
Abstract
Specific gene silencing through RNA interference (RNAi) holds great promise as the next-generation therapeutic development platform. Previously, we have shown that branched, tripodal interfering RNA (tiRNA) structures could simultaneously trigger RNAi-mediated gene silencing of three target genes with 38 nt-long guide strands associated with Argonaute 2. Herein, we show that the branched RNA structure can trigger effective gene silencing in Dicer knockout cell line, demonstrating that the Dicer-mediated processing is not required for tiRNA activity. The finding of this study confirms the flexibility of the structure of RNAi triggers as well as the length of the guide strand in RNAi-mediated gene silencing.
Introduction
S
However, several recent studies reported diverse structural repertoires of RNAi-triggering molecules for efficient target gene silencing. Moreover, some of the siRNA structural variants showed improved features, including increased potency, reduced nonspecific responses, and enhanced cellular delivery, when compared with the conventional siRNAs [4–6]. As a category of these structural variants, several reports described novel siRNA structures that can induce silencing of multiple target genes. Instead of linear siRNA structure, the multigene targeting siRNA molecules have nonlinear, branched structures and showed advantages over the classical siRNA structure, including enhanced and prolonged target gene silencing [7–11].
Our group developed a multimeric siRNA structural variant called tripodal interfering RNA (tiRNA), which has a branched, nonlinear tripartite structure built by annealing three 38-nt single-stranded (ss) RNAs, in such a way that each duplex region contained 19 bp of siRNA duplex [7,8]. In addition to triggering simultaneous silencing of three difference target genes, the initial analysis of the tiRNA activity suggested that Dicer-mediated processing might be dispensable for tiRNA-mediated target gene silencing. The introduction of partial chemical modification into the tiRNA junction region supposed to be the target site of the Dicer enzyme did not affect the target gene silencing activity of tiRNA. We also showed that the intact 38-nt-long antisense strand of tiRNA could be directly associated with Ago2 protein. These results imply that intact tiRNA can be incorporated into an active RNA-induced silencing complex and trigger RNAi without Dicer processing. In this study, further analysis on Dicer-independent tiRNA activity was conducted using genetically engineered Dicer knockout cell line.
Materials and Methods
RNAs
Chemically synthesized RNA oligonucleotides for siRNAs, tiRNAs, and miRNAs were purchased from Dharmacon and annealed according to the manufacturer's protocol. RNA sequences are shown hereunder. For let-7c miRNA, pre-miRNA (AM17100/PM10436) and mature form miRNA (4464066/MC10436) were purchased from Thermo Fisher Scientific. No chemical modification was introduced into RNA oligonucleotides used in the study.
siSurvivin (Survivn targeting siRNA)
Sense: 5′-AUGCAUGACUUGUGUGUGA-3′
Antisense: 5′-UCACACACAAGUCAUGCAU-3′
siMET (c-MET targeting siRNA)
Sense: 5′-CGAGAUGAAUGUGAAUAUG-3′
Antisense: 5′-CAUAUUCACAUUCAUCUCG-3′
siCTNNB (β-catenin targeting siRNA)
Sense: 5′- GUAGCUGAUAUUGAUGGAC-3′
Antisense: 5′- GUCCAUCAAUAUCAGCUAC-3′
siLamin (Lamin A/C targeting siRNA)
Sense: 5′-CUGGACUUCCAGAAGAACA(dTdT)-3′
Antisense: 5′-UGUUCUUCUGGAAGUCCAG(dTdT)-3′
siDBP (DBP targeting siRNA)
Sense: 5′-UCGAAGACAUCGCUUCUCA(dTdT)-3′
Antisense: 5′-UGAGAAGCGAUGUCUUCGA(dTdT)-3′
siTIG3 (TIG3 targeting siRNA)
Sense: 5′-CUGUCUCAGGCGUUCUCUA(dTdT)-3′
Antisense: 5′-UAGAGAACGCCUGAGACAG(dTdT)-3′
tiSMC
First strand (Survivin-c-MET):
5′-UCACACACAAGUCAUGCAUCGAGAUGAAUGUGAAUAUG-3′
Second strand (c-MET-β-catenin):
5′-CAUAUUCACAUUCAUCUCGGUAGCUGAUAUUGAUGGAC-3′
Third strand (β-catenin-Survivin):
5′-GUCCAUCAAUAUCAGCUACAUGCAUGACUUGUGUGUGA-3′
tiLDT
First strand (Lamin A/C-DBP):
5′-UGUUCUUCUGGAAGUCCAGUCGAAGACAUCGCUUCUCA-3′
Second strand (DBP-TIG3):
5′-UGAGAAGCGAUGUCUUCGACUGUCUCAGGCGUUCUCUA-3′
Third strand (TIG3-Lamin A/C):
5′-UAGAGAACGCCUGAGACAGCUGGACUUCCAGAAGAACA-3′
Plasmids
DNA oligonucleotides corresponding to the antisense strand of Survivin, β-catenin, c-MET and Lamin A/C, DBP, TIG3, and miR-let7c (mature form) were cloned into the 3′-UTR of the pMIR-REPORT Luciferase vector (Ambion) using the SpeI and HindIII cloning sites. The sequence information for each DNA oligonucleotide is available upon request.
