
Abstract
Adjuvants are essential for efficiently inducing an antigen-specific immune response in vaccine therapy. Single-stranded RNA (ssRNA) containing guanosine- and uridine-rich sequences is recognized by Toll-like receptor (TLR)7 and/or TLR8 and induces strong immune responses; thus, the application of ssRNA as an adjuvant is desirable. The development of a ssRNA-based adjuvant, however, requires the efficient delivery of ssRNA into the endosomes of antigen-presenting cells, where the TLRs exist. To achieve this, we developed a nanostructured RNA/DNA assembly using DNA nanotechnology, which can be efficiently recognized by antigen-presenting cells. The nanostructured RNA/DNA assembly, named tetrapodRD3, was designed using a 40-mer phosphorothioate-stabilized RNA and three 40-mer phosphodiester DNAs. TetrapodRD3 was more stable than ssRNA under serum conditions. The secreted alkaline phosphatase assay using HEK-Blue hTLR cells showed that tetrapodRD3 triggered human TLR8-specific responses. Fluorescently labeled tetrapodRD3 was efficiently taken up by murine dendritic DC2.4 cells and induced a high level of tumor necrosis factor-α release from the cells. Antigen presentation by the major histocompatibility complex class I on bone marrow-derived dendritic cells was significantly increased by the addition of an antigen along with tetrapodRD3. These results indicate that tetrapodRD3 constructed using DNA nanotechnology can be a useful adjuvant targeting human TLR8.
Introduction
Vaccine therapy is useful for inducing antigen-specific immune responses against diseases such as cancer and infections. In vaccine development, adjuvants are essential to efficiently induce an antigen-specific immune response [1]. However, existing adjuvants such as alum cause adverse effects such as fever and allergic reactions [2]. Therefore, the development of new adjuvants with high efficacy is an imperative task.
Adjuvants consisting of one or several oligonucleotides have the possibility of solving the problems of the existing adjuvants. Nucleic acids and oligonucleotides are recognized by Toll-like receptors (TLRs) in the endosomes of antigen-presenting cells and induce the release of several types of cytokines [3]. There are four types of TLRs in the endosome, that is, TLR3, TLR7, TLR8, and TLR9, and each type of antigen-presenting cells has a different pattern of TLR expression [4–6]. TLR-mediated activation of antigen-presenting cells increases their antigen-presenting ability. TLR7 and TLR8 are expressed abundantly in antigen-presenting cells, especially in dendritic cells and monocytes, compared with the other two TLRs for nucleic acids. Dendritic cells and monocytes are important cells to trigger an antigen-specific immune response. Single-stranded RNA (ssRNA) containing guanosine- and uridine-rich sequences, is a typical agonist of TLR7/8 [7] and, therefore, ssRNA is expected to be a promising adjuvant.
In the development of a ssRNA-based adjuvant, it is necessary to deliver it efficiently to the endosomes of antigen-presenting cells. However, the uptake of ssRNA by antigen-presenting cells is low and ssRNA is easily hydrolyzed by RNase in biological environments [8–10]. These problems can be circumvented by complex formation with lipid nanoparticles, but the complexity of manufacturing lipid nanoparticles, and their high manufacturing costs are the major problems for their clinical applications [11]. To expand the possibility of using ssRNA-based adjuvants, development of alternative delivery approaches to lipid nanoparticles is desired.
Efficient approaches using duplex formation of nucleic acids to improve the delivery of functional nucleic acids to target cells have been reported [12–14]. One of such possible approaches is the use of DNA nanotechnology [15]. RNA-containing nucleic acid assemblies formed using DNA nanotechnology have been reported. RNA cubes developed by Afonin et al., which were assembled by six long-chain RNAs/DNAs and six siRNAs, had high structural stability in serum and high RNA interference activity [16]. However, RNA/DNA nanostructures applicable as adjuvants have not been reported so far. We developed a polypod-like DNA nanostructure (polypodna) consisting of phosphodiester oligonucleotides as a DNA nanotechnology-based delivery carrier [17]. Polypodna is formed by DNA/DNA pairing and is efficiently taken up by macrophages and dendritic cells. In addition, polypodna induced high cytokine release when it contained CpG motif, which is the TLR9 ligand. It was also shown that polypodna can integrate antisense oligonucleotides [18] and improve their retention in the systemic circulation after intravenous injection in mice [19]. Considering these findings, a polypod-like nanostructure consisting of RNA and DNA can be a useful ssRNA-based adjuvant.
It has been reported that TLR7 and TLR8 selectively recognize guanosine and uridine nucleotides. Heil et al. showed that a GU-rich sequence derived from the U5 region of HIV-1 RNA stimulated dendritic cells and macrophages to secrete several cytokines [7]. Another study showed that a 4-mer GU-rich oligoribonucleotide (ORN) induced cytokine production by binding to human TLR7/8 in dendritic cells and monocytes [20].
Therefore, this study aimed to develop a new RNA adjuvant with a polypod-like structural backbone targeting TLR7/8. To this end, we designed a tetrapod-like nanostructure consisting of a 40-mer phosphorothioate RNA and three phosphodiester DNAs, tetrapodRD3. We first evaluated the structural stability of the complex of ssRNA and tetrapodRD3 in serum. Then, we examined the recognition of tetrapodRD3 by human TLRs using TLR-expressing cells; the cellular uptake of tetrapodRD3 and its effect on tumor necrosis factor (TNF)-α release and antigen presentation were also analyzed.
