
Abstract
MicroRNAs (miRNAs) regulate gene expression by forming RNA-induced silencing complexes (RISCs) and have been considered as promising therapeutic targets. MiRNA is an essential component of RISC for the modulation of gene expression. Therefore, the release of miRNA from RISC is considered as an effective method for the inhibition of miRNA functions. In our previous study, we reported that anti-miRNA oligonucleotides (AMOs), which are composed of the 2′-O-methyl (2′-OMe) RNA, could induce the release of miRNA from RISC. However, the mechanisms underlying the miRNA-releasing effects of chemically modified AMOs, which are conventionally used as anti-cancer drugs, are still unclear. In this study, we investigated the relationship between the miRNA releasing rate from RISC and the inhibitory effect on RISC activity (IC50) using conventional chemically modified AMOs. We demonstrated that the miRNA-releasing effects of AMOs are directly proportional to the IC50 values, and AMOs, which have an ability to promote the release of miRNA from RISC, can effectively inhibit RISC activity in living cells.
Introduction
M
The miRNAs that are highly expressed in cancerous cells (oncogenic miRNA), such as miR-21, miR-221, and miR-222, are attractive therapeutic targets for cancer therapy [12, 15–18]. The main approach for the inhibition of oncogenic miRNA functions is anti-miRNA oligonucleotides (AMOs), which have the complementary sequence of the oncogenic miRNAs [19–21]. In general, AMOs are chemically modified for increased nuclease resistance and binding affinity to the target miRNA. The commonly used types of chemical modifications in AMOs are 2′-O-methyl RNA (2′-OMe) [22–24], 2′-O-methoxyethyl RNA (2′-MOE) [25], and locked nucleic acids (LNAs) [26,27]. Especially, the effectiveness of LNA-containing AMOs as anticancer drugs has been demonstrated in clinical trials. For instance, Miravirsen, which contains LNAs and targets a liver-specific miRNA (miR-122), has demonstrated antiviral activity in patients with chronic hepatitis C virus infection in clinical phase II [28,29]. Thus, AMOs have been shown as promising tools for the inhibition of miRNA activity.
MiRNA can exhibit its functions by forming RNA-induced silencing complex (RISC); therefore, it is expected that the release of miRNA from RISC effectively inactivates the RISC activity [30–32]. In our previous study, we have revealed that AMOs, which were composed of 2′-OMe RNA, could induce the release of miRNA from RISC [33]. Furthermore, we also successfully enhanced the miRNA-releasing effects of AMOs by conjugating anionic peptides to the AMOs for inhibition of the holding ability of miRNA in RISC, resulting in enhancement of inhibitory effects of AMOs on RISC activity [33]. These results indicated that release of miRNA from RISC is a promising strategy for the inhibition of RISC activity. However, the mechanisms underlying the miRNA-releasing effects of chemically modified AMOs, which are conventionally used as anticancer drugs, are still unclear. Furthermore, the relationship between the miRNA-releasing effects and the inhibitory effects of AMOs on RISC activity is also still incompletely understood. Hence, in this study, we evaluated the half-time of the releasing rate of miRNA from RISC (T1/2) and the half maximal inhibitory concentration (IC50) of AMOs on RISC activity. We also investigated the relationships between T1/2 and IC50, as well.
Materials and Methods
All AMOs described in this study were purchased from Gene Design, Inc. (Osaka, Japan).
UV melting profiles of the duplex between an AMO and mir-Luc were obtained using a UV-spectrophotometer equipped with a programmed thermal controller at an increase rate of 1.0°C/min. The sample solutions were prepared in 10 mM phosphate buffer (pH 7.0) containing 0.1 M NaCl, and the concentration of oligonucleotides was fixed at 2.0 μM.
For the dual luciferase assay, HeLa cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% (v/v) fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin at 37°C in 5% CO2. The cells were seeded in 96-well plates at a density of 4.5 × 104 cells/mL in an antibiotic-free medium, transfected and harvested for 24 h. The next day, cells were transfected with 20 nM of mir-Luc using Lipofectamine RNAiMAX (GIBCO, CA) according to the manufacturer's protocol. The following day, the cells were cotransfected with plasmids pGL4.13 (25 ng/well) and pGL4.73 (62.5 ng/well) plasmids and AMOs using Lipofectamine 2000 (GIBCO). After 24-h incubation period, the cells were lysed, and the luminescence in the lysate was measured with a Dual Luciferase Assay Kit (Promega, WI).
