
Abstract
Diabesity is a growing pandemic with substantial health and financial consequences. We are developing microRNA (miRNA)-based drug candidates that transform fat storing adipocytes into fat burning adipocytes (browning effect) to treat metabolic diseases characterized by lipotoxicity. Through phenotypic screening in primary cultures of human subcutaneous adipocytes, we discovered that inhibition of miRNA-22-3p by several complementary antagomirs resulted in increased lipid oxidation, mitochondrial activity, and energy expenditure (EE). These effects may be mediated through activation of target genes like KDM3A, KDM6B, PPARA, PPARGC1B, and SIRT1 involved in lipid catabolism, thermogenesis, and glucose homeostasis. In the model of Diet-Induced Obesity in mice of various ages, weekly subcutaneous injections of various miRNA-22-3p antagomirs produced a significant fat mass reduction, but no change of appetite or body temperature. Insulin sensitivity, as well as circulating glucose and cholesterol levels, was also improved. These original findings suggest that miRNA-22-3p inhibition could become a potent treatment of human obesity and type 2 diabetes mellitus, the so-called diabesity characterized by lipotoxicity and insulin resistance.
Introduction
Human obesity is a worldwide pandemic due to sedentary lifestyle and excessive consumption of energy-dense foods rich in saturated lipids and carbohydrates [1]. Current medical treatments of obesity have poor benefit-to-risk profiles, fail to reach their long-term goals, and do not meet patients' expectations [2,3]. Therefore, there is a need for a convenient, effective, and safe medical treatment of obesity and related metabolic disorders with lipotoxicity that should significantly reduce health care costs and improve and save human lives [4].
Mammalian adipocytes are categorized into white adipocytes (WAT) storing energy in the form of lipids and brown adipocytes (BAT) rich in mitochondria and expending energy through uncoupled fatty oxidation. It is now well established that subcutaneous WAT can evolve into calorie-burning “beige” or “brite” adipocytes [5–7]. Thus, inducing “browning” of subcutaneous WAT is a promising strategy for treating human obesity and related cardiometabolic disorders [8–13].
Mitochondrial uncoupling protein 1 (UCP1) is responsible for thermogenesis and heat dissipation by uncoupling oxidative phosphorylation from adenosine triphosphate (ATP) synthesis [14–17]. The promoter region of the UCP1 gene contains many distinct regulatory sites modulating its transcription [18]. Furthermore, the UCP1 gene promoter region is heavily methylated and its demethylation increases UCP1 expression [19]. Zhang and colleagues have shown that the human lysine (K)-specific demethylase 3A (KDM3A/JHDM2A/JMJD1A) regulates the expression of metabolic genes and obesity resistance [20,21]. Its loss of function results in obesity and hyperlipidemia in mice. KDM3A regulates beta-adrenergic effects on metabolism and body weight control by modulating thermogenesis in brown adipose tissue [22]. Similarly, miR-199a-3p modulates the adipogenesis of bone marrow-derived mesenchymal stem cells by targeting KDM6A/WNT signaling [23]. Inhibition of Lysine-specific demethylase-1 (LSD1, KDM1A) reduced glucose uptake and glycolytic activity, with a concurrent activation of mitochondrial respiration in human hepatocellular carcinoma cells [24]. As reviewed recently by Inagaki et al., several histone methyl-modifying enzymes such as EHMT1, JMJD1A, JMJD3, and LSD1 are involved in the beige adipose cell fate determination. They suggested that KDM3A/JMJD1A is a potential therapeutic target for the treatment of obesity-related diseases, including metabolic syndrome and type 2 diabetes [25,26].
The microRNAs (miRNAs) play important roles in the pathogenesis of many diseases, including cancers, atherosclerosis, cardiovascular, and metabolic disorders [27,28]. miRNAs are attractive drug candidates because the simultaneous modulation of many target genes by a single miRNA may provide effective therapies of complex diseases like obesity and diabetes mellitus [29].
Our working hypothesis is that the degree of methylation of the promoter regions of genes like UCP1 and PPARA involved in metabolism and thermogenesis is under the control of demethylases like KDM3A. Removal of miRNA inhibition of these demethylases' expression should result in meaningful changes in metabolism and thermogenesis.