Materials and Methods
Chemicals
RPMI 1640 medium and Dulbecco's modified Eagle's medium (DMEM) were purchased from Nissui Pharmaceutical Co., Ltd. (Tokyo, Japan). Opti-modified Eagle's medium (Opti-MEM) and fetal bovine serum (FBS) were obtained from Thermo Fisher Scientific, Inc. (Waltham, MA). Sodium chloride, sodium bicarbonate, potassium chloride, Tris-HCl, EDTA, ammonium chloride, and glucose were purchased from Wako Pure Chemicals Industries, Ltd. (Osaka, Japan). Poly I:C was purchased from Invitrogen (San Diego, CA). Blasticidin, zeocin, normocin, CL264, and HEK-Blue detection reagents were purchased from InvivoGen. Monothioglycerol, MEM nonessential amino acids, and the penicillin/streptomycin/glutamine-mixed solution were purchased from Nacalai Tesque, Inc. (Kyoto, Japan). The 20 bp ladder was purchased from TaKaRa Bio (Otsu, Japan). R848 and ovalbumin (OVA) were obtained from Merck KGaA (Darmstadt, Germany). DOTAP was purchased from Avanti Polar lipids, Inc. (Alabaster, AL). All other chemicals were of the highest grade available and were used without further purification.
Oligonucleotides
Phosphorothioate ORN-1 and its labeled form, with 6-FAM conjugated at the 5′-end (FAM-ORN-1), were purchased from FASMAC Corporation (Kanagawa, Japan). Phosphodiester oligodeoxynucleotide (ODN)-1, ODN-2, ODN-3, ODN-4, and CpG2006 were purchased from Integrated DNA Technologies, Inc. (Skokie, IL). ODN-1 has the same sequence as ORN-1 with thymine instead of uracil. ORN-1, ODN-2, ODN-3, and ODN-4 were designed to form tetrapodRD3, and ODN-1, ODN-2, ODN-3, and ODN-4 were designed to form tetrapodD4, or tetrapodna. The sequences of ORN-1 and the ODNs are listed in Table 1.
Sequences of Oligonucleotides Used for Preparing TetrapodRD3 and Tetrapodna
Sequences in capital letters indicate phosphorothioate modifications of the internucleotide linkages. The asterisk (*) indicates the position of the 6-FAM modification. The italic t at the 5′-terminal of ODN-2 indicates a mismatch. All ODNs are phosphodiester linkages.
ODN, oligodeoxynucleotide.
Cell culture
HEK-Blue human TLR (hTLR)3, HEK-Blue hTLR7, HEK-Blue hTLR8, and HEK-Blue hTLR9 cells were obtained from InvivoGen. These cells were cultured in DMEM supplemented with 10% heat-inactivated FBS, 4.5 g/L glucose, 0.2% sodium bicarbonate, 100 IU/mL penicillin, 100 μg/mL streptomycin, 2 mM
Preparation of tetrapodRD3
Each oligonucleotide was dissolved in Tris-EDTA (TE) buffer (10 mM Tris, 1 mM EDTA, pH 7.5). Appropriate molar ratios of the oligonucleotides involved in tetrapodRD3 were mixed with 150 mM sodium chloride; heated at 95°C for 5 min, 75°C for 2 min, and 67°C for 1 min; and then slowly cooled to 4°C. The formulation of nanostructure was confirmed by polyacrylamide gel electrophoresis (PAGE). PAGE analysis was performed with a 9% polyacrylamide gel at 200 V for 60 min. Oligonucleotides were stained with SYBR GOLD and observed using a LAS3000 imaging system (Fujifilm, Tokyo, Japan). FAM-labeled oligonucleotides was observed without SYBR GOLD staining.
Measurement of melting temperature
The melting temperature (Tm) was obtained by measuring the absorbance of tetrapodRD3 in TE buffer containing 150 mM sodium chloride at 260 nm using a Shimadzu UV-1600 PC spectrometer (Shimadzu, Kyoto, Japan) equipped with a TMSPC-8 temperature controller.
Stability of tetrapodRD3 in 10% FBS
FAM-ORN-1 or tetrapodRD3 containing FAM-ORN-1 (FAM-tetrapodRD3) were mixed with nonheat-inactivated FBS in a final concentration of 10% and incubated at 37°C. After 0, 15, 30, 45, 60, 90, or 120 min of incubation, EDTA was added to the sample solution to stop the hydrolysis. These samples were run on a 9% PAGE at 4°C. The density of the RNA bands was quantitatively evaluated using the Multi Gauge software (Fujifilm).
Secreted alkaline phosphatase assay
Four types of HEK-Blue hTLR cells were seeded on 96-well plates at a density of 5 × 104 cells per well. ORN-1, ODN-2, ODN-3, ODN-4, or tetrapodRD3 complexed to DOTAP in HEK-Blue detection solution were added to the cells to produce a final RNA concentration of 0.2 μM. After 20 h of incubation at 37°C, the optical density of the samples was measured at a wavelength of 620 nm using a microplate reader. The sampling time of 20 h was decided according to the manufacturer's instructions.
Cellular uptake of tetrapodRD3 by DC2.4 cells
DC2.4 cells were seeded on a chamber slide (WATSON Co., Ltd, Tokyo, Japan) at a density of 3 × 104 cells per well and then cultured for 24 h. The culture medium was replaced with fresh medium containing 100 nM LysoTracker Red DND-99 (Molecular Probes, Eugene, OR). The medium was removed after 1 h, then ORN-1 or tetrapodRD3 with a final RNA concentration of 0.2 μM diluted in Opti-MEM was added to the wells and incubated at 37°C for 0.5, 1, 2, 4, or 8 h. The cells were fixed with 4% paraformaldehyde for 30 min and washed twice with phosphate-buffered saline (PBS). The cells were incubated with 300 nM of 4′,6-diamidino-2-phenylindole (DAPI; Life Technologies, Eugene, OR) for 10 min at room temperature and washed twice with PBS. The slide was observed using a confocal microscope (A1R MP; Nikon Instech Co., Ltd., Tokyo, Japan).