To prepare the immobilized RISC on beads, HEK293T cell lysate expressing hAgo2 was prepared, according to a published method [34]. For RISC assembly, 4 μL of the cell lysate was added to 3 μL of 40× reaction mix and 2 μL of mir-Luc duplex (32P-labeled, stock concentration 100 nM) and incubated at 37°C for 30 min. After incubation, the reaction mix was added to 2 μL anti-hAgo2 antibody and stirred at 4°C for 30 min. Then, 12 μL of the reaction mix was added to 20 μL Dynabeads® Protein G for immunoprecipitation (Thermo Fisher Scientific, Inc., MA) and stirred at 4°C for 2 h. To remove unbound material, the beads were washed with a washing buffer (20 mM Tris–HCl (pH 8.0), 150 mM NaCl, 2 mM MgCl2, and 0.5 mM TCEP).
For the unloading assay, 20 μL of RISC-bound beads were suspended in a 1.5 mL centrifuge tube and was preincubated at 37°C for 5 min. The unloading reaction was initiated by the addition of 1 μM (final concentration) of AMOs and subsequent incubation at 37°C. After the incubation, the beads were isolated, and the supernatant was carefully removed and named as the “unloaded miRNA” fraction. The beads were washed immediately with 40 μL washing buffer, and all traces of the supernatant were removed. The washed beads were saved and named as the “RISC (including miRNA)” fraction. The radioactivity of each fraction was measured using a radiation analyzer. The dissociation ratio of miRNA from RISC was evaluated by measuring the radioactivity.
Results and Discussion
We prepared six kinds of AMOs with sequences entirely complementary to the target miRNA sequence. As a target of AMOs, miRNA targeting firefly luciferase mRNA (mir-Luc) was selected. The melting temperature (Tm) values of AMOs duplexed with mir-Luc are summarized in Table 1. From the results, the Tm values were determined to be as follows: AMO(DNA) (53°C), AMO(DNA/OMe) (54°C), AMO(OMe) (69°C), AMO(4LNA) (76°C), AMO(DNA/LNA) (79°C), and AMO(7LNA) (88°C). These results showed that the binding affinity of AMOs containing DNA to mir-Luc was lower compared with AMOs which do not contain DNA. In contrast, AMOs containing LNA, such as AMO(4LNA), AMO(DNA/LNA), and AMO(7LNA), showed a higher binding affinity for miRNA compared with AMOs which did not contain LNA.
augc, RNA; atcg, DNA; AUGC, 2′-OMe RNA;
Tm: [mir-Luc] = [AMO]s = 2 μM in 10 mM phosphate buffer (pH 7.0), 100 mM NaCl.
First, we evaluated the miRNA-releasing effects of AMOs using the unloading assay (Fig. 1A). Time course analyses of the percentage of released miRNA from RISC by AMOs are shown in Fig. 1B. In the case of AMO(ctrl), in which the sequence was not complementary to mir-Luc, the release of the miRNA was not observed. In contrast, in the case of AMOs that have a fully complementary sequence to the target miRNA, the release of the miRNA from the RISC was observed in a time-dependent manner. The half-time of the releasing rate of miRNA from RISC by AMOs (T1/2) was determined by fitting the releasing rates as pseudo-first-order reactions (Figs. 1B and 2A). The kinetic rates of T1/2 increased in the following order: AMO(7LNA) [14 min], AMO(DNA/LNA) [20 min], AMO(4LNA) [23 min], AMO(OMe) [25 min], AMO(DNA) [49 min], and AMO(DNA/OMe) [51 min] (Fig. 1D). Figure 2B shows the plot of the kinetics rates (T1/2) versus the Tm values. We calculated the correlation coefficient and found that there was strong correlation between the Tm values and the kinetics rate (T1/2) of AMOs (R2 = 0.96). The result clearly indicates that AMOs, which have a high binding affinity for the target miRNA, can efficiently induce the release of miRNA from RISC. We consider that these results are appropriate, because association between the AMO and RISC is required to induce the release of miRNA from RISC.
