
Abstract
Amyloid-β (Aβ) plaque deposits in the brain are considered to be one of the main pathological markers of Alzheimer's disease (AD). The sequential proteolytic cleavage of amyloid precursor protein (APP) by the aspartyl proteases β-site APP-cleaving enzyme 1 (BACE1) and γ-secretase produces Aβ. Therefore, BACE1 inhibition is a very attractive target for the treatment of AD. Our previous work identified a DNA aptamer named A1 that can bind to BACE1 with high affinity and specificity and exhibits a distinct inhibitory effect on BACE1 activity in an AD cell model. The purpose of this research was to test the effect of aptamer A1 in Tg6799 mice. Four-month-old Tg6799 mice were randomly divided into two groups and treated with aptamer A1 and ineffective aptamer A1scr, respectively, by intracerebroventricular injection. Subsequent behavioral experiments showed that treatment with the aptamer A1 improved the cognitive abilities of the AD mice. Western blot indicated that BACE1 and soluble amyloid precursor protein β (sAPPβ) expression significantly decreased in the A1-treated mice. Moreover, aptamer A1 reduced the content of Aβ42 and the number and density of senile plaques in AD mice. Therefore, our results indicate that aptamer A1 is a novel specific and potent BACE1 inhibitor and is a promising potential target for the treatment of AD.
Introduction
Alzheimer's disease (AD) is acknowledged as the most common progressive and irreversible degenerative disease of the central nervous system and is the leading cause of dementia in the elderly. According to the World Alzheimer Report 2018, there are 50 million people worldwide living with dementia, and this number will more than triple to 152 million by 2050 [1]. Currently, the treatment of AD is limited, and there is no cure for the disease. Therefore, new drugs for the treatment of AD are urgently needed to delay the progress or improve the symptoms of AD.
Extracellular amyloid plaque deposits in the brain are considered to be one of the main pathological markers of AD. The amyloid-β (Aβ) peptide, a component of amyloid plaques, is produced by the sequential cleavage of amyloid precursor protein (APP) using the aspartyl proteases β-site APP-cleaving enzyme 1 (BACE1) and γ-secretase. The cleavage of APP by BACE1 produces amino-terminal soluble amyloid precursor protein β (sAPPβ) and carboxy-terminal protein C99. C99 is further cleaved by γ-secretase to produce Aβ peptides of different lengths.
BACE1 is considered to be an effective target for the development of AD drugs. Numerous studies have shown a significant increase in BACE1 activity in the plasma [2], brain tissue [3–6], and cerebrospinal fluid [7–9] of subjects with mild cognitive impairment and probable AD. The targeted deletion of the BACE1 gene in mice abolishes Aβ production, reduces the number of amyloid plaques, and prevents Aβ-dependent memory deficits [10,11]. The downregulation of BACE1 gene expression can reduce Aβ neuropathology and rescue memory deficits in mouse models of AD [12,13]. Furthermore, BACE1 inhibitors efficiently lower Aβ levels in the brains of primates and other animals [14–16]. In addition, several BACE1 inhibitors have successfully progressed into clinical trials [17], but these trials were terminated, due to off-target safety concerns or a lack of potency. Therefore, there are currently no highly effective BACE1 inhibitors that can be used clinically.
Aptamers are single-stranded DNA/RNA oligonucleotides screened by the Systematic Evolution of Ligands by EXponential Enrichment (SELEX) technique [18]. As “chemical antibodies,” due to their folded three-dimensional conformations, aptamers are capable of binding a variety of targets, including proteins, peptides, metal ions, small molecules, or even intact cells, with high affinity and specificity. Compared to conventional antibodies, aptamers have further advantages, such as a small size, controllable and cost-effective synthesis, facile chemical modification, good stability, low toxicity, low immunogenicity, and rapid tissue penetration. Therefore, aptamers have been widely used in therapy, diagnostics, and imaging applications [19–21]. Based on our previous in vitro studies on BACE1-specific DNA aptamers, we found that A1 has inhibitory effects on BACE1 activity [22], showing preliminarily that A1 can serve as a potential BACE1 inhibitor. This finding needs further confirmation by in vivo experiments in an animal model of AD, and our previous work has proven that the Tg6799 mouse line is an effective AD animal model [23]. Therefore, the purpose of this research was to test the effect of aptamer A1 in Tg6799 mice.
