
Abstract
Cholesterol (Chol) conjugation to the 5′ or 3′ end of antisense oligonucleotide (ASO) enables delivery to the liver, and Chol conjugation at the gap region can also be expected to improve delivery to the liver. In this study, we synthesized ASOs bearing the Chol-conjugated thiono triester and evaluated their activity and hepatic accumulation. We found that Chol conjugations at the gap region improved in vitro activity and hepatic accumulation when compared to unconjugated ASOs. However, Chol conjugation with phosphorothioate linkage did not improve in vivo activity in the liver, suggesting the importance of cleaving the phosphodiester between ASO and Chol. These results offer useful information for tuning the oligonucleotide structure to improve pharmaceutical properties and designing ASOs for multiple ligand conjugations and combinations with end modification.
Introduction
A
Recently, ligand conjugations have dramatically enabled successful delivery to the liver to improve the cell penetration and biodistribution of ASOs [8,9]. Cholesterol (Chol) conjugation of ASO is a well-known lipid conjugation, used since the 1990s, that greatly alters the biodistribution of the conjugated ASO and enhances its in vitro and in vivo activities [10,11].
As for the chemical structure of Chol conjugation, 3′ or 5′ end conjugation of Chol generally improves the hepatic accumulation of ASO and gene silencing activity [11,12]. Wada et al. synthesized different types of linkers for the 5′ or 3′ end of Chol conjugation to ASO and described the linker chemistry that alters their hepatic accumulation and gene silencing activity [13]. Although 5′ or 3′ end conjugation of Chol has already been reported, the number and position of Chol conjugations that can be incorporated into an ASO by the end conjugation are limited.
A few studies have focused on the synthesis of Chol conjugation at the intrachain position (gap region). Chol can be incorporated at the intrachain position using modified nucleotide building blocks such as 2′-derivatized uridine, derivatized heterocyclic bases, and a thymine amino-LNA monomer [14–17]. However, their gene silencing activity and pharmacokinetic properties are not known. Zhang et al. reported the synthesis of a gapmer ASO bearing Chol-conjugated thiono triester at the gap region and showed that it possessed an antivirus activity, but they did not report its gene silencing activity or pharmacokinetic properties [18]. Chol conjugation at the gap region may be useful for tuning the oligonucleotide structure to improve pharmaceutical properties and for designing ASOs for multiple ligand conjugations and combinations with 5′ or 3′ end modifications.
In this study, we synthesized gapmer ASOs with conjugated Chol at the gap region using thiono triester chemistry based on ethylene glycol linker tethering of Chol to PS. We also evaluated their gene silencing activity and hepatic accumulation, and compared these properties with those possessing 5′ or 3′ end conjugation of Chol.
Materials and Methods
Chemicals
Unconjugated ASO was purchased from GeneDesign, Inc. (Osaka, Japan). The sequence of the ASO is 5′-GCattggtatTCA-3′ (gapmer oligonucleotide with LNA (upper case letters) and DNA (lower case letters); all internucleoside linkages are phosphorothioated), which has the highest affinity and potency for ApoB mRNA [19]. Saline was purchased from Otsuka Pharmaceuticals (Tokyo, Japan). RNAlater was purchased from QIAGEN (Hilden, Germany). TE buffer (pH 8.0) was obtained from Nippon Gene, Co., Ltd. (Tokyo, Japan). All other reagents and solvents were commercial products of reagent grade.