DC2.4 cells on 96-well plate at a density of 5 × 104 cells per well were incubated with FAM-labeled ORN-1 or tetrapodRD3 diluted in 0.1 mL of Opti-MEM in a final RNA concentration of 0.2 μM for 2 h at 37°C. Cells were harvested and washed twice with PBS. The fluorescent intensity of cells was determined by flow cytometry (Beckman Courter, Indianapolis, IN) using the Kaluza software (Beckman Courter), and the mean fluorescence intensity (MFI) was calculated.
Preparation of murine bone marrow-derived dendritic cells
C57BL/6 J mice (7–9 weeks, male) were provided from Japan SLC, Inc. (Shizuoka, Japan). Protocols for all animal experiments were approved by the Animal Experimentation Committee of the Graduate School of Pharmaceutical Sciences, Kyoto University. Mice were euthanized under anesthesia using isoflurane. Femurs and tibiae were collected, and the marrow was flushed out with RPMI 1640 media with a 27G needle. The marrow suspension was filtered using 0.4 μm pore filter. After one centrifugation (430 g, 10 min), blood erythrocytes were depleted with erythrocyte lysis buffer (0.017 M Tris-HCl, pH 7.2, 0.75% ammonium chloride). Bone marrow cells were cultured in RPMI 1640 medium supplemented with 10% heat-inactivated FBS, 0.2% sodium bicarbonate, 100 IU/mL penicillin, 100 μg/mL streptomycin, 2 mM
Cytokine release from DC2.4 cells
DC2.4 cells were seeded on 96-well plates at a density of 5 × 104 cells per well and incubated for 24 h before treatment. Then, ORN-1, ODN-2, ODN-3, ODN-4, or tetrapodRD3 diluted in 0.1 mL of Opti-MEM were added to cells. The cells were incubated for 20 h, and the supernatants were collected and stored at −80°C until use. The level of TNF-α in the supernatants was determined by ELISA using OptEIA sets (BD, San Diego, CA).
Antigen presentation on BMDCs
BMDCs seeded on 96-well plates at a density of 5 × 104 cells per well were incubated with oligonucleotides (ORN-1 or tetrapodRD3) and OVA cationized by ethylenediamine, in agreement with a previous report [21], and diluted in 0.1 mL of RPMI 1640 medium for 24 h at 37°C. Cells were harvested and washed twice with PBS supplemented with 10% FBS. Cells were stained by OVA257-264 peptide bound to phycoerythrin-labeled H-2Kb antibody (Invitrogen, Carlsbad, CA) for 20 min on ice. After staining, cells were washed twice with PBS supplemented with 10% FBS, and the fluorescence intensity of the cells was determined by flow cytometry.
Statistical analysis
The data were statistically evaluated by one-way analysis of variance followed by the Tukey-Kramer test for multiple comparisons and the Student's t-test for comparisons between two groups. A P value of <0.05 was considered statistically significant.
Results
TetrapodRD3 is formed with high yield
ORN-1, ODN-2, ODN-3, and ODN-4 were used to prepare tetrapodRD3. Figure 1A shows the schematic image of tetrapodRD3 formation. The underlined part indicates ORN-1. One base pair at the 3′ end of ORN-1 and the 5′ end of ODN-2 was designed to be mismatched to accelerate the dissociation of ORN from tetrapodRD3 after cellular uptake. Figure 1B and C show the PAGE analyses of tetrapodRD3. TetrapodRD3 majorly appeared as a single band at around 400 bp, which was different from the bands of ORN-1, the mixture of ORN-1 and ODN-2, and the mixture of ORN-1, ODN-2, and ODN-4. This indicates that tetrapodRD3 was formed with high yield by the simple annealing of ORN-1 with the three types of ODNs. In addition, the band of tetrapodna consisting of ODN-1, ODN-2, ODN-3, and ODN-4 appeared at almost the same position as the band of tetrapodRD3 (Supplementary Fig. S1). This suggests that tetrapodRD3 has a similar size to tetrapodna, whose diameter was reported to be ∼7.4 nm [17]. The Tm of tetrapodRD3 was 66.5°C, which was higher than the body temperature, suggesting that tetrapodRD3 is stable under in vivo physiological conditions.