Materials and Methods
Materials
Animals
We used Tg6799 mice (No. 008730; Jackson Laboratories, Bar Harbor, ME), which coexpress human APP and PS1 transgenes containing five FAD mutations on the C57/B6XSJL background, in our experiments. The transgenic mice were identified by polymerase chain reaction. All animals were housed in standard laboratory cages (three to five per cage) under a 12-h light/12-h dark cycle (lights on at 8:00 A.M.) in a temperature-controlled room (21°C–25°C). The mice had free access to water and food. Only male mice were used in the experiments. Behavioral testing was performed during the light cycle between 12:00 and 17:00. All procedures were conducted in accordance with the Chinese Council on Animal Care Guidelines and were approved by the Southern Medical University Animal Ethics Committee. Efforts were made to minimize animal suffering and reduce the number of animals used.
Aptamer synthesis
Aptamer A1 and A1scr were synthesized by Invitrogen (Carlsbad, CA). The aptamer A1 is an aptamer for the BACE1 protein. Aptamer A1: 5′-GCAATGGTACGGTACTTCCGTCATCAGCTTGTGATGTGGATGCGAACTGCAAAAGTGCACGCTACTTTGCTAA-3′. Aptamer A1scr: 5′-TAGTTTGTCAGGGAAGACATTCCTGTGACCTCCCATGATACGAGTGGGTGCAGGGTACCGAACATATTTATCC-3′. Indirect enzyme-linked immunosorbent assay (ELISA) was used to verify the binding affinity between aptamer A1 or A1scr and protein BACE1. And our previous results indicated that A1 can specifically bind to BACE1 with high affinity, whereas unrelated aptamer did not have high affinity to BACE1 [22]. Thus the random ineffective sequence aptamer A1scr was used as a negative control of the A1 aptamer. Before drug administration, the aptamers were diluted to 100 μM in 0.9% saline solution.
Methods
Implantation of the cannula system and the experimental schedule
The implantation of the cannula system (Shenzhen Ruiwode Life Technology Co., Ltd.) was performed in 4-month-old male Tg6799 mice as described by Huang et al. [24] with few modifications. Briefly, the male mice were deeply anesthetized by an intraperitoneal (i.p.) injection of 4% chloral hydrate (0.2 mL per 20 g). Then, the surgical area (0.5 cm around bregma) was exposed after skin preparation. The mice were fixed to an adapter of a stereotaxic instrument, and the skull was kept parallel with the table top. A cannula drug delivery system was implanted in the right cerebral ventricle of the mice using the following plane skull coordinates; 0.6 mm posterior to bregma, 1.1 mm lateral to the midline, and 2.1 mm ventral to the surface of the skull (Fig. 1b). After implantation, the cannula system was secured with dental cement. The cap of the cannula was inserted into the cannula to prevent occlusion and infection. Trypan blue dye was injected, and the diffusion of the dye throughout ventricle was detected to ensure accurate positioning of the implanted cannula. The mice were housed two per cage in compartments separated with a clapboard, and the treatments were administered by a four-channel drug pump beginning 3–4 days after surgery. The experimental schedule is shown in Fig. 1a. The Tg6799 transgenic mice were randomly divided into two groups. The mice in the experimental group were treated with aptamer A1 and the control group with the ineffective aptamer A1scr by intracerebroventricular injection. The injections were given a total of 15 times at a frequency of once every 3 days and a dose of 0.4 nmol per injection.