Materials and Methods
In silico experiments
Candidate miRNAs and their target genes were screened, analyzed, and selected from public and commercial databases (eg, TargetScan, BioGPS, Geneious), using literature reports and in silico modeling results. Selection was based on the known and/or predicted interactions of miRNAs with selected target genes.
In vitro experiments
Human subcutaneous preadipocytes purchased from ZenBio (NC; Superlots pooled from several donors, catalog no.: SP-F-SL) were plated at confluence in 96-well plates and allowed to attach overnight in preadipocyte medium (Dulbecco's modified Eagle's medium [DMEM]/Ham's F-12 [1:1, v/v], 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid buffer, fetal bovine serum, and antibiotics) following the supplier's instructions. The next day, the medium was removed and replaced with differentiation medium for 2 days (DMEM/Ham's F-12 (1:1, v/v), 100 μM ascorbic acid, 0.85 μM insulin, 20 nM sodium selenite, 0.2 nM triiodothyronine, 1 μM dexamethasone, 100 μM isobutyl-methylxanthine, 100 nM rosiglitazone, and antibiotics). On day 3, the cells were transfected with miRNA analogs (50 nM final concentration, each condition in sixplicate) using DharmaFECT 1 and allowed to incubate for 3 days. Thereafter, cells were cultured in maintenance medium (DMEM/Ham's F-12 [1:1, v/v], 100 μM ascorbic acid, 0.85 μM insulin, 20 nM sodium selenite, 0.2 nM triiodothyronine, and antibiotics). Positive controls included 100 nM rosiglitazone throughout the cultures. After 3 days, medium was exchanged every other day with fresh maintenance medium until day 14 postdifferentiation. On day 14, the cells were gently washed. Using a 96-well format, gene expression was determined by qRT-PCR Kit following the manufacturer's instructions (TaqMan Gene Expression Cells-to-Ct kit; Life Technologies; cat no. AM1728) and a 7900 HT Fast Real Time PCR System (Applied Biosystems). Control genes were TBP, GAPDH, LRP10, and UCP3. Using a six-well format, mRNA profiling was performed by Next Generation Sequencing (Illumina HiSeq 2500).
Mitochondrial mass and activity, as well as UCP-1 protein expression, were determined by high content imaging (ArrayScan XTI and BMG FLUOstar Galaxy reader), with UCP-1 antibody (abcam; cat no. ab10983) and MitoTracker
To characterize the effects of miR-22-3p inhibition on energy utilization in human subcutaneous adipocytes in culture, oxygen consumption rate (OCR) was measured with the XF24 Analyzer (Seahorse Bioscience) using the XF Palmitate-BSA Fatty Acid Oxidation Substrate Kit during the successive addition of oligomycin (to assess ATP production and proton leak), carbonyl cyanide-p-trifluoromethoxyphenylhydrazone (FCCP) (to assess maximum respiration), and antimycin/rotenone (to assess reserve capacity and nonmitochondrial respiration), following the manufacturer's instructions. Human subcutaneous preadipocytes (ZenBio) were seeded into 96-well plates and differentiated for 7 days, then placed in maintenance medium according to the manufacturer's instructions. On day 14, the adipocytes were transfected with 50 nM of the miR-22-3p antagomir APT-110. Medium was exchanged every 2–3 days, and Seahorse assay was performed on day 21. Negative control was bovine serum albumin (BSA), and positive control was 100 nM rosiglitazone throughout the culture.
To test the ability of miR-22-3p antagomirs to de-repress luciferase expression in cells cotransfected with a miR-22-expression plasmid and a miR-22–3p luciferase reporter gene, HeLa cells were transfected at a 3:1 ratio of miR-22 expression plasmid to miR-22-3p luciferase reporter plasmid. Untransfected cells served as a control for background luminometer readings. As “No drug” neutral control, wells received phosphate-buffered saline (PBS) alone, and as a positive control, cells were only transfected with miR-22-3p luciferase reporter plasmid alone. On day 0, the cells were seeded at 10,000 cells/well into 96-well plates, and control or test compounds were added at a final concentration of 4, 20, and 100 nM in triplicate wells. On day 1, luciferase activity was read on luminometer.
In vivo experiments
To assess the metabolic effects of miR-22 inhibition in animal models of obesity, we treated C57BL/6J male mice of various ages with miR-22 antagomirs for up to 8 weeks [30].