Synthesis of Chol-conjugated ASOs
Chol-conjugated ASOs (Fig. 1) were synthesized using standard phosphoramidite chemistry. For the synthesis of the Chol-conjugated thiono triester, 5′-O-dimethoxytrityl(DMT)-2′-deoxyribothymidine 3′-O-(2-(2-(2-(cholesteryloxycarbonylamino)ethoxy)ethoxy)ethyl)phosphoramidite (
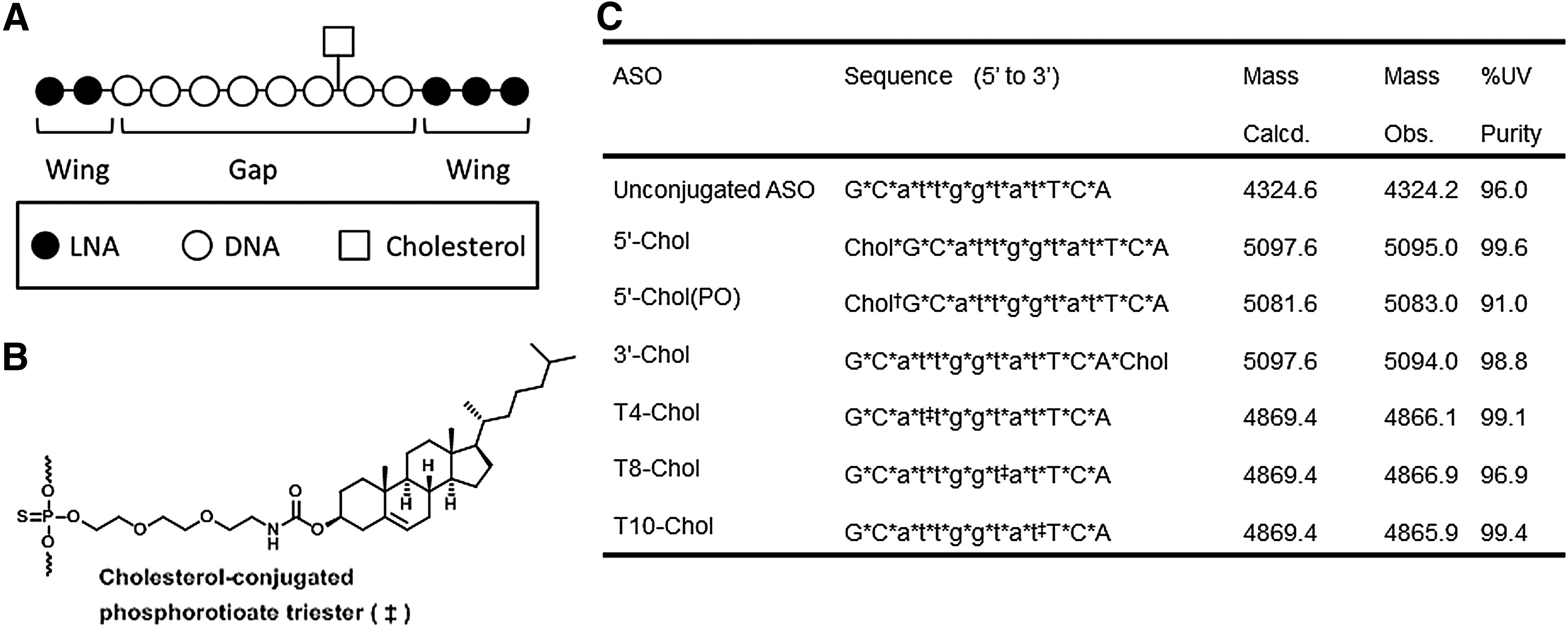
Design and structure of ASOs used in this study.
In vitro activity of ASOs
HepG2 was obtained from HSRRB (Osaka, Japan) and maintained at 37°C and 5% CO2 in Dulbecco's modified Eagle's medium (DMEM; SIGMA, Tokyo, Japan) supplemented with 10% heat-inactivated fetal bovine serum (FBS) and antibiotics. HepG2 was seeded at 2.0 × 103 cells/well in 96-well collagen-coated plates. After 24 h, ASOs at 1 μM were added, and cells were cultured in medium. After 72 h, cells were lysed using the CellAmp Direct RNA Prep Kit for RT-PCR (Takara Bio, Otsu, Japan). Quantitative PCR was performed with One Step SYBR PrimeScript (Takara Bio) using Applied Biosystems 7500 Real-Time PCR System (Applied Biosystems, Foster City, CA). ApoB mRNA levels were normalized to GAPDH and are presented relative to the untreated control.