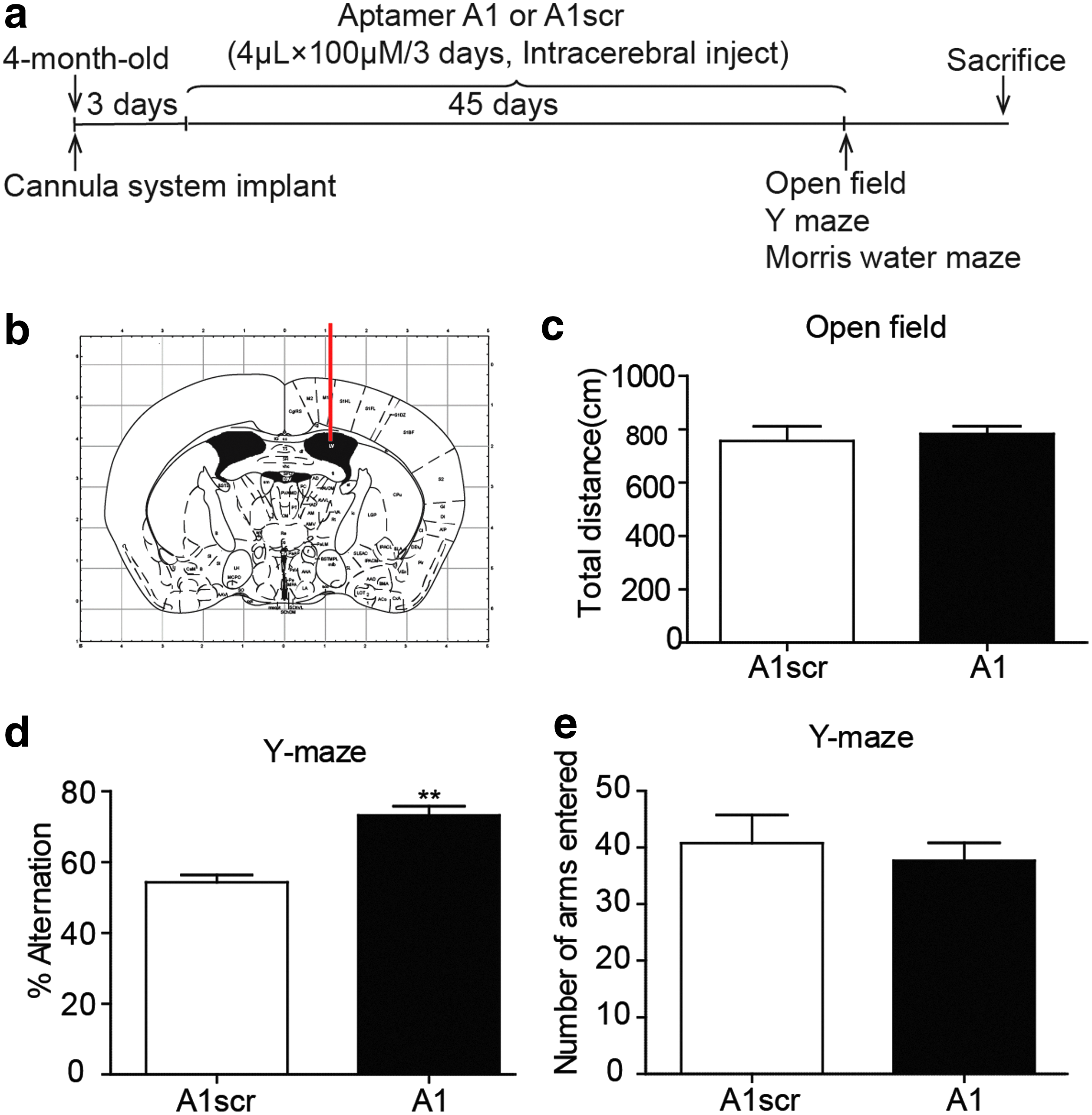
The BACE1-specific DNA aptamer A1 improves working memory in Tg6799 mice.
Open field test
The open field test (OFT) is a widely used procedure for examining locomotor and anxiety-like behavior. The open-field apparatus was a rectangular chamber (60 × 60 × 40 cm) composed of gray polyvinyl chloride. The center area was illuminated using 25-W halogen bulbs (200 cm above the field). The mice were gently placed into the center of the testing chamber for a 5-min recording period and monitored using an automated video tracking system. The digitized image of the path taken in those 5 min was automatically analyzed using the DigBehv animal behavior analysis program.
Spontaneous alternation Y-maze test
The Y-maze consisted of three 35 × 5 × 10 cm black arms and was used to assess short-term spatial working memory in the mice. The test was performed as published before [25,26]. Before the initial use and after each trial, the Y-maze was thoroughly cleaned with a paper towel and 70% ethanol and allowed to dry. The arms of the maze were clearly designated as “1,” “2,” and “3” (Supplementary Fig. S1). Each mouse was placed at the same end of one arm and allowed to freely explore the maze during an 8-min session. The entire process was video recorded. The sequence of the arms entered and the total number of arms entered were recorded. A mouse with good spatial working memory will remember the arms of the maze that it has already explored and show a tendency to enter an unvisited arm. Spontaneous alternation was defined as a mouse entering a different arm of the maze in each of three consecutive arm entries, for example, 123, 231, or 312 but not 121. The spontaneous alternation percentage was calculated by the following formula [25]:
Morris water maze
Morris water maze is a procedure for assessing spatial and related forms of learning and memory. A pool (diameter 1.2 m) and platform (10 cm) were utilized for the water maze studies. The temperature of the pool was maintained at 22°C, and nontoxic white paint was added to the water. The pool was surrounded by a white circular curtain that held three visual cues. The platform was submerged 1 cm beneath the surface of the water. The pool was divided into eight parts. Except for the four points adjacent to and opposite to the platform, the remaining points around the edge of the pool were served as the starting positions. The mice were trained for 6 days with four trials a day and allowed a maximum of 90 s per trial to locate the hidden platform. If the mice failed to find the platform within 90 s, the animal was manually guided to the platform. On the 7th day, the platform was removed, and the mice were placed opposite the position of the platform and scored for the time spent in the target quadrant and the number of platform crossings within 60 s.