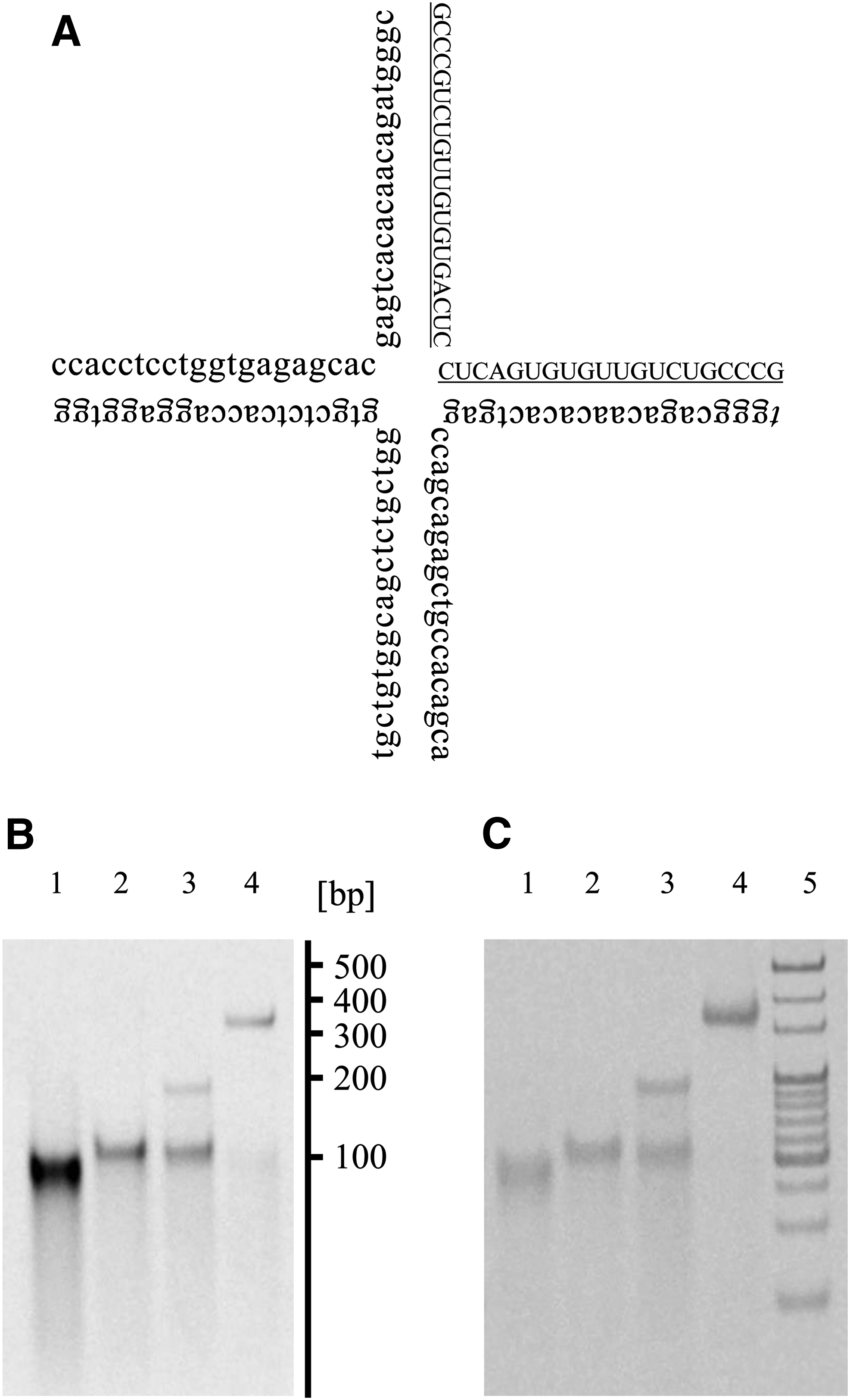
Preparation of tetrapodRD3.
TetrapodRD3 is more stable than ORN-1 in serum
Figure 2A shows the PAGE analysis of tetrapodRD3 incubated in a solution containing 10% nonheat-inactivated FBS. The bands of FAM-ORN-1 weakened with time because of the hydrolysis of ORN-1. On the other hand, the bands of FAM-tetrapodRD3 were almost unchanged for at least 2 h. A densitometric analysis of the gels indicated that the formation of a tetrapod-like structure stabilized ORN-1 (Fig. 2B).
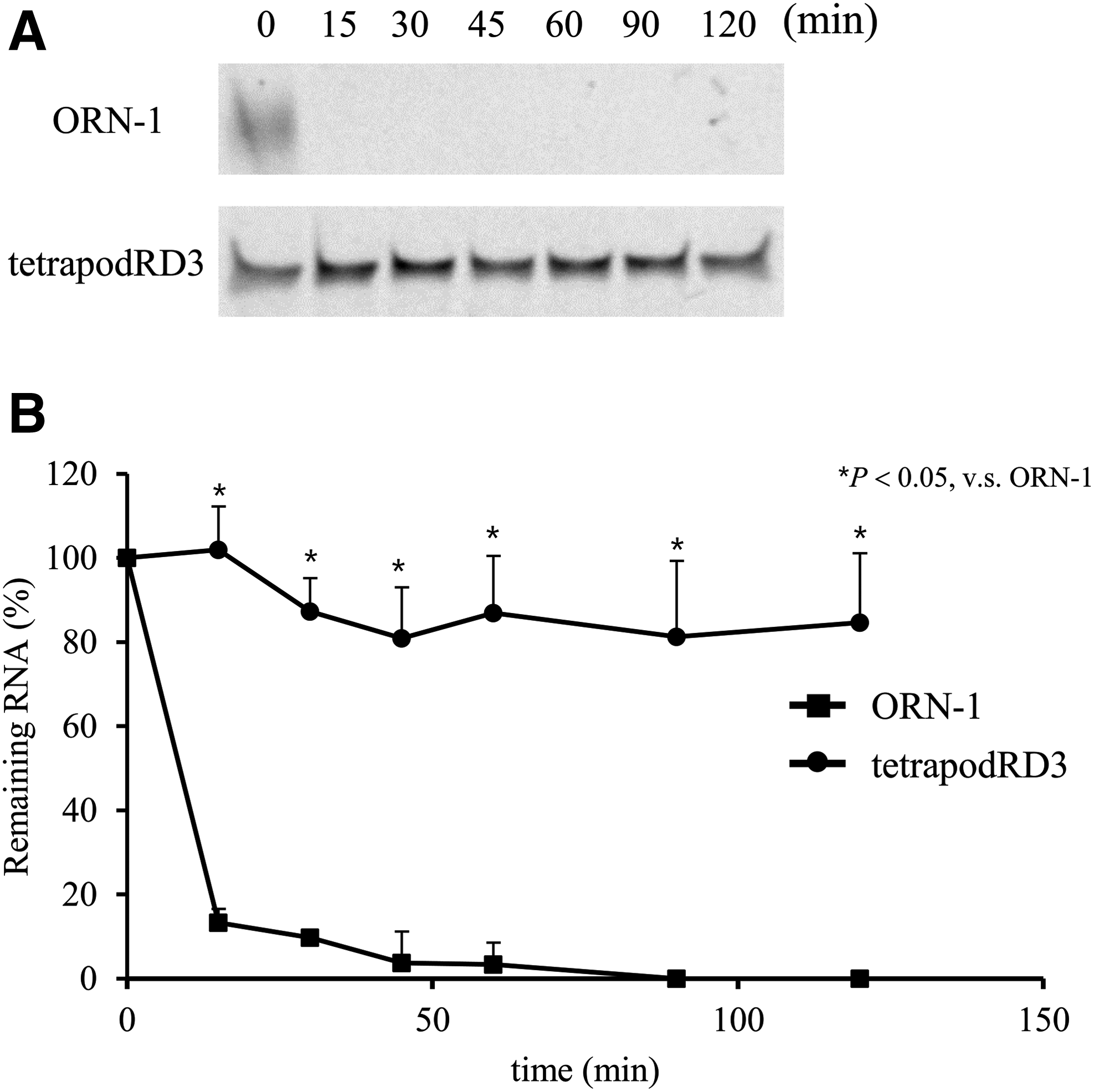
Hydrolysis of ORN-1 and tetrapodRD3 in 10% nonheat-inactivated FBS.
TetrapodRD3 triggers human TLR8-specific responses
HEK-Blue hTLR cells secrete alkaline phosphatase (SEAP) upon stimulation with TLR ligands. Figure 3 shows SEAP activity after the addition of ORN-1 or tetrapodRD3 to each type of HEK-Blue