Cells
HeLa (CCL-2) cell line was purchased from ATCC and cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco) supplemented with 10% fetal bovine serum (FBS; Gibco). Dicer1(−/−) mesenchymal stem cells (CRL-3221) lack expression of the Dicer1 protein, and wild-type control Dicer1(f/f) (CRL-3220) cells were purchased from ATCC and cultured in minimum essential medium (MEM) α (Gibco) supplemented with 10% FBS.
Western blot
The whole cell extracts were lysed with lysis buffer containing protein inhibitor. The protein concentration of the cell extracts was measured by bicinchoninic acid assay. Equal amounts of protein (30 μg) were prepared and subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis. After the electrophoresis, proteins were transferred to polyvinylidene difluoride membranes using a submerged transfer apparatus (Bio-Rad). The membranes were blocked with 5% skim milk in TBS-T (50 mM Tris-HCl at pH 8.0, 150 mM NaCl, and 0.1% Tween 20). After overnight incubation at 4°C with a 1/2,000 dilution of a rabbit monoclonal antibody to Dicer (Novus Biologicals), or a 1/400 dilution of a mouse polyclonal antibody to Tubulin (Santa Cruz Biotechnology) as the loading control, the membranes were washed in 0.05% Tween-20 in PBS and incubated with a 1/5,000 dilution of goat antirabbit or goat antimouse horseradish peroxidase-conjugated antibody in blocking buffer (Santa Cruz Biotechnology). Membranes were washed and developed using enhanced chemiluminescence detection system (Bio Rad).
Transfection and luciferase assay
Cells were plated on 24-well plates at 50%–60% cell density in complete medium. After 24 h, 100 ng of pMIR-REPORT-based plasmids containing the antisense-strand target sequence, 1 ng of Renilla luciferase vector (pRL-SV40; Promega), and RNAi triggering molecule such as tiRNA, siRNA mixture, and miRNA were cotransfected using Lipofectamine 2000 (Invitrogen) reagent following the manufacturer's protocol. Twenty-four or 45 h after the transfection, cells were lysed using passive lysis buffer (Dual-Luciferase Reporter Assay System; Promega). Firefly and Renilla luciferase activity was then measured in 20 μL of the cell extract using multiwell plate reader Victor X2 (PerkinElmer). The Firefly luciferase signal was normalized to the Renilla luciferase signal, and the silencing efficacy was calculated relative to the luciferase activity.