Western blot analysis
The tissue was harvested and lysed in ice-cold RIPA lysis buffer (P89901; Thermo Fisher Scientific, Waltham, MA) containing 1 × protease inhibitors (P1862209; Thermo Fisher Scientific). After calculating the protein concentration using a BCA Assay Kit (23227; Thermo Fisher Scientific), 20 μg of total protein was separated using 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a PVDF membrane (Millipore, Bedford, MA). The membrane was blocked with 5% defatted milk powder at room temperature (RT) for 1.5 h, immunoblotted with a primary antibody at 4°C overnight, and subsequently incubated with an HRP-conjugated secondary antibody (ZSGB-BIO, Beijing, China) at RT for 1 h. The primary antibodies that were used are as follows: anti-BACE1 (1:1,000; Abcam, Cambridge, MA), anti-sAPPβ (1:500; IBL, Fujioka, Japan), and anti-GAPDH (1:5,000; Sigma-Aldrich, St. Louis, MO). The blots were visualized using a super-enhanced chemiluminescence (ECL) system (Bio-Rad), and the protein expression levels were evaluated by quantifying the gray density of the western blot bands using Image Lab software. The expression of each target protein was normalized to the expression of the internal control (GAPDH).
Quantitation of Aβ40 and Aβ42
To detect Aβ40 and Aβ42 by ELISA, brain tissue was homogenized as described in the user guide of the ELISA Kit (Invitrogen). Briefly, 8 volumes of cold 5 M guanidine-HCl in 50 mM Tris were added to the brain tissue, and the brain tissue was homogenized thoroughly with a mechanical homogenizer. The homogenate was mixed on a shaker at RT for 3 h, and the sample was diluted 10 fold with cold phosphate buffered saline (PBS) containing 1 × protease inhibitor cocktail and centrifuged at 16,000 g for 20 min at 4°C. The supernatant was carefully transferred to a new tube and diluted with standard diluent buffer. The assay was performed by adding 50 μL of detection antibody to 50 μL of each standard and sample and incubating with shaking for 3 h at RT, aspirating all of the liquid and washing 4 times, incubating the samples with 100 μL of the HRP anti-rabbit antibody for 30 min at RT, aspirating all of the liquid and washing four times, incubating the samples with 100 μL of stabilized chromogen for 30 min at RT, adding 100 μL of the stop solution, and reading the absorbance of each well at 450 nm. Curve fitting software was used to generate the standard curve, and the concentrations of the unknown samples were read from the standard curve.
Thioflavin S staining
After behavioral tests, mice were deeply anesthetized with 4% chloral hydrate (100 g/mL, i.p.), intracardial perfused with 0.9% saline solution, and subsequently 4% paraformaldehyde. The brain tissues were postfixed overnight in 4% paraformaldehyde and immersed in 30% sucrose for 48 h for cryoprotection. And then the tissues were cut at 35 μm using a cryostat (Leica CM1950; Leica Biosystems, Wetzlar, Germany). For thioflavin S staining, brain slices were allowed to defrost at RT and were then washed with PBS for 5 min. Later, the sections were stained in freshly prepared 0.4% thioflavin S solution for 5 min in the dark, washed twice for 3 min with 70% alcohol, washed twice for 5 min with PBS, and mounted. Image acquisition was performed under a fluorescence microscope.
Statistical analysis
Statistical analysis was conducted using GraphPad Prism version 6.0 (GraphPad Software). All data are expressed as the mean ± standard error of mean. “n” refers to the number of mice per group. Student's t-test was used to compare the data from two groups. The statistical significance level for all experiments was set at *P < 0.05, **P < 0.01.
