only when combined with long contiguous PS stretches.
In our hands, in vitro cytotoxicity was dependent on the delivery route. Higher cytotoxicity was observed with Lipofectamine 2000 transfection optimized for nuclear DNA delivery than with Lipofectamine RNAiMax transfection optimized for cytosolic siRNA delivery. We observed increased siSCR-hiF/loPS nuclear punctate staining with Lipofectamine 2000 relative to RNAiMax. In addition to the transfection reagent, relatively hydrophobic 2′-F and PS substitutions themselves may promote nuclear accumulation as reported previously after a microinjection [45]. ASOs and ssRNAs may be imported to the nucleus more easily than larger siRNAs. For instance, previous reports have shown that upon transfection or microinjection, PS-ASOs accumulate in nuclear PS bodies, ASO-induced filaments, and nuclear paraspeckles [46–51], and nuclear PS body formation requires at least 10 PS bonds and is independent of 2′ modifications [53]. Therefore, factors that promote nuclear accumulation, such as transfection reagents, PS- and 2′-F substitutions, and single-strandedness may increase the likelihood of ON cytotoxicity. Lipid transfection agents should be used with caution when assessing cytotoxicity of ONs.
Importantly, no cytotoxicity was observed with ASGR-mediated uptake into HepG2 cells either for ASOs or siRNAs when comparable exposure was achieved by increasing ON doses. ASGR-mediated uptake likely results in accumulation of ONs in endolysosomal compartments [60], followed by a slow release into the cytosol by poorly understood mechanisms. To accurately assess cytotoxicity of ONs, artificial delivery methods into irrelevant cell types should be avoided because both the subcellular ON localization and cell-specific proteomes can affect cytotoxicity outcomes.
In conclusion, ASGR-mediated uptake of GalNAc-conjugated ASOs or siRNAs with various combinations of 2′-F and PS modifications is well tolerated in vitro, and the previously reported cytotoxicity for ss 2′-F-modified PS-ONs is dependent on both PS content and the transfection method. PS-ASOs with either 2′-MOE or 2′-F wings were more cytotoxic than 2′-F- and PS-containing siRNAs, likely due to differences in protein binding and the degree of nuclear accumulation. ASO cytotoxicity was highly PS and delivery route dependent, but the 2′-F dependence was cell-type specific. The absence of increased DSBs with 2′-F- and PS-modified siRNAs is consistent, with all siRNA-GalNAc conjugates in Alnylam's clinical development pipeline to date testing negative in systems of the standard test battery for genotoxicity [35,36].
Footnotes
Acknowledgments
The authors thank Drs. Natalie D. Keirstead, Martin A. Maier, and Michael E. Placke for critical reading of the manuscript.
Author Disclosure Statement
M.M.J., Y. J., M.K.S., S.W., and S.K. are employees of Alnylam Pharmaceuticals.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.