hTLR cells. The HEK-Blue hTLR8 cells released SEAP upon the addition of ORN-1 and tetrapodRD3, whereas SEAP activity observed upon the addition of tetrapodRD3 to the other types of cells was equivalent to that of the DOTAP group. Furthermore, no significant SEAP activity was detected after addition of any ODNs to HEK-Blue hTLR cells (Supplementary Fig. S2). These results suggest that ORN-1 and tetrapodRD3 are selectively recognized by hTLR8.
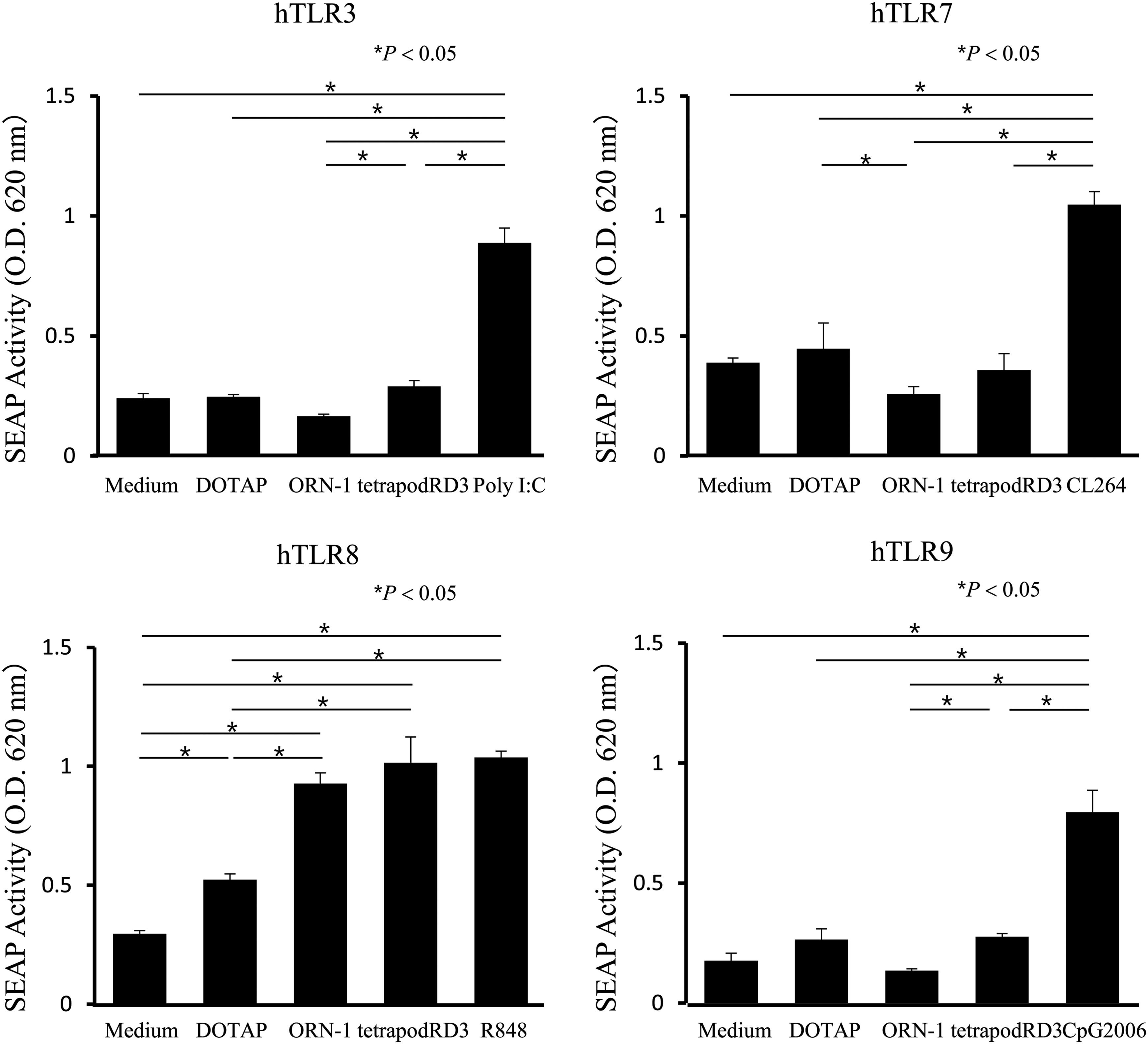
SEAP release from HEK-Blue hTLR3, hTLR7, hTLR8, and hTLR9 cells. Each TLR ligand was added to the cells at a final concentration of 1 μg/mL. The results are expressed as mean + SD of three independent experiments. Poly I:C, a TLR3 ligand; CL264, a TLR7 ligand; R848, a TLR7/8 ligand; CpG2006, a TLR9 ligand. hTLR, human Toll-like receptor; SEAP, secreted alkaline phosphatase.