Results
Chronic DNA aptamer A1 administration rescues behavioral deficits in Tg6799 mice
To test whether the BACE1-specific DNA aptamer affected animal behavior and to explore the potential side effects of A1, the mice were subjected to a series of behavioral tests. In the OFT, the total distance between the A1scr-treated group and A1-treated group (Fig. 1c) is similar (A1: n = 9; A1scr: n = 9), which showed that the side effects of A1 were limited. Furthermore, we tested the spontaneous alternation of the mice in the Y-maze test to assess spatial working memory. The results showed that the A1-treated group had an elevated mean alternation percentage compared to that of the A1scr-treated group (Fig. 1d, e), which showed that the BACE1-specific DNA aptamer improved working memory in the Tg6799 mice.
Furthermore, we performed the Morris water maze test to explore whether A1 can improve spatial learning and memory in AD mice. The AD mice treated with A1 exhibited a similar swimming velocity as that of the A1scr-treated group (Fig. 2b). During the training phase, the latency to find the hidden platform of the mice treated with A1 was shorter compared with the A1scr-treated mice (Fig. 2a), suggesting that the learning ability of the mice was improved. Similarly, during the probe test, compared to the A1scr-treated group, the experimental mice crossed over the absent platform more times and spent more time in the target quadrant (Fig. 2c–f). Together, these results suggest that the BACE1-specific DNA aptamer improved spatial learning and memory in the AD mice.

The BACE1-specific DNA aptamer A1 rescues impaired water maze learning in Tg6799 mice.
Aptamer A1 decreases the protein expression of BACE1 and sAPPβ in the hippocampus of Tg6799 mice
To test whether aptamer A1 can reduce the protein expression of BACE1 and sAPPβ in vivo, we conducted western blotting for BACE1 and sAPPβ. sAPPβ is produced through the cleavage of APP by BACE1. The western blot results showed that the protein expression levels of BACE1 and sAPPβ in the aptamer A1-treated group were significantly lower than those in the control group (Fig. 3a, b), which showed the inhibitory effects of A1 on BACE1 activity in vivo.
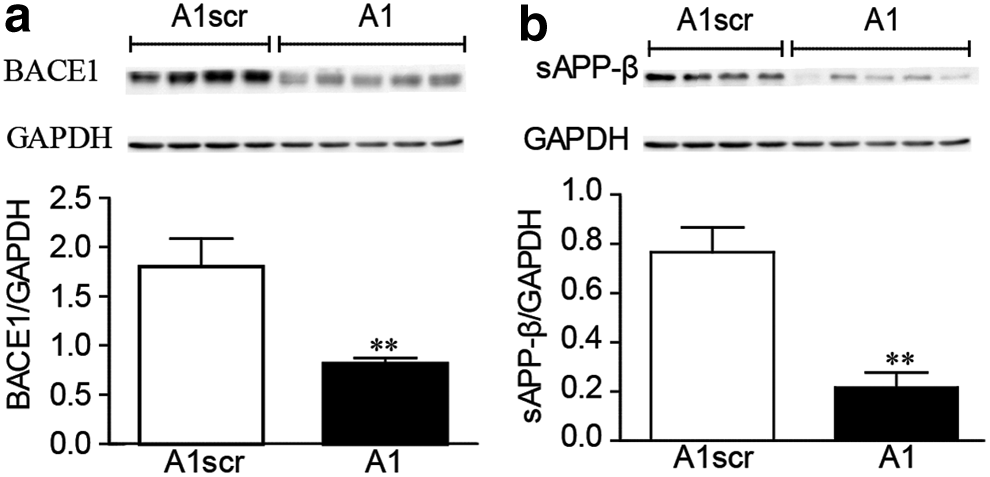
Chronic intracerebroventricular injections of aptamer A1 inhibit the protein expression of BACE1 and sAPPβ. Western blot detection of the protein expression levels of
Aptamer A1 inhibits Aβ deposition in Tg6799 mice
BACE1 is considered the rate-limiting enzyme in the production of Aβ. Because the protein expression levels of BACE1 and sAPPβ in the A1-treated group were significantly lower than those in the control group, we speculated that Aβ expression was also decreased. To test whether A1 can reduce the expression of Aβ and the number of amyloid plaques, we conducted ELISA and thioflavin S staining. The ELISA results showed that aptamer A1 dramatically reduced the levels of Aβ42 (Fig. 4b) but not significantly affected the levels of Aβ40 (Fig. 4a) in the hippocampus of the mice. Meanwhile, the results of plaque staining with thioflavin S showed that the number and density of senile plaques in the brains of the aptamer A1-treated group were lower than those in the control group (Fig. 4c). These results showed that aptamer A1 diminished Aβ42 levels and amyloid plaques in the Tg6799 mice.