After acclimation for 1–2 weeks, C57BL/6J male mice (Jackson Laboratory, CA) were randomized according to their body weight into treatment groups (12 animals per group, 3 animals of the same group per cage). The animals were randomized to a normal 10% fat chow diet (Research Diet; D12450B) or to a 60% high fat diet (Research Diet; D12492). The mice were administered single subcutaneous injections of saline or a miR-22 antagomir (15 mg/kg) in the left inguinal fat pad (injections on days 0, 2, and 4 of week 1, then once a week for up to 8 weeks) while they remained on normal chow or the 60% high fat diet. The mice were housed in positively ventilated polysulfonate cages with high-efficiency particulate air filtered air. The cages were changed every 2 weeks. The animal room was lighted entirely with artificial fluorescent lighting on controlled 12-h light/12-h dark cycle (6 a.m.–6 p.m. light). The temperature and relative humidity ranges in the animal rooms were maintained at 22°C ± 4°C and 50% ± 15%, respectively. The animal room was set for 15 air exchanges per hour. Filtered tap water acidified to a pH of 2.5–3.0 was provided ad libitum. Mice were monitored on a daily basis. Body weights were measured weekly. Body weight changes were calculated for each mouse by subtracting the body weight on the first day of dosing (baseline) from the body weight on the subsequent day of dosing and calculating the percent change from baseline body weight. Average percent body weight change ± standard error of the mean (SEM) was then calculated by treatment group. Food consumption per cage was measured twice weekly at 3- and 4-day intervals. Rectal temperature was measured with a Physitemp, BAT-10 Thermometer. Mice underwent body composition analysis by nuclear magnetic resonance (NMR) (Bruker LF50 BCA-Analyzer) while they were gently restrained. Blood samples were collected by retro-orbital bleeds or cut tip of the tail for measurements of glucose, insulin, cholesterol, and liver function tests. At the end of the study, liver, heart, inguinal fat, perirenal and/or epididymal fat, and subscapular fat were collected, weighed, then frozen for future gene expression analysis or placed in 10% neutral buffered formalin solution (Sigma-Aldrich; cat no. HT501128), then washed in PBS, pH 7.4, and transferred to 70% ethanol for subsequent processing for histologic and immunohistochemistry analyses. Spleens were weighed and discarded. Blood was collected and processed into serum or plasma aliquots at the time of necropsy by cardiocentesis. Serum levels of liver enzymes, glucose, insulin, cholesterol, and triglycerides were measured.
miR-22 antagomirs
miR-22-3p antagomirs (and control miRNAs) were purchased from Dharmacon GE (CO; miRIDIAN microRNA Hairpin Inhibitors), Exiqon (MA; LNA™ microRNA Inhibitors) or designed by AptamiR Therapeutics, Inc., and custom synthesized (US Patent 62/329,537 on “Inhibition of mir-22 miRNA by APT-110” initially published on 02 November 2017, WO2017/187426 A1).
Statistical analysis
Results given in the text and data points in the figures are shown as the mean ± SEM. Statistical analysis used analysis of variance (ANOVA) and Student's t-test, unless nonparametric test was selected, based on data distribution (GraphPad Prism 8).
Study approval
All animal studies were performed according to Institutional Animal Care and Use Committee (IACUC)-approved protocols and in compliance with the Guide for the Care and Use of Laboratory Animals (National Research Council, 2011) in Office of Laboratory Animal Welfare (OLAW)-assured and Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC)-accredited facilities (The Jackson Laboratory or the University of Texas at Austin, TX).
Results
Structure of the human UCP1 gene
The 5′UTR region of the human UCP1 gene contains many distinct regulatory sites, allowing a wide range of factors to modulate its transcription (Fig. 1). In addition, the promoter region of the UCP1 gene is heavily methylated. Methylation of CpG islands within gene promoters usually leads to their silencing. Conversely, the CpG islands of promoters are usually unmethylated when genes are expressed. For instance, UCP1 expression involves demethylation of CpG dinucleotides found in regulatory cyclic AMP response elements (CREs) in the UCP1 enhancer region, as well as modification of histone tails [19].