Animal treatment
Animal care and all experimental procedures were performed with the approval of the Institutional Animal Care and Use Committee of Shionogi in terms of the 3R (Replacement/Reduction/Refinement) principle. Male C57BL/6J Jcl mice were purchased at 7 weeks of age from CLEA Japan, Inc. (Osaka, Japan). After quarantine for a week, the mice were acclimated for several days in the animal compartment. The mice were used for the experiments at 8–9 weeks of age (body weight: 22–24 g). During the acclimation and experimental periods, the mice were placed under the conditions of room temperature of 20°C–26°C, relative humidity of 30%–70%, and lighting for 12 h (light [8:00–20:00]/dark [20:00–8:00]) and allowed free access to tap water and solid laboratory food (CE-2; CLEA Japan, Inc.).
ASO was dissolved in saline, which was used as the vehicle and administered intravenously to mice (n = 3/time point) at single doses. ASOs were administered to mice at 0.1 and 0.5 mg as unconjugated ASO/kg, respectively. Whole liver samples were collected at 3 days after administration and flash-frozen on dry ice for determination of ASO amount. Some tissues were stored in RNAlater for the quantitation of ApoB mRNA. The whole liver samples were stored frozen at −80°C until analysis.
Determination of ASO amount and ApoB mRNA in whole liver
The liquid–liquid extraction is a standard method of unconjugated ASO from the liver [8,21,22]. In our previous report, the extraction efficiency of ASO conjugated with lipophilic ligands using the deproteinization method was higher than that using the liquid–liquid extraction method because the lipophilicity of ASO conjugated with lipophilic ligands is very high [8]. Therefore, we used the deproteinization method for Chol-conjugated ASO and the liquid–liquid extraction method for unconjugated ASO.
The concentrations of Chol-conjugated ASOs except for 5′-Chol (PO) in whole liver were determined by LC-MS/MS using API5000 (SCIEX; Framingham City, MA). In brief, whole liver samples (50–100 mg tissue) were homogenized in extraction buffer (0.5% Nonidet P-40, 25 mM ethylenediaminetetraacetic acid, 100 mM sodium chloride, 25 mM tris hydroxymethyl aminomethane, 0.5 mg/mL proteinase K, pH 8.0) and incubated at 37°C for 0.5 h. In case of preparation of standard, the blank whole liver samples spiked with the standard solutions were homogenized as described above. Fifty microliters of the above homogenate samples was added to 50 μL of 300 mM triethylamine and mixed for 5 min. The mixtures were added to 250 μL of isopropanol and mixed for 5 min. After the centrifugation of mixture, 10 μL of the supernatants was injected into the LC-MS/MS system. Chromatographic separations were performed at a flow rate of 0.4 mL/min on Cadenza CD-C18 HT (2.0 × 50 mm, 3 μm; Imtakt Corporation, Kyoto, Japan) for 4 min. A binary gradient was used to perform the separations. Mobile phase A consisted of 10 mM diisopropylamine (DIPA) and 25 mM 1,1,1,3,3,3-hexafluoro-2-propanol (HFIP) in water, and mobile phase B consisted of 10 mM DIPA and 25 mM HFIP in methanol/acetonitrile (1/1). The column temperature was maintained at 60°C using a column heater. Mass determination was performed by MRM mode using m/z 1018.5/94.9 for 5′-Chol, and m/z 1216.4/94.9 for T4-Chol, T8-Chol, and T10-Chol. The calibration standards of Chol-conjugated ASO ranged from 0.3 to 100 μg as unconjugated ASO/g. The precision and accuracy of the calibration standards were within 20%.