Ago2 incorporation assay
Dicer1(f/f) and Dicer1(−/−) cells were grown in MEM α (Welgene) supplemented with 10% FBS (Welgene). HEK293T cells were maintained in DMEM (Welgene) supplemented with 10% FBS. The day before the transfection, 1 × 106 cells were seeded in 100-mm plates to achieve ∼50% confluency by the time of transfection. Cells were cotransfected with 10 nM of siRNA or tiRNA and 10 μg of pcDNA3.1-FLAG-hAGO2 using Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. After 48 h, cells were washed twice with ice-cold 1× phosphate-buffered saline and harvested by scraping. The cells were resuspended in NET-2 buffer [100 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.05% NP-40, 1 mM DTT, 1× protease inhibitor cocktail (Roche), 1 U/mL RNaseOUT (Invitrogen)] and incubated on ice for 20 min. The lysate was further sonicated on ice for 1 min and centrifuged at 21,200 g at 4°C for 10 min. The supernatant was incubated with 15 μL of anti-Flag antibody-conjugated agarose beads (Sigma) with constant rotation at 4°C for 2 h. The beads were washed four times with NET-2 buffer and treated with TRI Reagent (Ambion) to extract coimmunoprecipitated RNAs. The RNAs recovered from the input or Flag-immunoprecipitate samples were resolved on a 12.5% urea–polyacrylamide gel, transferred to a Hybond N+ membrane (GE Healthcare), and UV crosslinked. DNA oligonucleotides complementary to the antisense strand sequence of siLamin or let-7a-5p were radiolabeled at the 5′ ends and used as the probes for Northern blotting. The membrane was exposed to a phosphorimager and read using the BAS-2500 system (Fujifilm).
Results and Discussion
Gene silencing by tiRNA in HeLa cells
First, a tiRNA composed of three 19 bp siRNA units targeting Survivin, β-catenin, and c-MET was designed (tiSMC; Fig. 1A). To analyze the target gene inhibition activity of tiRNA, three luciferase reporter vectors harboring the target sequence of each antisense strand of tiSMC were constructed. Cotransfection of each reporter vector