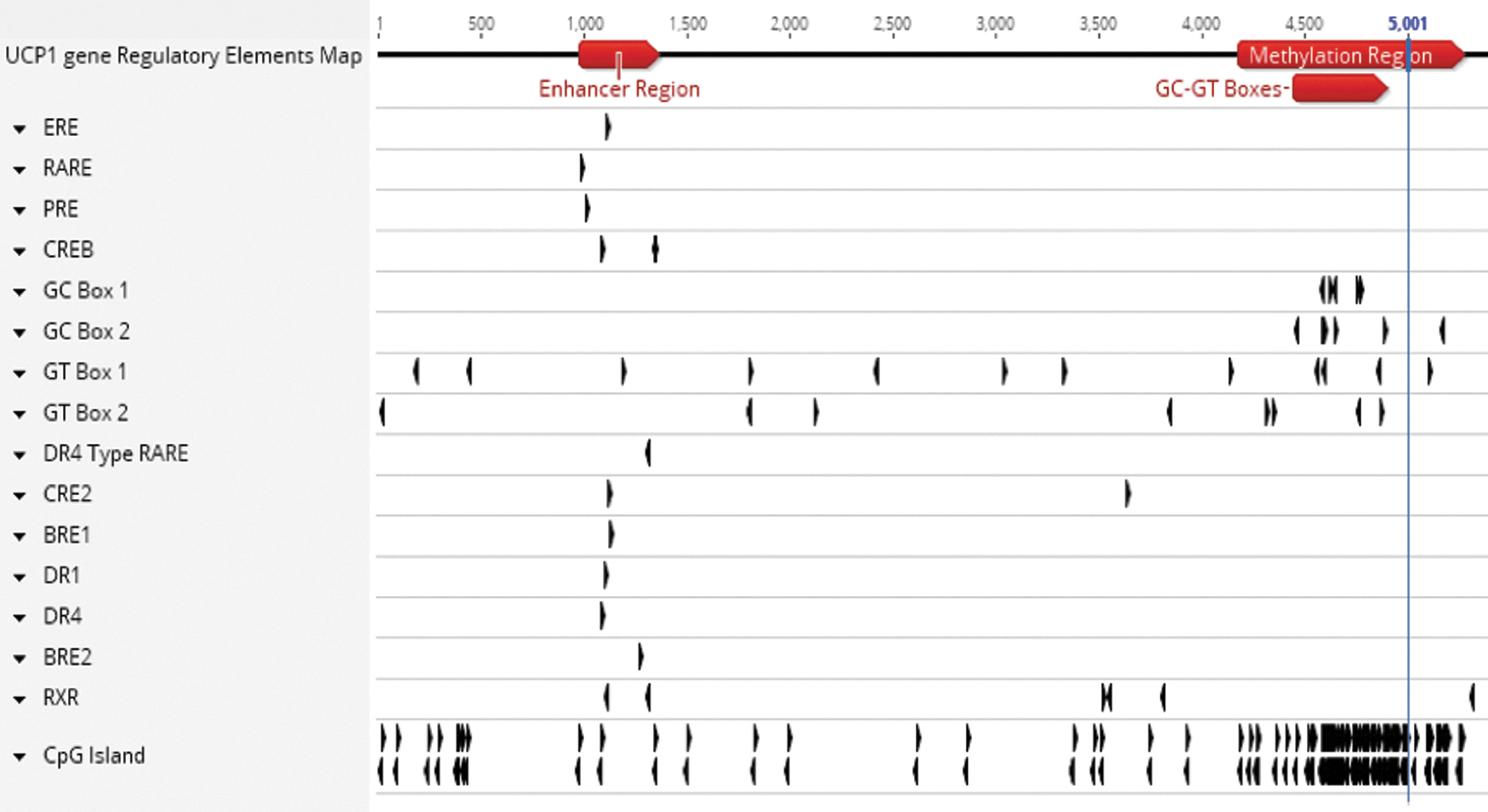
Structure of the human UCP1 gene 5′UTR region. A map of the 5′UTR region of the human UCP1 gene (Entrez gene ID: 7350) was generated with the molecular biology analysis program Geneious®. This region contains many response elements, including a 350-base pair “Enhancer Region” and a heavily methylated (“CpG islands”) promoter region (the transcription start site is located at position 5,001). BRE, brown fat regulatory element; CRE, cyclic AMP response element; CREB, cyclic AMP response element-binding protein; DR-1, peroxisome proliferator-activated receptor (PPAR)/RXR binding element; DR-4, liver X receptor (LXR) binding element; ERE, estrogen response element; PRE, progesterone response element; RARE, retinoic acid response element; RXR, retinoid X receptor; UCP1, uncoupling protein 1.
