
Abstract
Gapmer antisense oligonucleotides (gapmers) sometimes cleave nontarget pre-mRNAs by recognizing target-like intronic/exonic portions. This off-target RNA cleavage could be a major cause of the hepatotoxicity that is induced by gapmers. In line with these findings, we hypothesized that gapmers with higher specificity have less hepatotoxicity, and that those with lower specificity have greater toxicity. To examine this concept, we investigated various Malat1-targeting gapmers with various computationally evaluated target specificities. We had expected that higher specificity gapmers would have lower hepatotoxicity, but these factors were not significantly related. In silico analysis of gapmer sequences does not always contribute to mitigating the risk of hepatotoxicity. Transcriptome analysis indicated that nontoxic gapmers do not cleave off-target RNAs, although they have many target-like RNA sequences. The present results shed light on the mechanism of the hepatotoxicity of gapmers.
Introduction
G
Other groups also reported that hepatotoxic gapmers tended to cleave off-target RNAs. They found that the expression levels of many unintended transcripts, especially long pre-mRNAs, were downregulated by administration of toxic gapmers [6]. These findings led us to consider that gapmers with higher specificity effect less hepatotoxicity, and that those with lower specificity have greater toxicity. An ultimately target-specific gapmer that does not have any presumptive target in the entire genome is expected to show no hepatotoxicity.
One of the most simple and easy way to evaluate target specificity of a gapmer is counting mismatch-containing off-targets in whole transcripts, utilizing genome database. To examine if this simple in silico approach could contribute to estimating the hepatotoxicity of gapmers, we investigated the hepatotoxicity of several locked nucleic acid (LNA)-modified, Malat1-targeting gapmers with various computationally evaluated target specificities.
First, we synthesized two gapmers: Malat1-naïve, which is a previously reported gapmer [7], and Malat1 gapmer containing one mismatch (Malat1-1MM), the presumptive off-target genes of which there were fewer than for Malat1-naïve. We administered these gapmers to mice at a high dose (≤200 mg/kg), but neither naïve nor 1MM showed significant toxicity. We could not detect significant off-target knockdown in the liver of 1MM-administered mice using microarray.
Next, we selected four novel Malat1 gapmers with low to high specificity. We expected that the gapmer with lower specificity would cause more toxicity; however, only the gapmer with the highest specificity caused hepatotoxicity in vivo. We found that one of the presumptive off-target genes was downregulated by the hepatotoxic gapmer. Thus, the computationally predicted sequence specificity of gapmers is not always related to their hepatotoxicity. The present results shed light on the mechanism of the hepatotoxicity of gapmers.
Materials and Methods
Chemical compounds
LNA-modified, fully phosphorothioated gapmers were supplied from GeneDesign (Osaka, Japan). Some gapmers were designed using siDirect (http://sidirect2.rnai.jp). Lyophilized gapmers were dissolved in saline (Otsuka Pharmaceuticals, Tokyo, Japan), and utilized for in vitro and in vivo experiments. RNAlater was purchased from QIAGEN (Hilden, Germany). All other reagents and solvents were biochemical-grade commercial products.
Cell culture and transfection
The Hepa1c1c7 murine hepatoma cell line was obtained from ATCC (Manassas, VA), and maintained at 37°C and 5% CO2 in Minimum Essential Medium Eagle-Alpha Modification (α-MEM; Invitrogen, Tokyo, Japan) that was supplemented with 10% heat-inactivated fetal bovine serum and antibiotics. Hepa1c1c7 was seeded at 2.0 × 104 cells/well in 96-well culture plates. After 24 h, gapmers were transfected using Lipofectamine3000 (Invitrogen) according to the manufacturer's instructions. Cell lysates were prepared using the CellAmp Direct RNA Prep Kit for RT-PCR (Takara Bio, Otsu, Japan) 24 h after transfection and stored at −80°C.
Animal experiment
Male C57BL6/j mice (7-weeks old; CLEA Japan, Tokyo, Japan) were purchased and maintained per the Association for Assessment and Accreditation of Laboratory Animal Care International. All experimental protocols were approved by the Institutional Animal Care and Use Committee of Shionogi Co., Ltd., and all experiments were performed in accordance with the committee's guidelines.