and tiSMC into HeLa cells for 24 h and analysis of the target knockdown showed that the tiSMC induces multiple target gene silencing activity, as shown in Fig. 1B. In addition, we performed dose-dependent endogenous target gene silencing analysis of the tiRNA and the result showed multiple target gene silencing activity of tiSMC (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/nat).
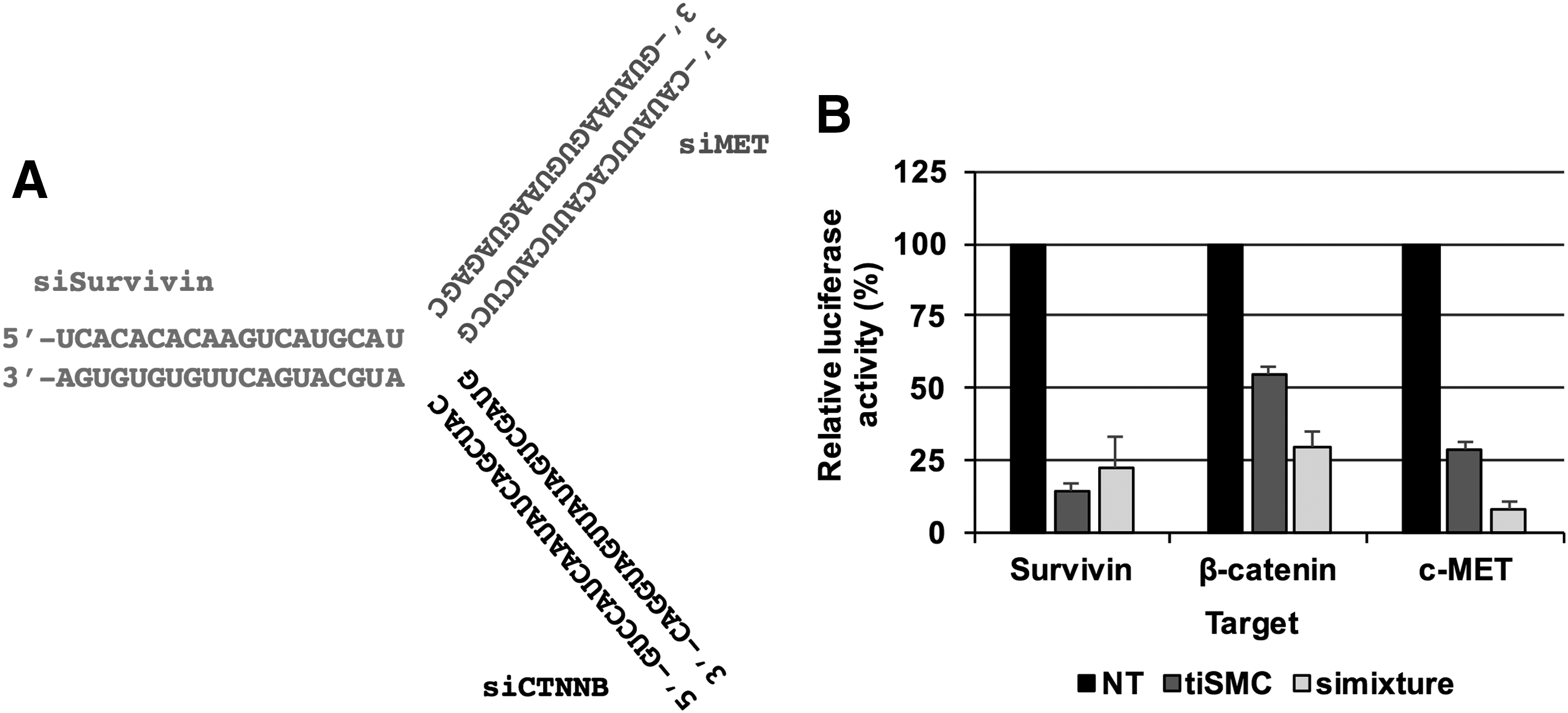
Validation of multiple gene targeting tiRNA.
Analysis of target knockdown of tiRNA in Dicer knockout cell line
Dicer is regarded as a key factor in the endogenous RNAi pathway and functions as a molecular ruler to produce dsRNA fragments 25 nt in length [12]. As the tiRNA structure constructed by annealing of three 38 nt strands has unique features including three arms with 19 bp dsRNA segment and a central junction region (Fig. 1A), it is not clear whether tiRNA is processed by Dicer when introduced into cells. To unambiguously answer this question, we examined target gene knockdown of tiRNA in Dicer knockout cell line termed as Dicer1(−/−), a model of homozygous Dicer1 deletion in murine mesenchymal stem cells established from an adult Dicer1(f/f) mouse and immortalized in vitro [13,14]. Western blot analysis was conducted for Dicer1(f/f) and Dicer1(−/−) cells and Dicer protein was detected in Dicer1(f/f) cells, but not in Dicer1(−/−) cells (Fig. 2A). Functional validation of Dicer deficiency in Dicer1(−/−) cells was also conducted. Luciferase reporter harboring the target sequence of mature miR-let7c along with pre-let7c (precursor of miR-let7c) was introduced into both Dicer1(f/f) and Dicer1(−/−) cells, and luciferase activity was analyzed. As shown in Fig. 2B, knockdown of luciferase activity in Dicer1(f/f) rather than the control was observed, reflecting the processing of pre-miRNA into mature miRNA through Dicer activity. Meanwhile, Dicer1(−/−) did not show a decrease in the luciferase activity and is in line with the absence of Dicer protein in the cell.