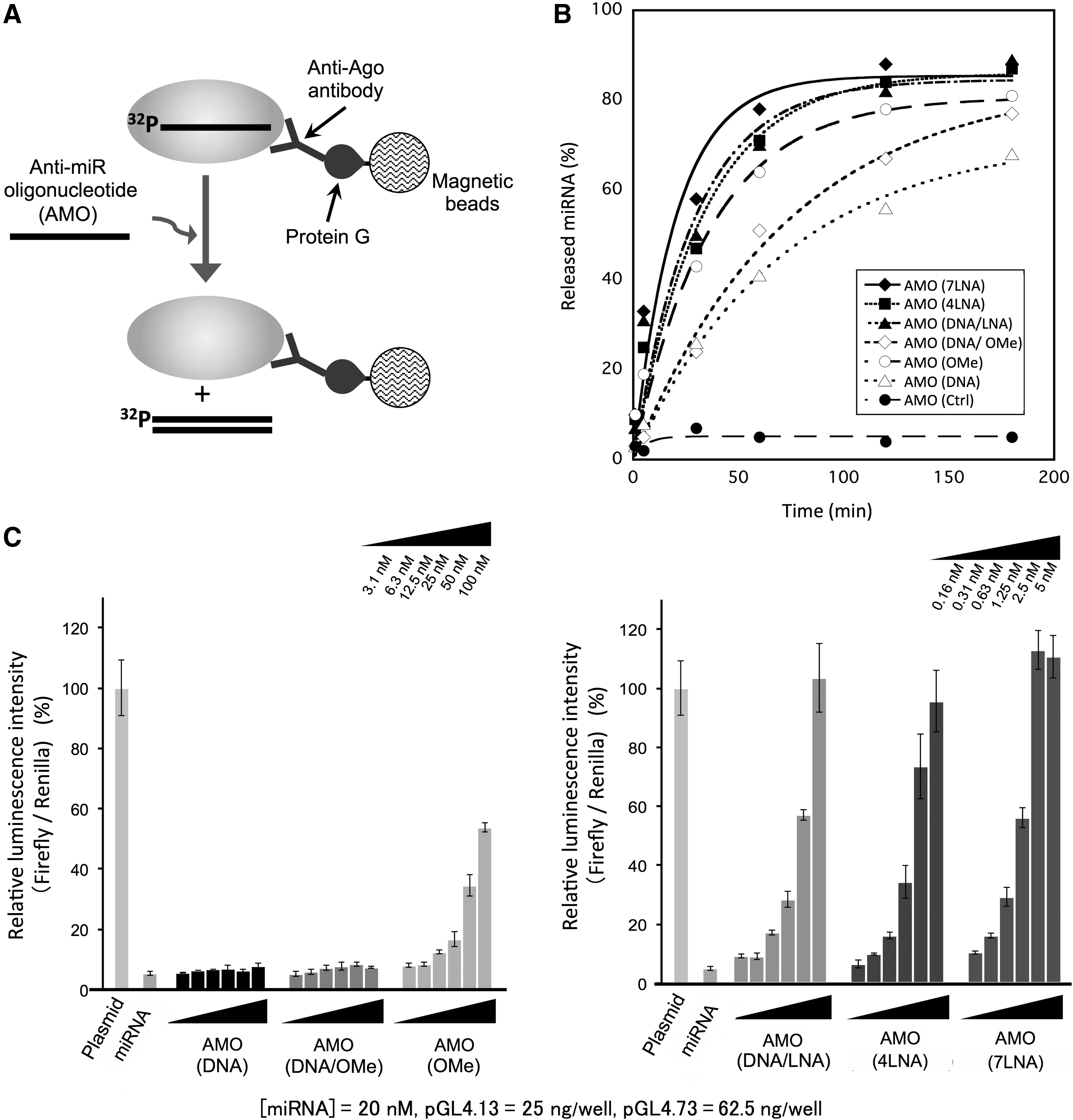
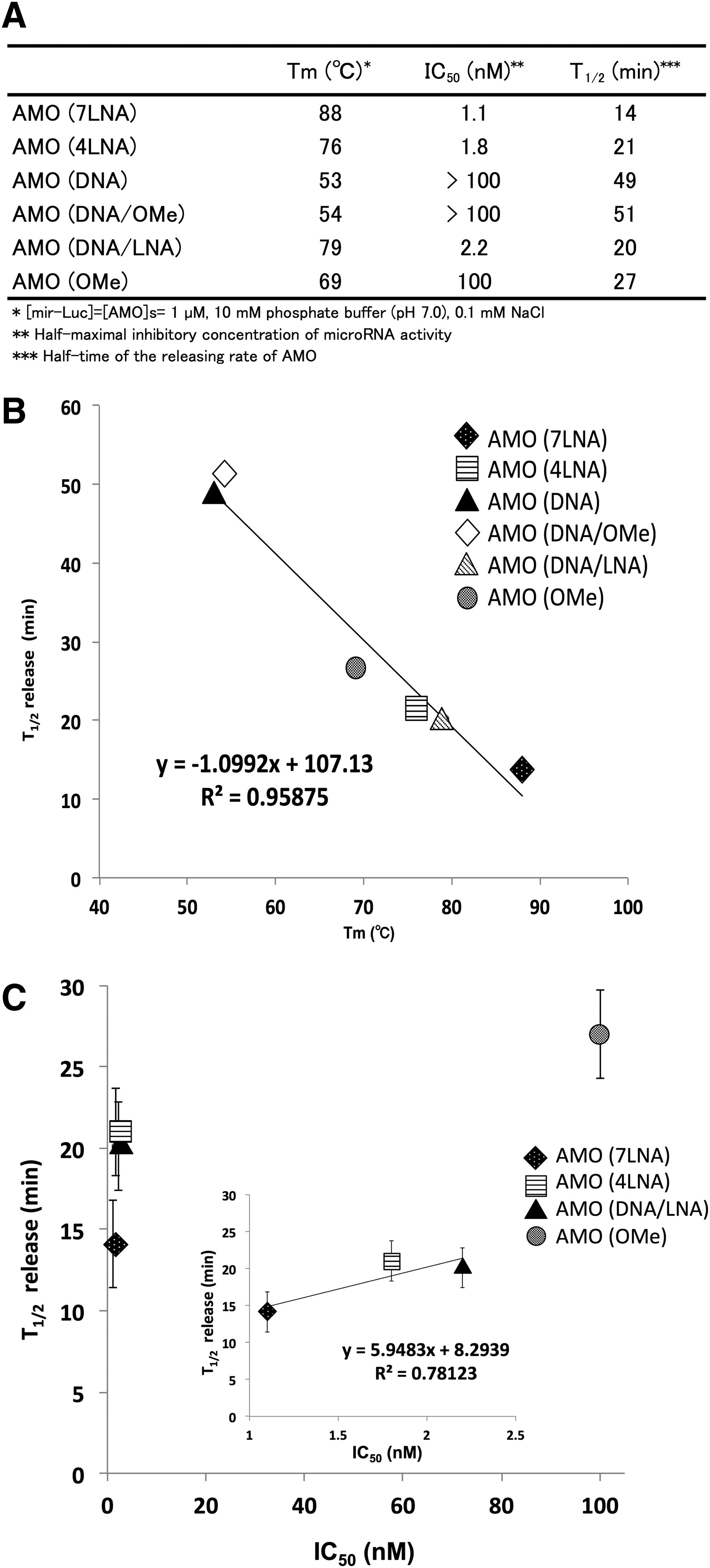
Next, we evaluated the inhibitory effect of AMOs on RISC activity in HeLa cells using the luciferase reporter assay. In this assay, luminescence intensity of firefly luciferase is already downregulated by RISC incorporated with mir-Luc. If AMOs inhibit the RISC activity, it is expected that luminescence intensity would be recovered. The results are shown in Fig. 1C. In the case of AMO(DNA) and AMO(DNA/OMe), the luminescence intensity was hardly recovered, indicating that neither of these AMOs had significant inhibitory effects on RISC functions. In contrast, concentration-dependent inhibitory effects were observed when using AMO(OMe), AMO(DNA/LNA), AMO(4LNA), and AMO(7LNA). The half maximal inhibitory concentrations (IC50) of AMOs are summarized in Fig. 2A. In order of increasing the inhibitory effect of AMOs on RISC activity, the IC50 values are determined as follows: AMO(7LNA) [1.1 nM], AMO(4LNA) [1.8 nM], AMO(DNA/LNA) [2.2 nM], and AMO(OMe) [100 nM].