Chronic intracerebroventricular injections of aptamer A1 inhibit Aβ deposition. The quantification of total
Discussion
The causal role of Aβ in the pathogenesis of AD has been supported by much genetic, clinical, and cell biological evidence [27]. Meanwhile, BACE1 activity is known to be upregulated upon aging in the brains of animals and humans [28–30]. Our previous research has shown that amyloid plaque deposits can be markedly observed in 3-month-old Tg6799 mice, and we found that memory impairment occurs in 4-month-old Tg6799 mice, unlike in wild-type mice [23]. Thus, multiple efforts have been made to develop various types of BACE1 inhibitors as treatments to rescue recognition impairment in AD in both animals and patients. Although several BACE1 inhibitors have successfully progressed into clinical trials, there are currently no highly effective BACE1 inhibitors that can be used clinically [31].
Previous work by our groups has demonstrated that the inhibition of BACE1 with aptamer A1 decreases the expression of Aβ in an AD cell model [22]. The current study demonstrates that chronic intracerebroventricular injections of aptamer A1 can effectively rescue Aβ pathology and memory deficits in Tg6799 mice. The side effects of aptamer A1 in the OFT were limited. Therefore, the in vitro and in vivo results to date show that aptamer A1 is a potent BACE1 inhibitor.
To date, there are intense efforts to develop two types of BACE1 inhibitors, peptidomimetic molecules and nonpeptidic molecules [32]. Unfortunately, the peptide-based BACE1 inhibitors possess ineffective in vivo pharmacological properties, such as low oral bioavailability, long serum half-life, and restricted brain exposure due to low blood–brain barrier (BBB) penetration. However, nonpeptidic BACE1 inhibitors also have their limitations; they are both too small to bind with sufficient affinity to the enzymatic active site and too large to exhibit satisfactory pharmacokinetics and suitable brain penetration.
The intrinsic characteristics of aptamers, such as a small size and high affinity binding to targets, make them a great option as therapeutic agents for the inhibition of BACE1. The use of aptamers in therapy, diagnostics, and imaging applications, both alone and coupled with a variety of agents, has made great progress in the past 29 years [33–35]. There are currently 11 aptamers that can be used to treat various diseases, including macular degeneration, cancer, thrombotic diseases, and inflammation, in different stages of clinical trials [36]. Furthermore, aptamers that target brain diseases for diagnosis and treatment are underdeveloped [37–39]. Decreased Aβ42 production and sAPPβ levels following A1 treatment in Tg6799 mice illustrate the impressive effects of A1 on BACE1 activity and validate BACE1 inhibition as a disease-modifying and symptomatic therapy for AD.
In this study, the administration of aptamer A1 was carried out through repeated intracerebroventricular injections—an effective method of administration for studying brain diseases [40], which inevitably damaged the brain tissue due to the chronic administration. Moreover, the degradation of the unmodified nucleic acids [41,42], as well as the short half-life of aptamers and their poor ability to penetrate the BBB [18], should be considered when applying aptamers in vivo. Therefore, polyethylene glycol (PEG) modifications and combining aptamers with cell-penetrating peptides and drug delivery carriers such as liposomes [43,44], poly(lactic-co-glycolic acid) (PLGA) nanoparticles [45], and gold nanoparticles can enhance the half-life and BBB penetration of aptamers [46]. Furthermore, the underlying cause of AD remains elusive, and the amyloid hypothesis is not sufficient to explain the intricate multifactorial pathogenesis of AD. Therefore, a clear diagnosis in the early stages of AD, early disease intervention, and the development drugs that can synergistically inhibit BACE1 and modulate a second interrelated AD target may be a feasible solution. Thus, we will next consider using two aptamers that bind different targets of AD to enhance the effectiveness of the treatment.
In conclusion, the BACE1-specific DNA aptamer A1 was observed to exhibit symptomatic modification in a mouse model of AD. Our findings revealed that aptamer A1 is an effective BACE1 inhibitor that, with modification, has the potential to be further used in clinical trials.
Footnotes
Author Disclosure Statement
The authors declare no competing interests.
Funding Information
This project was supported by the National Natural Science Foundation of China (nos. 81471388, 81771484, 81871418), the Natural Science Foundation of Guangdong Province, China (no. 2018A030313835), the Program for Changjiang Scholars and Innovative Research Team in University (no. IRT_16R37), and Guangzhou Science and Technology Project (no.201707020027).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.