The amount of intact 5′-Chol (PO) was already lower than that of unchanged ASO in the whole liver 24 h after the administration [8]. Therefore, we performed the determination of unconjugated ASO, which is an active form, for the whole liver after the administration of 5′-Chol (PO). The concentrations of unconjugated ASO in the whole liver were determined by LC-MS/MS using API5000. In brief, whole liver samples (50–100 mg tissue) were homogenized in extraction buffer and incubated at 37°C for 0.5 h. To prepare the standard, the blank whole liver spiked with the standard solutions was homogenized as described above. ASO in the above homogenate samples was extracted by the liquid–liquid extraction method using phenol/chloroform (1/1, v/v), followed by solid-phase extraction (Oasis HLB, 10 mg; Waters, Inc., Milford, MA). The eluant was evaporated to dryness under nitrogen and reconstituted in TE buffer (pH 8.0)/methanol (9/1, v/v). Five microliters of reconstituted solution was injected into the LC-MS/MS system. Chromatographic separations were performed at a flow rate of 0.4 mL/min on a Cadenza CD-C18 HT (2.0 × 50 mm, 3 μm; Imtakt Corporation, Kyoto, Japan) for 4 min. A binary gradient was used to perform the separations. Mobile phase A consisted of 10 mM DIPA and 25 mM HFIP in water, and mobile phase B consisted of 10 mM DIPA and 25 mM HFIP in methanol/acetonitrile (1/1). The column temperature was maintained at 60°C using a column heater. Mass determination was performed by MRM mode using m/z 719.9/94.9 for unconjugated ASO. The calibration standards ranged from 0.03 to 30 μg/g for whole liver. The precision and accuracy of the calibration standards were within 20%.
Total RNA was extracted from whole liver using the RNeasy 96 Universal Tissue Kit (QIAGEN). One microgram of total RNA was converted to cDNA using SuperScript™ III First-Strand Synthesis SuperMix for qRT-PCR (Invitrogen, Carlsbad, CA), according to the manufacturer's instructions. Quantitative PCR was performed with SYBR® Premix Ex Taq II (Takara Bio) using Applied Biosystems 7500 Real-Time PCR System. ApoB mRNA levels were normalized to GAPDH and are presented relative to the saline control.
Data analysis
All data represent mean ± standard deviation (SD). The Dunnett test was used to compare values between groups. P values <0.05 were considered significantly different. Statistical probability (P) in figures and figure legends are expressed as *P < 0.05 and **P < 0.01.
Results and Discussion
Effect of thiono triester on the in vitro efficacy of cholesterol-conjugated ASOs
First, we designed Chol-conjugated thiono triester for gap modification by the O-alkyl phosphoramidite method (Fig. 1A, B) because O-alkyl phosphoramidite can be synthesized from alcohol and 5′-O-DMT-2′-deoxyribonucleoside in a single one-pot reaction and adapted to four nucleosides (adenine, guanine, thymidine, and cytidine), unlike other internucleotide modifications such as the modified base method [20]. This parallel feature of the synthesis is suitable for incorporating Chol at any phosphate position of the ASO. As for the linker between thiono triester and Chol, we selected an ethylene glycol-based linker instead of a hydrophobic linker because the former would enable more expression of ASO activity compared to a hexamethylene succinimide linker [13]. We synthesized O-alkyl phosphoramidite with thymidine base and incorporated 8-mer (T8-Chol) and 10-mer (T10-Chol) from 5′ end of ASO (Fig. 1C).
We compared the in vitro activity of the Chol-conjugated ASOs in a cell line (Fig. 2A). Surprisingly, the knockdown activities of Chol-conjugated ASOs were enhanced compared to the unconjugated ASO. The knockdown activity of T10-Chol was the highest among the Chol-conjugated ASOs, and that of 5′-Chol was higher than that of 3′-Chol.