In silico identification of miRNAs binding to KDM3A and related genes
We found that the human KDM3A 3′UTR region is a conserved target (Fig. 2) for hsa-miR-22-3p, a unique, conserved, and universal miRNA that is highly expressed in human and murine subcutaneous adipocytes and is transported by adipocyte exosomes [31–34]. Similarly, the human lysine (K)-specific demethylase 6B (KDM6B) 3′UTR region is a conserved target for hsa-miR-22-3p. Therefore, we inferred that an increased production of the demethylases KDM3A and KDM6B induced by a miR-22-3p antagomir could lead to demethylation of the UCP1 gene promoter region, thus facilitating binding of several regulatory elements to increase UCP1 production. Furthermore, we also noticed that the human PPARA, PPARGC1B, and SIRT1 3′UTR regions are conserved targets for miR-22-3p (Fig. 2), suggesting that miR-22 also modulates the peroxisome proliferator-activated receptor (PPAR) pathway, PPARGC1B, and SIRT1 that are involved in glucose/lipid metabolism and thermogenesis [35–39].
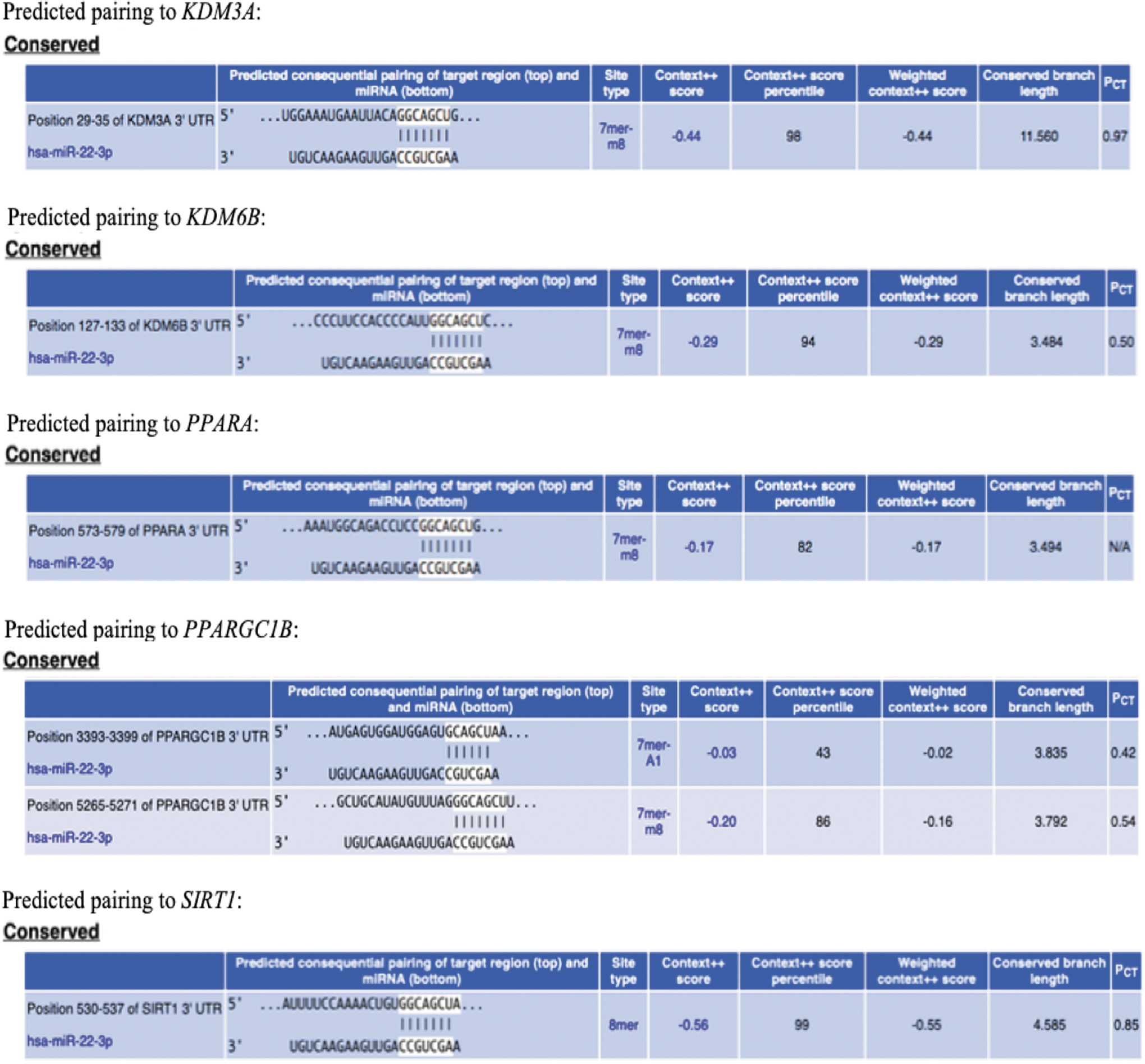
Prediction of hsa-miR-22-3p conserved pairing to the 3′UTR region of KDM3A and other metabolic/thermogenic genes (TargetScanHuman 7.2 prediction of miRNA target database). miRNA, microRNA.
In silico search of miR-22-3p gene targets
We curated a list of 721 genes involved in lipid metabolism, oxidative phosphorylation, mitochondrial functions, respiratory cycle, browning of adipocytes, and thermogenesis (Supplementary Table S1). We then utilized several miRNA Targets and Expression in silico tools and our own proprietary in silico meta tool (R-AptamiR) to identify 542 putative mRNA targets of miR-22-3p. These 542 putative mRNA targets were matched against our AptamiR curated list of 721 target genes, and we found that 60 putative target genes of miR-22-3p belong to our curated list (Supplementary Table S2).
In vitro testing of miR-22-3p inhibition
In vitro studies of miR-22-3p inhibition were carried out in primary cultures of human subcutaneous adipocytes, the ultimate targets for an antiobesity drug (“clinical trial in a dish”) (Supplementary Fig. S1).