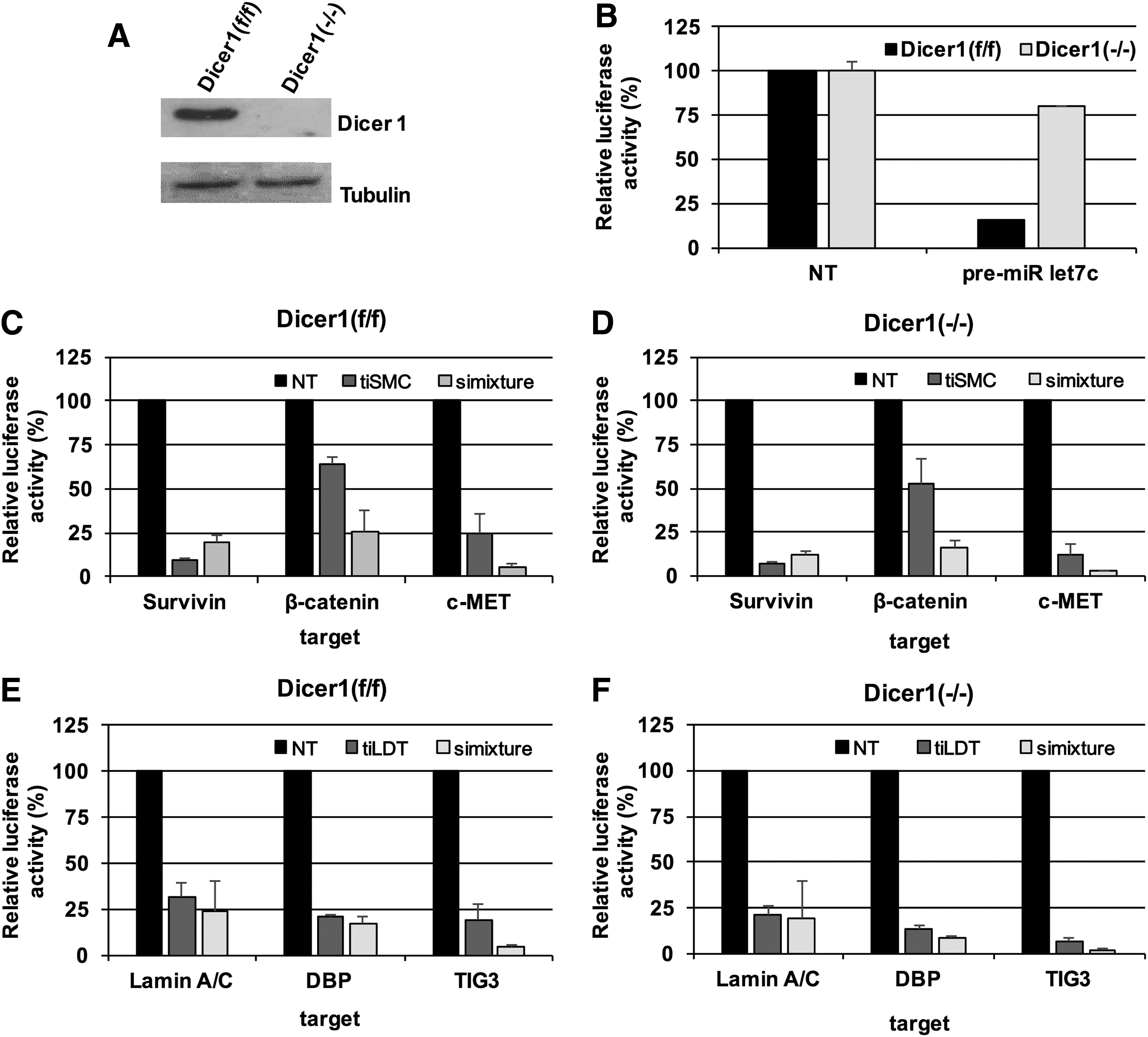
Analysis of target gene silencing of tiRNA in Dicer1knockout cell line.
Target gene silencing by tiRNA is independent of Dicer-mediated RNA processing
Next, the gene silencing activity of tiRNA in Dicer1(−/−) and Dicer1(f/f) cells was examined. tiSMC was cotransfected with reporter plasmid harboring tiSMC target sequence into Dicer1(f/f) or Dicer1(−/−) cells for 24 h, and the level of luciferase activity was analyzed through luciferase assay (Fig. 2C, D). Mixture of each siRNA for Survivin, β-catenin, and c-MET was also tested as the control. The results showed target gene silencing efficiency of tiSMC comparable with that of the siRNA mixture. When target gene silencing of tiSMC between Dicer1(f/f) and Dicer1(−/−) cells was compared, the target gene silencing activity of tiSMC was found to be maintained in Dicer1(−/−) cells. As shown in Fig. 2C and D, tiSMC showed similar level of target repression in both Dicer1(f/f) and Dicer1(−/−) cells. This result demonstrates that Dicer-mediated processing is dispensable for tiRNA-mediated gene silencing. In addition to the tiSMC, another version of tiRNA targeting Lamin A/C, DBP, and TIG3 termed as tiLDT was tested (Fig. 2E, F). tiLDT and corresponding luciferase plasmid were introduced into Dicer1(f/f) or Dicer1(−/−) cells, and level of luciferase was analyzed as already described. Similar to tiSMC, tiLDT also showed potent silencing for all the three targets in both the Dicer1(f/f) and Dicer1(−/−) cells, and the results support that multiple target gene silencing effect of tiRNA is independent of Dicer activity.