TetrapodRD3 is more efficiently taken up by DC2.4 cells than ORN
Confocal microscopy images at different time points showed that FAM-terapodRD3 was taken up by the DC2.4 cells with time. In addition, the green signal of 6-FAM, which colocalized with the red signal of LysoTracker Red at 1 h or later time points (Supplementary Fig. S3), indicates that FAM-ORN-1 and FAM-tetrapodRD3 taken up by DC2.4 cells were mainly localized to the endosomes. Figure 4 shows the representative confocal microscopy images. These images showed that the cells added with FAM-tetrapodRD3 were brighter than those added with FAM-ORN-1. Figure 4B shows the MFI values of DC2.4 cells after the addition of FAM-ORN-1 samples. The MFI value of DC2.4 cells added with FAM-tetrapodRD3 was significantly higher than that added with FAM-ORN-1.
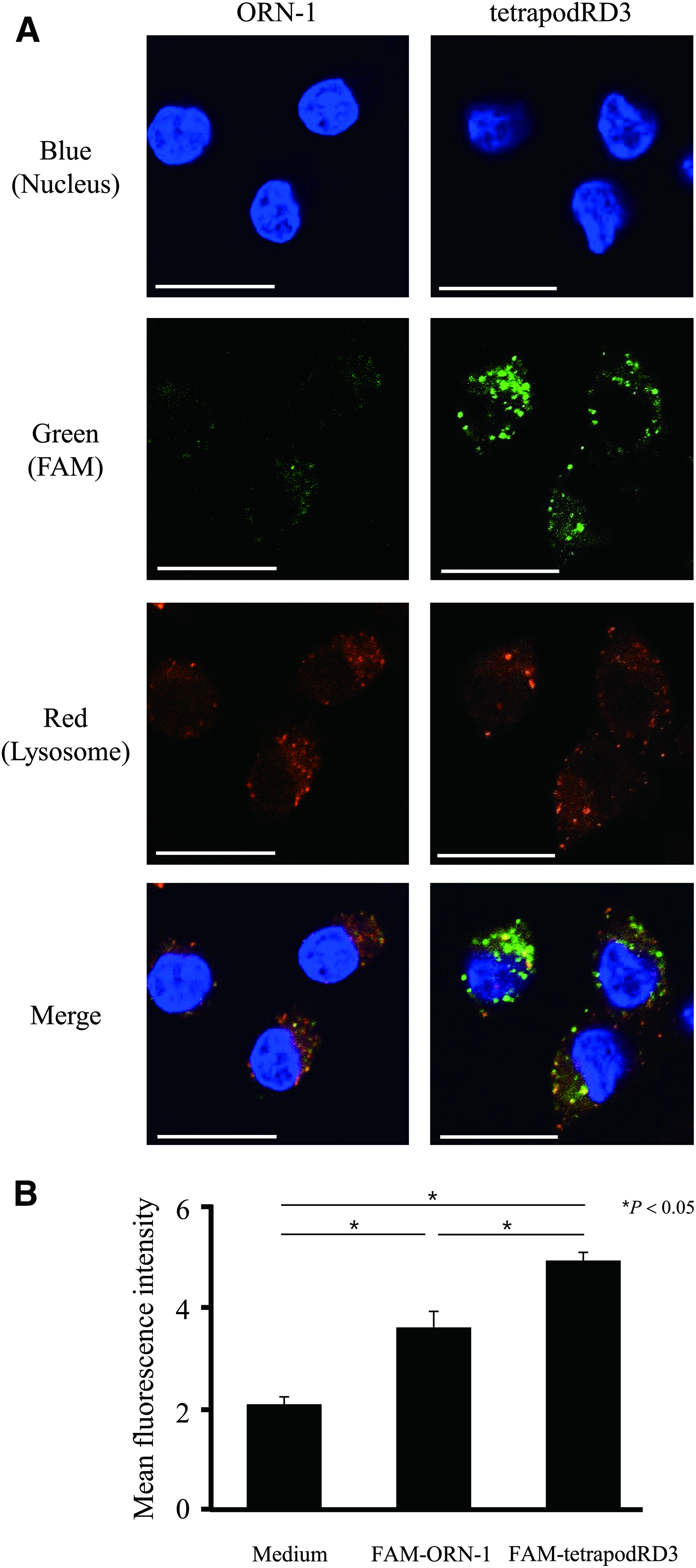
Uptake of FAM-ORN-1 and FAM-tetrapodRD3 by DC2.4 cells.