Gapmers, dissolved in saline, were administered subcutaneously into the back of mice at the indicated doses. Plasma was collected from the vena cava, and livers were extracted under isoflurane anesthesia at the time when the mice were sacrificed. Plasma markers for hepatotoxicity—aspartate transaminase (AST) and alanine transaminase (ALT)—were measured using the Transaminase CII-test Wako Kit (Wako Pure Chemical Industries, Osaka, Japan) according to the manufacturer's instructions for 96-well format. Approximately 40 mg of liver sample was stored in RNAlater (QIAGEN) at 4°C overnight, and then at −80°C. Total RNA was purified using the RNeasy 96 Universal Kit (QIAGEN) according to the manufacturer's instructions. In brief, liver tissue was homogenized in 750-μL Qiazol reagent. The homogenate was mixed with 150-μL chloroform, and the supernatant, containing the RNA, was transferred to an RNeasy 96-well plate. The eluted RNA solutions were stored at −80°C.
Quantitative reverse-transcription PCR
Quantitative reverse-transcription PCR (qRT-PCR) was performed with One Step SYBR primescript (Takara Bio) on an ABI7500 Fast Real-Time PCR System (Applied Biosystems, Foster City, CA) using 2 μL of cell lysate (for in vitro experiments) or 100 ng of total RNA (for in vivo experiments) as template. Quantified RNA levels were normalized to Gapdh, and are expressed relative to a saline or untreated control.
Microarray analysis and database screening
Total RNA isolated from mouse liver was analyzed using a whole-mouse genome (8 × 60K) Oligo Microarray Kit (Agilent Technology, Santa Clara, CA) according to the manufacturer's instructions. RNA transcripts containing a sequence that was similar to the target sequence of the gapmers (containing no or one mismatch) were screened using the ultrafast sequence searching database GGGenome (https://gggenome.dbcls.jp/en).
Results and Discussion
Sequence and target specificity of Malat1-naïve and -1MM
A previous report has shown that a Malat1 gapmer (Malat1-naïve) has potent knockdown effects at ≥3 mg/(kg·week) [7]. First, we focused on Malat1-naïve as a highly active gapmer model to investigate the relationship between target specificity and hepatotoxicity. Malat1-naïve possesses 5 target sequences that contain no mismatch and 89 one-mismatch-containing target sequences in the mouse genome. We redesigned Malat1-naïve by substituting a single nucleotide in the 3′ terminal region (G→T), (Malat1-1MM). Malat1-1MM showed higher specificity, with 1 presumptive target sequence with no mismatch and 76 presumptive 1-mismatch-containing target sequences in the mouse genome (Table 1).
Uppercase: LNA, lowercase: DNA. Underline indicates mismatch.
In vivo knockdown activity and hepatotoxicity
We compared the knockdown activity and hepatotoxicity of Malat1-naïve and -1MM in vivo. We had anticipated that Malat1-naïve would cause hepatotoxicity, whereas Malat1-1MM would show less hepatotoxicity than Malat1-naïve or none at all. Mice were sacrificed 7 days after gapmer injection, plasma and liver samples were collected, and knockdown activity and hepatotoxicity were examined. Both gapmers showed high knockdown activity; >95% of Malat1 expression was suppressed by only 10 mg/kg of the gapmers, and dose escalation did not increase the knockdown effect (Fig. 1A). Unexpectedly, neither of them showed hepatotoxicity at 200 mg/kg (Fig. 1B, C). This result was surprising, because all of the potent gapmers that we have ever examined were hepatotoxic at high doses (<100 mg/kg). Thus, Malat1-naïve and -1MM could be extraordinarily effective and nontoxic gapmers.