Thirty-eight nucleotide-long antisense strand of tiRNA is incorporated into Ago2
Although tiRNA showed efficient gene knockdown activity in Dicer knockout cell line, the mechanism of the 38 nt-long antisense strand of tiRNA triggers gene silencing in the absence of Dicer was still unclear. To examine whether the long antisense strand of tiRNA is incorporated into RISC, Ago2 incorporation assay was performed. Cells were transfected with siRNA or tiRNA along with FLAG-tagged Ago2-expressing plasmid, and Ago2-incorporated RNA was analyzed by Northern blot analysis (Fig. 3). In all the three tested cell lines including Dicer1(f/f), Dicer1(−/−), and HEK293T, similar pattern of the incorporation of the antisense strand of siRNA or tiRNA into Ago2 was observed. In detail, the major signal for the Ago2-associated guide strand of the tiRNA was detected at ∼38 nt in all the tested cells, suggesting that 38 nt long antisense strand of tiRNA is incorporated into Ago2 in the presence or absence of Dicer and triggers RNAi-mediated target gene silencing. Minor species ranging from ∼18 to 40 nt in length were also detected in both the input and Flag-hAGgo2 IP samples, which is most likely the degraded products during the course of experimental procedure. Higher Ago2 incorporation efficiency in HEK293T cell line than other cell lines is likely to be derived from minimum level of endogenous Ago2 expression [15] and high efficiency of transfection and protein production.
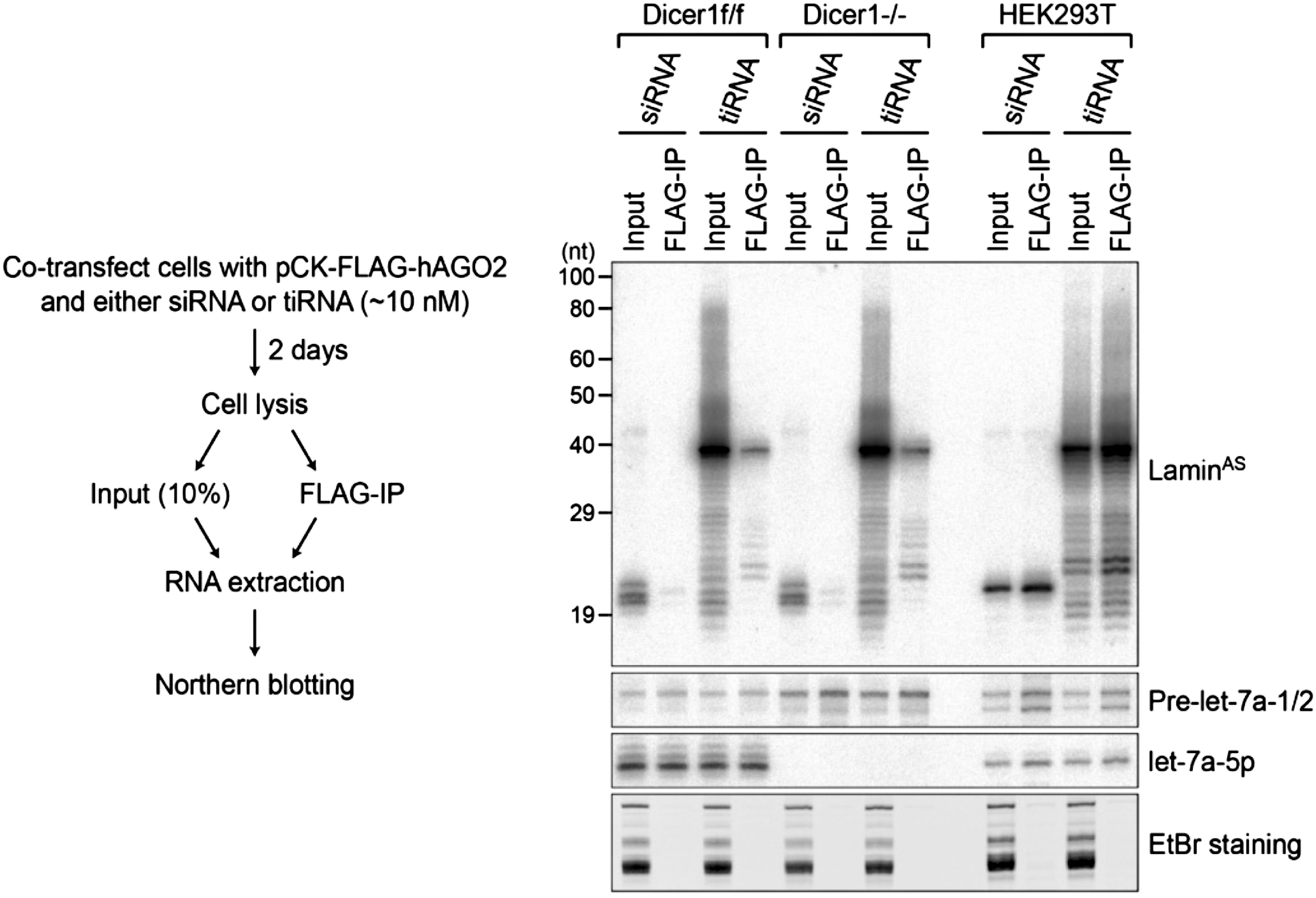
Incorporation of tiRNA antisense strand into Ago2 in Dicer1(f/f) and Dicer 1(−/−) cells. Immunoprecipitation of Flag-hAGO2 in Dicer1(f/f), Dicer1(−/−), and HEK293T cells transfected with tiRNA followed by Northern blot analysis to detect the RNA strand targeting the Lamin mRNA. Input and FLAG-IP represent input cell lysate (10%) and Flag-immunoprecipitate, respectively. Endogenous Pre-let-7a-1/2 and let-7a-5p serve as controls for Ago2 immunoprecipitation. Ago2, Argonaute 2.