TetrapodRD3 induces a high level of TNF-α release from DC2.4 cells
Figure 5 shows the level of TNF-α release from DC2.4 cells after the addition of ORN-1 or tetrapodRD3. No significant TNF-α release was observed upon addition of ORN-1. By contrast, tetrapodRD3 induced an ∼15-times higher level of TNF-α release from DC2.4 cells than ORN-1. No significant TNF-α release was observed after addition of ODN-2, ODN-3, or ODN-4 (Supplementary Fig. S4).
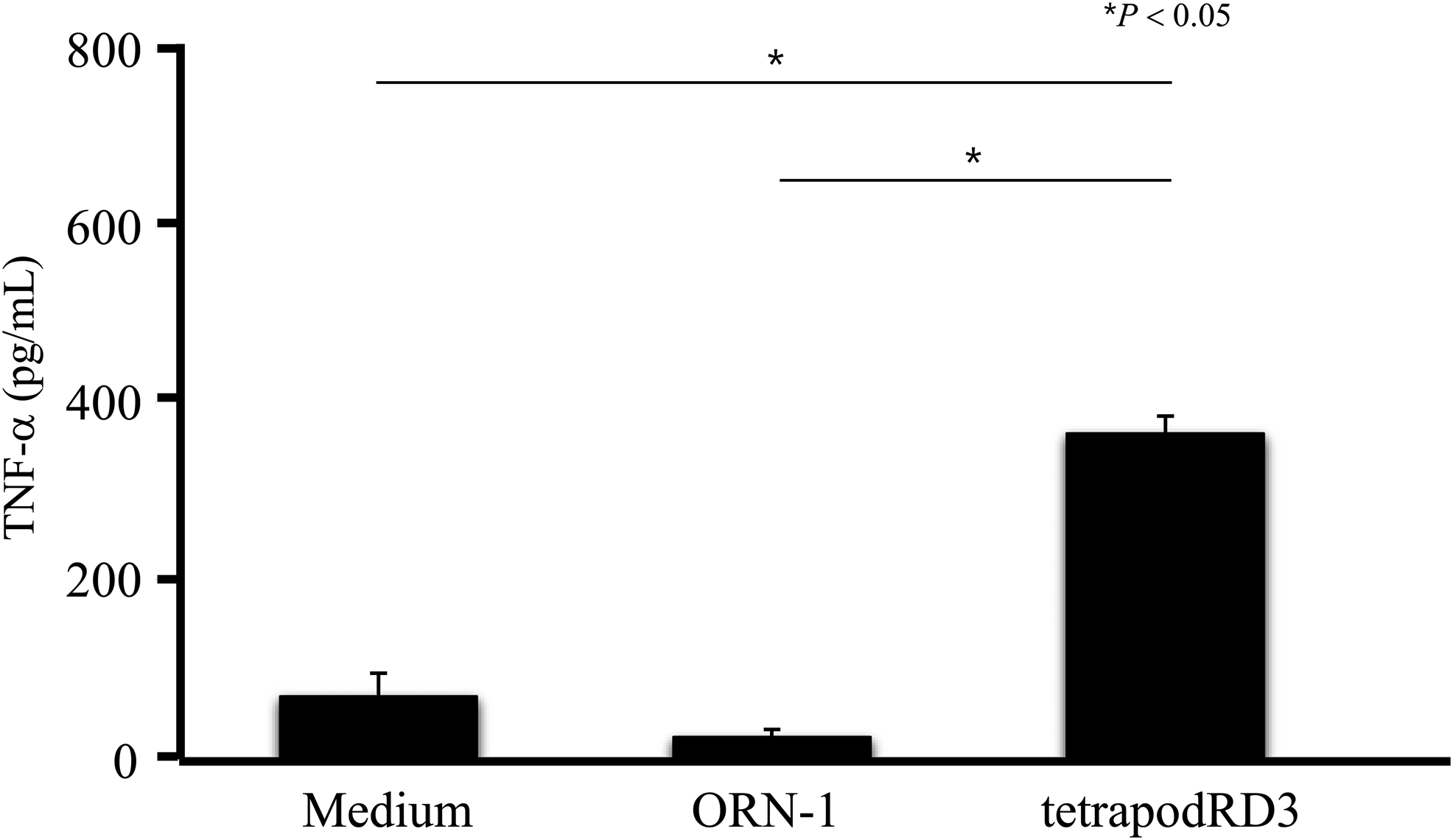
TNF-α release after addition of ORN-1 or tetrapodRD3 to DC2.4 cells. The results are expressed as mean + SD of three independent experiments. TNF, tumor necrosis factor.
TetrapodRD3 increases antigen presentation by BMDCs
Figure 6 shows detection of OVA peptide binding to major histocompatibility complex (MHC) class 1 on BMDC after the addition of RNA samples and cationized OVA. BMDC with added OVA and tetrapodRD3 showed higher peptide detection than cells with added OVA or a mixture of ORN-1 and OVA. This suggests that tetrapodRD3 increases antigen presentation by MHC class 1 on BMDC.

Antigen-presenting induction ability of ORN-1 or tetrapodRD3 in BMDCs. MFI of BMDCs upon addition of the indicated samples and staining with antibodies against the OVA peptide bound to MHC class 1. The results are expressed as mean + SD of three independent experiments. BMDC, bone marrow-derived dendritic cell; MHC, major histocompatibility complex; OVA, ovalbumin.