Transcriptome analysis of Malat1-1MM
The major cause of gapmer-derived hepatotoxicity is considered to be off-target knockdown. Thus, we examined whether off-target knockdown was induced by Malat1-1MM. Liver samples of the mice that received 20 mg/kg Malat1-1MM at 24 h before sacrifice were analyzed using microarray. We sacrificed mice at 24 h after gapmer administration because hepatotoxic side effect does not occur at 24 h (it appears after 48–72 h) [5]. Before the analysis, presumptive targets that contained <2 mismatches were identified using the GGGnome database. We analyzed the expression of these genes and found that eight genes were reduced by Malat1-1MM, although none reached statistical significance (Table 2); four of these genes harbored multiple target-like sequences in their pre-mRNA. No transcripts were upregulated by Malat1-1MM. Next, qRT-PCR was performed to confirm off-target knockdown in livers of mice that received 200 mg/kg of the gapmer 7 days before sacrifice. If the change of gene expressions (found by microarray analysis after 20 mg/kg administration) were caused by off-target pre-mRNA cleavage, 200 mg/kg of gapmer administration would cause much more drastic downregulation of those genes and the knockdown effect should continue until day 7. However, none of these eight genes was downregulated, indicating that off-target knockdown of these genes did not occur in the livers of mice that received even 200 mg/kg of Malat1-1MM (Fig. 1D). We cannot conclude that off-target knockdown did not occur with 200 mg/kg of Malat1-1MM, but these results imply that hepatotoxicity could not have developed without off-target RNA cleavage.
Malat1, shown in bold, is the ‘target gene’, not off-target candidate.
Knockdown activity and hepatotoxicity of the four gapmers
Next, we examined 10 newly designed gapmers to investigate the relationship between target specificity and hepatotoxicity. We examined their in vitro knockdown activity by transfecting them into the Hepa1c1c cell line (Fig. 2A). Six gapmers showed effective (>75%) knockdown activity, and we selected four of them that had varying computationally evaluated target specificities (Table 3). For example, #8 had the lowest specificity, harboring 2 target transcripts with no mismatches and 171 with target sequences containing 1 mismatch (68 of the 171 encode RNA transcripts) in the mouse genome. Surprisingly, #4 showed much higher specificity than the others, nearly targeting Malat1 exclusively; only five target sequences containing one mismatch were in the mouse genome, and only two of these five encoded RNA transcripts.
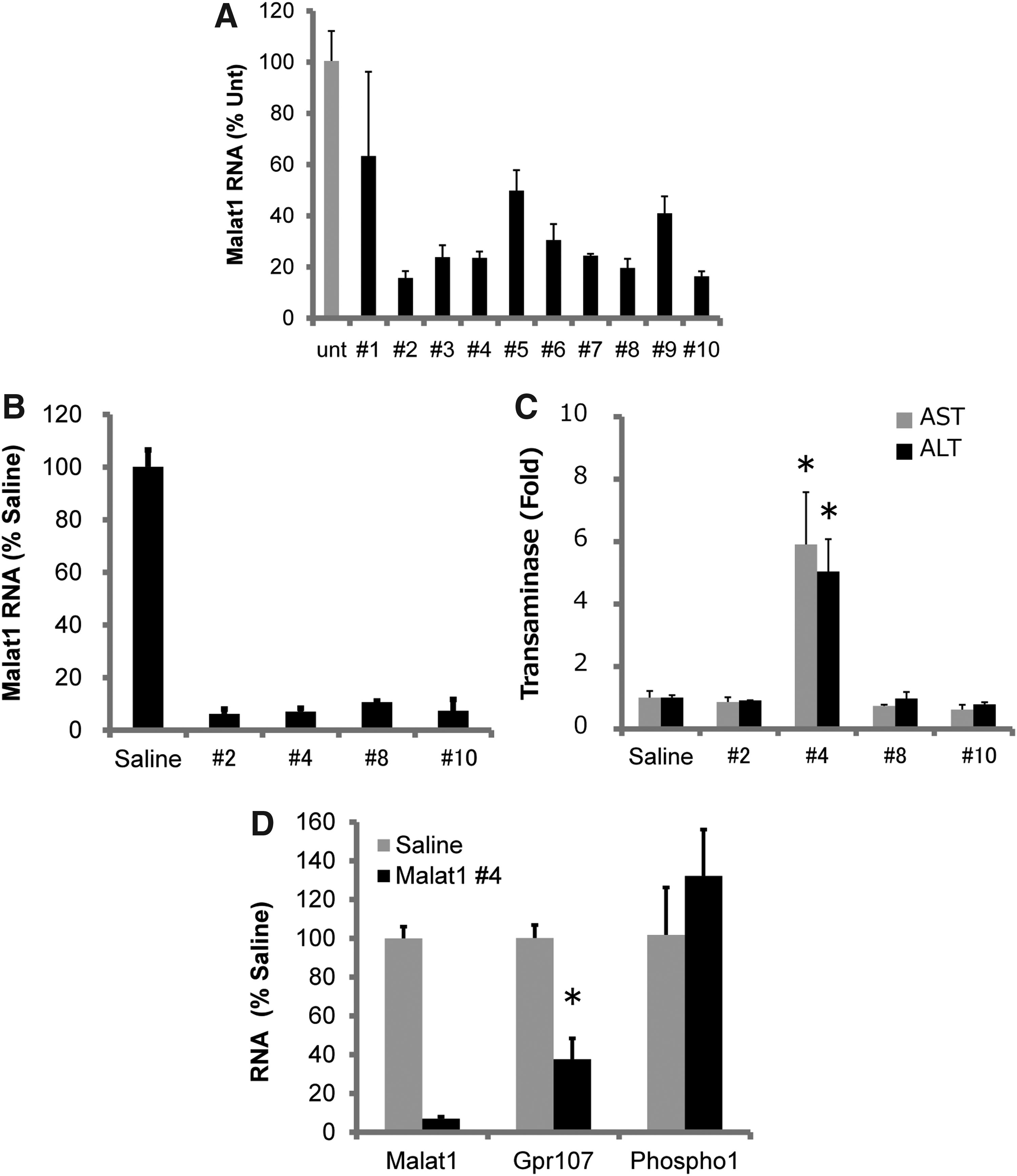
Knockdown activity and hepatotoxicity of newly selected MALAT1 gapmers.