Several groups including us developed diverse structural variants of RNAi-triggering molecules with improved features over the conventional siRNAs, such as reduced nonspecific effects, increased delivery efficiency, and multiple target gene silencing [4–11,16–18]. tiRNA is one of the novel RNAi triggering structures developed by our group, and we showed that tiRNA can target three different genes simultaneously. Because tiRNA has a unique tripodal RNA duplex structure, molecular mechanism underlying target gene silencing through RNAi still remains elusive. Our previous study provided an indirect evidence of Dicer-independent target gene silencing by showing potent target gene silencing by chemically modified tiRNA, which is not cleaved by Dicer in vitro [7]. In this study, Dicer dependency of tiRNA was investigated by applying Dicer knockout cell line, Dicer1(−/−). After the confirmation of the absence of Dicer protein and impaired miRNA processing in Dicer1(−/−) cell line, target knockdown activity of tiRNA was analyzed. The results using two different tiRNAs revealed that similar levels of target knockdown were achieved in Dicer1(−/−) and Dicer1(f/f) cells. In addition, we also observed direct evidence of Dicer-independent Ago2 incorporation of tiRNA strand through Ago2 incorporation assay in Dicer1(−/−) and Dicer1(f/f) cells. Altogether, we conclude that gene silencing triggered by branched tripodal RNA structure such as tiRNA is independent of Dicer-mediated processing in mammalian cells. Further studies are warranted to reveal the detailed mechanism of RISC loading and guide strand selection from the tiRNA structure.
Footnotes
Acknowledgment
This work was supported by a Global Research Laboratory grant from the Ministry of Education, Science, and Technology of Korea (no. 2008-00582). This work was also supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2017R1D1A1B03036001).
Author Disclosure Statement
S.W.H and D.-k.L. are employees and shareholders of OliX Pharmaceuticals, Inc. The other authors have no competing financial interests to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.