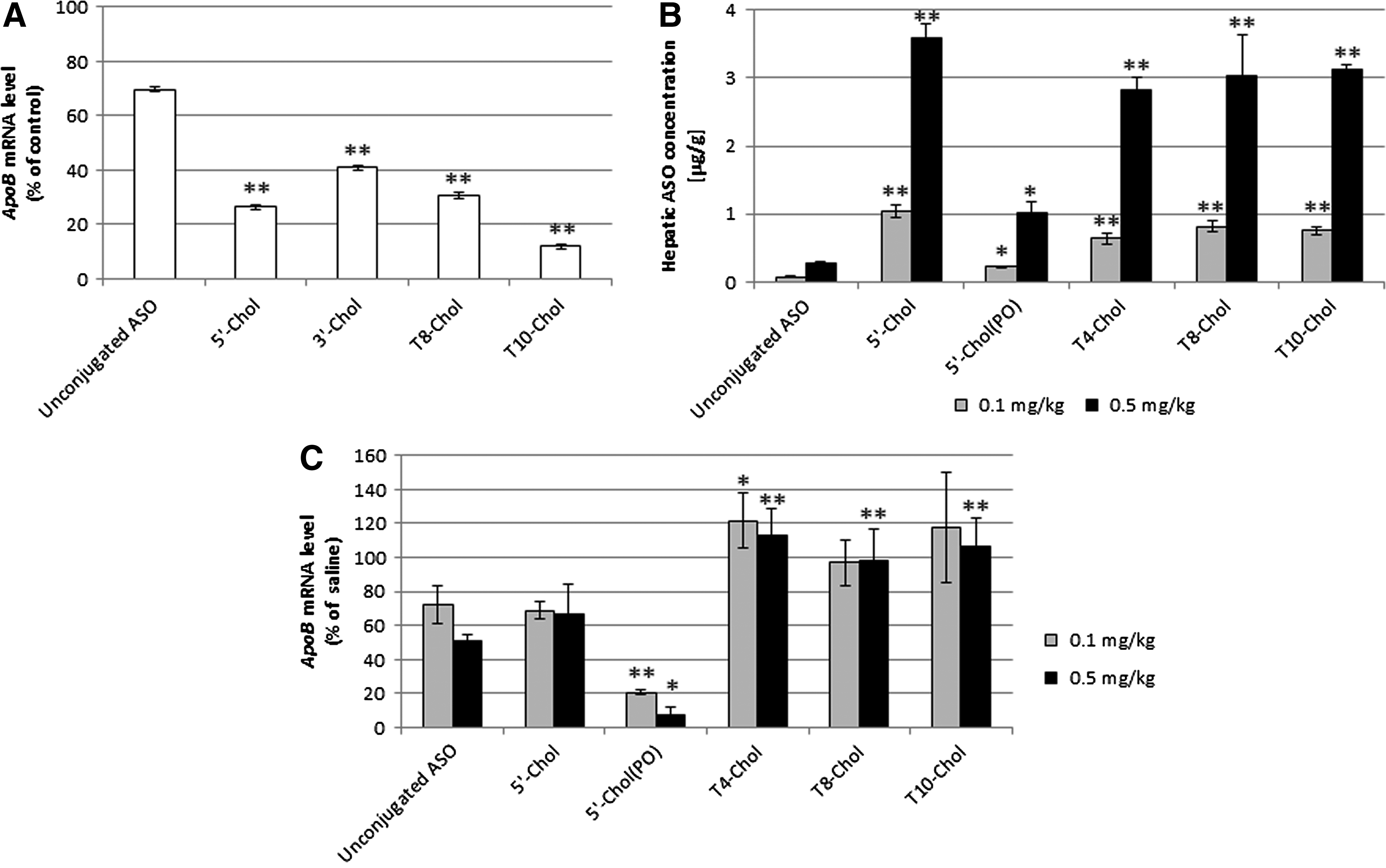
Inhibitory activities and hepatic accumulation of modified ASOs. ASOs were transfected into HepG2. After 72 h, ApoB mRNA
Letsinger et al. previously reported that the in vitro activity of Chol-conjugated PS was higher than unconjugated ASO due to improved cell penetration with high lipophilicity [10]. Our Chol-conjugated ASOs could also improve the in vitro activity with their high lipophilicity in the same manner. As for the position of the conjugation, the activity of ASO with Chol on the gap region was comparable to that with Chol on the end. To maximize the in vitro activity of Chol-conjugated ASO, it might be good to explore the gap region using thiono triester in addition to the end position.