Discussion
Several approaches such as the use of lipid nanoparticles can efficiently deliver ssRNA to antigen-presenting cells [8–10]. However, such approaches are complex and expensive to manufacture. Therefore, the development of a simple and versatile delivery approach is in high demand. Recently, DNA nanotechnology has been attracting attention as a new delivery approach for biomolecules through the development of DNA nanostructures. We previously reported that polypod-like DNA nanostructures increased the cellular uptake and immunostimulatory activities of CpG DNA [17]. The present study further shows that this DNA nanotechnology-based strategy can be applied to RNA delivery. We revealed that targeted delivery of ssRNA to antigen-presenting cells by polypod-like nanostructured RNA can be a useful approach to increasing the therapeutic potential of RNA.
PAGE analyses suggested that tetrapodRD3 is in a similar size to tetrapodna. It has been verified by atomic force microscopy that tetrapodna was formed in a tetrapod-like structure as designed [17]. Therefore, it can be speculated that tetrapodRD3 could also be in a tetrapodna-like structure.
It was shown that nanostructured RNA was markedly stable in serum compared with ssRNA. TetrapodRD3 consisted of RNA/DNA and DNA/DNA duplexes. Duplex formation of RNA with DNA is predicted to reduce the hydrolysis by RNase, compared with ssRNA and RNA/RNA duplex. Furthermore, it is expected that the complex structural properties of tetrapodRD3 contribute to avoid the access of RNase to the RNA and prevent its hydrolysis. We have reported that the tetrapod-like DNA nanostructure was not significantly hydrolyzed in 50% nonheat-inactivated FBS for 2 h [22], indicating that the nanostructure formation increased the resistance against DNase degradation.
This study suggests that tetrapodRD3 is recognized specifically by human TLR8. The SEAP activity of each HEK-Blue hTLR cell type induced by tetrapodRD3 showed a similar trend to ssRNA-induced SEAP activity. Therefore, similarly to ssRNA, structured RNA is also recognized specifically by hTLR8. There is a species difference in the recognition of ssRNA by TLR7/8. In humans, ssRNA is preferentially recognized by TLR8, whereas in mice it is preferentially recognized by TLR7. Therefore, tetrapodRD3 might also be preferentially recognized by TLR7 in mice. It is reported that the activation of hTLR8 by hTLR8 ligands, such as resiquimod, leads to the release of several types of cytokines, such as TNF-α, IL-6, and IL-12 [23,24]. This hTLR8-mediated activation reduces virus proliferation and tumor size. These reports suggest the usefulness of tetrapodRD3 as an adjuvant targeting hTLR8.
There are several RNA receptors in antigen-presenting cells. It is known that retinoic acid-inducible gene-I (RIG-I) is an RNA receptor in the cytoplasm of antigen-presenting cells [2,25]. RIG-I recognizes 5′-triphosphate ssRNA with a short double-stranded copyback section and 5′-triphosphate dsRNA [26]. Therefore, we assume that the ssRNA used in this study might not be recognized by RIG-I as it does not contain the mentioned features. The delivery to RIG-I is dependent on the mechanism of cellular uptake. We reported that polypod-like nanostructures are taken up, at least partially, through macrophage scavenger receptor-1 (MSR1) [27]. It has been reported that ligand binding to MSR1 induces clathrin-mediated endocytosis, and that the ligands are then sorted to the endosomes [28]. Furthermore, our results suggest that tetrapodRD3 is localized in the endosomes after being taken up by DC2.4 cells. These results suggest that tetrapodRD3 avoids direct migration to the cytoplasm, where RIG-I-mediated recognition takes place.
To achieve delivery to the endosomes of antigen-presenting cells, ssRNA has to be efficiently taken up by the cells. TetrapodRD3 has a three-dimensional structure, and its physicochemical properties are different from those of ssRNA. These differences could explain the high cellular uptake of tetrapodRD3. In addition, tetrapodRD3 was more stable compared with ssRNA under the presence of nucleases, thus it is expected that upon addition of tetrapodRD3, the hydrolysis of ssRNA in the medium is avoided before its delivery to the endosomes. TNF-α release from dendritic cells after the addition of RNA showed a similar trend to their uptake of RNA. Our results proved the hypothesis that increased cellular uptake of tetrapodRD3 increased the recognition of ssRNA by TLR8 and cytokine release.
This study suggests that tetrapodRD3 increases the antigen-presenting ability of dendritic cells. In general, an efficient stimulation of antigen-presenting cells leads to efficient antigen presentation by MHC class 1. Therefore, the elevated antigen presentation by dendritic cells upon addition of tetrapodRD3 might be due to the efficient stimulation of the cells through increased delivery of ssRNA by the nanostructured RNA/DNA assembly.
We have previously reported that polypodna containing CpG DNA induced significant cytokine production, not only in vitro, but also in vivo [29]. Based on the results obtained from the present study, polypod-like nanostructure formation of RNA is considered to be useful for enhancing the in vivo immunostimulatory activity of RNA. In the future, we aim to evaluate the application of tetrapodRD3 in vivo, and its therapeutic effects in various diseases, such as cancers and viral infections.
In conclusion, the nanostructured RNA/DNA assembly, tetrapodRD3, developed in this study showed high biological stability, efficient uptake by antigen-presenting cells, high immunostimulatory activity, and high antigen presentation. Altogether, these results indicate that tetrapodRD3 can be a useful adjuvant for the antigen targeting of TLR7/8.
Footnotes
Acknowledgments
This work was supported by the Japan Science and Technology Agency (JST) Core Research for Evolutional Science and Technology (CREST) [grant no. JPMJCR1521]; by the Japan Society for the Promotion of Science (JSPS) (grant no. 26293008); and the Uehara Memorial Foundation.
Author Disclosure Statement
The authors declare no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.