Figure 2C shows the plot of the kinetics rates (T1/2) versus the IC50 values. In the case of AMO(DNA) and AMO(OMe/DNA), the IC50 values were immeasurable (IC50: >100); therefore, the IC50 values of both AMOs were not plotted in Fig. 2C. In the case of AMO(7LNA), AMO(4LNA), and AMO(DNA/LNA), there was a positive correlation between T1/2 and IC50 (R2 = 0.78); however, the plot of AMO(OMe) was deviated from the linear approximation. We speculated that AMO(OMe) hardly associates with RISC in living cells for the following reasons. In the case of unloading assay, we evaluated the releasing effect of AMOs at 1 μM (final concentration), and RISC complexes (10 fmol) were purified on magnetic beads. On the contrary, in the case of luciferase assay, AMOs were transfected into HeLa cells in the concentration range of 0.16–100 nM and we could not know exact amount of RISC in the cells. Therefore, the experimental condition of both assays was not exactly the same. Furthermore, in the case of AMO(7LNA), AMO(4LNA), and AMO(DNA/LNA), there was a positive correlation between Tm and IC50 (R2 = 0.67); however, the plot of AMO(OMe) deviated from the linear approximation (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/nat). From this result, it was considered that the miRNA-releasing from RISC by AMO(OMe) was not adequately induced in living cells. If the binding affinity of an AMO to RISC is not adequately strong in living cells, the releasing rate of the AMO could be slow in living cells. Recently, it was revealed that the transient binding between RISC and target RNA, such as mRNA and AMO, exists in the gene-silencing pathway mediated by RISC [35]. Thus, we inferred that an AMO was dissociated from RISC through the transient binding pathway when the releasing rate of miRNA by AMO was slow. From these reasons, we considered that the inhibitory effect of AMO(OMe) on RISC activity was lower compared with AMO(4LNA), AMO(7LNA), and AMO(DNA/LNA). However, there was not a linear relationship but a positive correlation between T1/2 and IC50. This result suggests that the AMOs, which can efficiently promote the release of miRNA from RISC, have high inhibitory effects on RISC activity.
To investigate the role of inhibitory effects of AMOs targeting endogenous miRNA, we evaluated the inhibitory effects of AMOs targeting miR-21 using a luciferase reporter assay and a cell growth inhibition assay. In both cases, we observed that anti-miR21 AMO(7LNA) significantly inhibited RISC activity; however, the other AMOs induced cytotoxicity (Supplementary Figs. S2 and S3). To investigate the role of the inhibitory effect of AMOs targeting endogenous miRNA, we will examine the inhibitory effects of AMOs targeting other endogenous miRNA and investigate the relationship between the inhibitory effect of AMOs on RISC activity and the releasing effect of AMOs.
Taking the results into consideration, it is clear that chemically modified AMOs induced the release of miRNA from RISC. These results suggest that the release of miRNA from RISC is directly correlated to the inhibitory effect of AMOs on RISC activity. Recently, it was reported that the AMO targeting miR-122 induced the degradation of miR-122 in vivo [36,37]. It was also reported that the inhibition of miR-21 activity using AMOs reduced the amount of miR-21 in living cells [20]. Because miRNA released from RISC can be rapidly degraded by nucleases, the above findings support our hypothesis that the release of miRNA from RISC is generally induced by AMOs in living cells.
Most importantly, we also found a proportional relationship between the kinetic rates (T1/2) and the IC50 values. These findings suggest that the strategy to release the miRNA from RISC is a promising method for the inhibition of miRNA activity in cancerous cells. It is also suggested that the miRNA-releasing effects of AMOs are valuable information for the development of AMOs as RISC inhibitors. The details of the mechanisms underlying the release of miRNA from RISC are still unclear. Understanding these mechanisms is important to develop more effective methods to inhibit RISC activity using AMOs. For instance, we previously reported that the chemical structure of wild-type RNA is favorable for the release of miRNA from RISC and that the cleavage reaction by RISC is necessary to improve the releasing rate of miRNA from RISC. These experiments suggest that chemically modified AMOs, which are partly composed of wild-type RNA to accept cleavage by RISC, might be beneficial to improve the release of miRNA from RISC and to the inhibitory effect on RISC activity of AMOs. In near future, based on our findings, we will try to develop the novel AMOs to promote the release of miRNA from RISC.
Footnotes
Acknowledgments
The authors would like to thank Prof. Hiroyuki Asanuma and Associate Prof. Yukiko Kamiya (Nagoya University, Japan) for providing the luciferase-expressing plasmid containing miR-21 binding sites and for giving us valuable suggestions.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.