Chol conjugation enhances ASO delivery to the liver
To evaluate the effect of the position of Chol conjugation on hepatic accumulation, we administered ASOs with Chol at the gap region (T4-Chol, T8-Chol, and T10-Chol) and at the end (5′-Chol) to mice. In addition, we examined T4-Chol to evaluate the effect of the position of conjugation on hepatic accumulation.
The amounts of intact ASO after administration of 5′-Chol, T4-Chol, T8-Chol, and T10-Chol were 10- to 15-fold higher than that after administration of unconjugated ASO at the same doses (Fig. 2B). The amounts of intact ASO after administration of ASOs conjugated with Chol at the gap region (T4-Chol, T8-Chol, and T10-Chol) were comparable with ASO conjugated with Chol at the end (5′-Chol). We also investigated the delivery of Chol conjugation with the PO linkage (5′-Chol [PO]), but not 3′-Chol because the in vitro activity of 5′-Chol was higher than that of 3′-Chol. The amount of unconjugated ASO after administration of 5′-Chol (PO) was three- to fourfold higher than that after unconjugated ASO.
This is the first report that ASOs conjugated with Chol at the gap region can enhance the delivery of ASO to the liver to an extent similar to that by ASO conjugated with Chol at the end. Therefore, Chol conjugation at the gap region may be useful for tuning the oligonucleotide structure to improve pharmaceutical properties and for designing ASOs for multiple ligand conjugations and for combinations with 5′ or 3′ end modifications.
In vivo activity of ASO is improved by cholesterol PO linkage but not by cholesterol PS linkages
The in vivo activities of 5′-Chol, T4-Chol, T8-Chol, and T10-Chol were not enhanced compared to unconjugated ASO (Fig. 2C). However, comparison of different doses (0.1 and 0.5 mg/kg) showed that the activity of 5′-Chol (PO) was more than fivefold higher than that of unconjugated ASO.
Although in vitro activities and hepatic accumulations of Chol-conjugated ASOs (5′-Chol, T4-Chol, T8-Chol, and T10-Chol) were higher than those of unconjugated ASO, no enhancement of in vivo activities was observed. On the contrary, the in vivo activity of 5′-Chol (PO), an analogue of 5′-Chol with a phosphodiester linkage, was higher than that of unconjugated ASO although its delivery to the liver was lower than those of the other Chol-conjugated ASOs. It is known that the phosphodiester bond between ASO and Chol is promptly cleaved in the liver to liberate unconjugated ASO [8]. Our results for the conjugation at the gap region suggest that it is important for the in vivo activity to promptly cleave the phosphodiester bond between ASO and Chol, as well as the conjugation at the end [13]. With respect to modification of the gap region using thiono triester or phosphonate triester, several types of biodegradable linkage such as S-acyl-2-thioethyl phosphotriester, GSH-sensitive benzyl protecting groups, and reduction-activated protecting groups have been developed [23–25]. Meade et al. showed that the in vivo activity of these oligonucleotides was higher than that of unconjugated oligonucleotides [23]. The in vivo activity of our conjugation at the gap region may be improved by combinations of these linker designs or PO moiety insertion to the linker.
In conclusion, we demonstrated that Chol conjugations at the gap region could improve in vitro activity and its hepatic accumulation as well as observed with the conjugation at the end. However, Chol conjugation with PS linkage could not improve in vivo activity in the liver, suggesting that it is important for the in vivo activity to promptly cleave the phosphodiester bond between ASO and Chol. These results offer useful information for tuning the oligonucleotide structure to improve pharmaceutical properties and for designing ASOs for multiple ligand conjugations and combinations with end modification.
Footnotes
Acknowledgment
The authors greatly appreciate the support from Dr. Shuji Yonezawa (Shionogi & Co., Ltd.).
Author Disclosure Statement
The authors declared no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.