Then, we examined the knockdown activity and hepatotoxicity of the four gapmers. The mice that received 40 mg/kg of the gapmers were sacrificed 7 days after injection, and plasma and liver samples were collected. As a result, all of the gapmers had potent (>90%) knockdown effects (Fig. 2B). Next, we analyzed ALT/AST levels in plasma and found that contrary to our expectation, only #4 showed hepatotoxicity, increasing ALT and AST levels fivefold to sixfold over the control (Fig. 2C). To investigate whether off-target knockdown had occurred, we performed qRT-PCR analysis of two presumptive targets that harbor one-mismatch-containing target sequences (Table 4). There were two candidate genes, Gpr107 and Phospho1, of which Gpr107 was significantly downregulated (Fig. 2D).
Relationship between knockdown activity and hepatotoxicity
In summary, we first examined the relationship between knockdown activity and hepatotoxicity by redesigning a previously reported Malat1 gapmer, but it showed no hepatotoxicity, even at very high doses (200 mg/kg). We performed microarray analysis but could not detect off-target knockdown, although the specificity of these gapmers was not high. Conversely, the extremely specific gapmer, #4, which possesses only 2 one-mismatch-containing RNA-coding off-target candidates, showed significant hepatotoxicity in vivo, and one of the presumptive targets was significantly downregulated. These results indicate that in silico analysis of gapmer sequences does not always select for nontoxic gapmers. The computationally evaluated target specificity of gapmers might not be related directly to hepatotoxicity or off-target knockdown.
All gapmers have the potential to cause off-target knockdown and hepatotoxicity. The most important approach for the development of gapmers as drugs is to identify those that have potent knockdown activities without causing side effects at the appropriate dose. In terms of their effect on the transcriptome, gapmers do not differ from already approved small drugs; therefore, hybridization-mediated side effects must be carefully investigated [8]. Focusing on genome-wide, computationally evaluated target specificity is considered to be an important approach for avoiding the toxicity of gapmers but is insufficient to mitigate the risk of the side effects. In silico sequence analysis is quick and simple way, but we should not rely too much on it to predict sequence specificity and hepatotoxic potential. Both experimental and computational investigations could be required to assess the side effects of gapmers [9–11]. Several effective, nontoxic gapmers have been identified, and some remain in clinical trials [7,12,13]. Furthermore, recently, novel modifications have been reported to lower the toxicity while maintaining or increasing the knockdown effect of the original gapmers [14–16]. Analysis of sequence motifs has contributed to lowering the toxicity of gapmers [17]. Comprehensive studies that focus on target specificity and other factors should be performed to select gapmer candidates for clinical trials.
Footnotes
Acknowledgments
Author Disclosure Statement
No competing financial interests exist.