Gene profiling of human adipocytes in culture during miR-22 inhibition
A preliminary experiment suggested that a human miR-22-3p antagomir (Dharmacon GE) induced several fold increases of UCP1 (41-fold) and UCP2 (61-fold) gene expression, whereas the expression of UCP3 was not altered (1.45-fold), as assessed at day 14 after a single transfection on day 3 (data not shown). The total number of significantly differentially expressed genes in the presence of the miR-22-3p antagomir was 387 compared to preadipocytes (negative control) and 174 compared to 100 nM rosiglitazone (positive control).
Subsequent mRNA profiling was done by Next Generation Sequencing after a miR-22-3p antagomir single transfection (n = 3 per condition). In this experimental setting, the expression of 18 of the above 60 target genes seems to be significantly altered (fold changes and/or P values) in the presence of a miR-22-3p antagomir: they are CEBPD, CREB1, EIF4EBP2, KDM3A, KDM6B, KLF11, KLF6, LAMC1, MFGE8, NAA20, NCOA1, PPARA, PPARGC1B, PRDX3, RUNX2, SIRT1, TRPM8, and UCP1 (Supplementary Table S3).
By gene set enrichment analysis (Gene Ontology pathway database), we found that the number of gene sets significantly enriched (nominal P value <1%) in human adipocytes in culture was 129 in the presence of a miR-22-3p antagomir and 160 in the presence of 100 nM rosiglitazone (Supplementary Table S4). According to the Kyoto Encyclopedia of Genes and Genomes (KEGG) resource, it appears that the PPAR signaling pathway is activated in human adipocytes in culture in the presence of the miR-22-3p antagomir (Fig. 3). Analysis of transcriptome changes after treatment with the miR-22-3p antagomir, the positive control rosiglitazone, or an unrelated miRNA (miR-515-3p) by clustering based on expression intensity demonstrates different and distinct downstream effects of the miR-22-3p antagomir and miR-515-3p versus control or rosiglitazone (Supplementary Fig. S2). Modulation of gene expression by the miRNAs is more subtle than that observed with rosiglitazone.
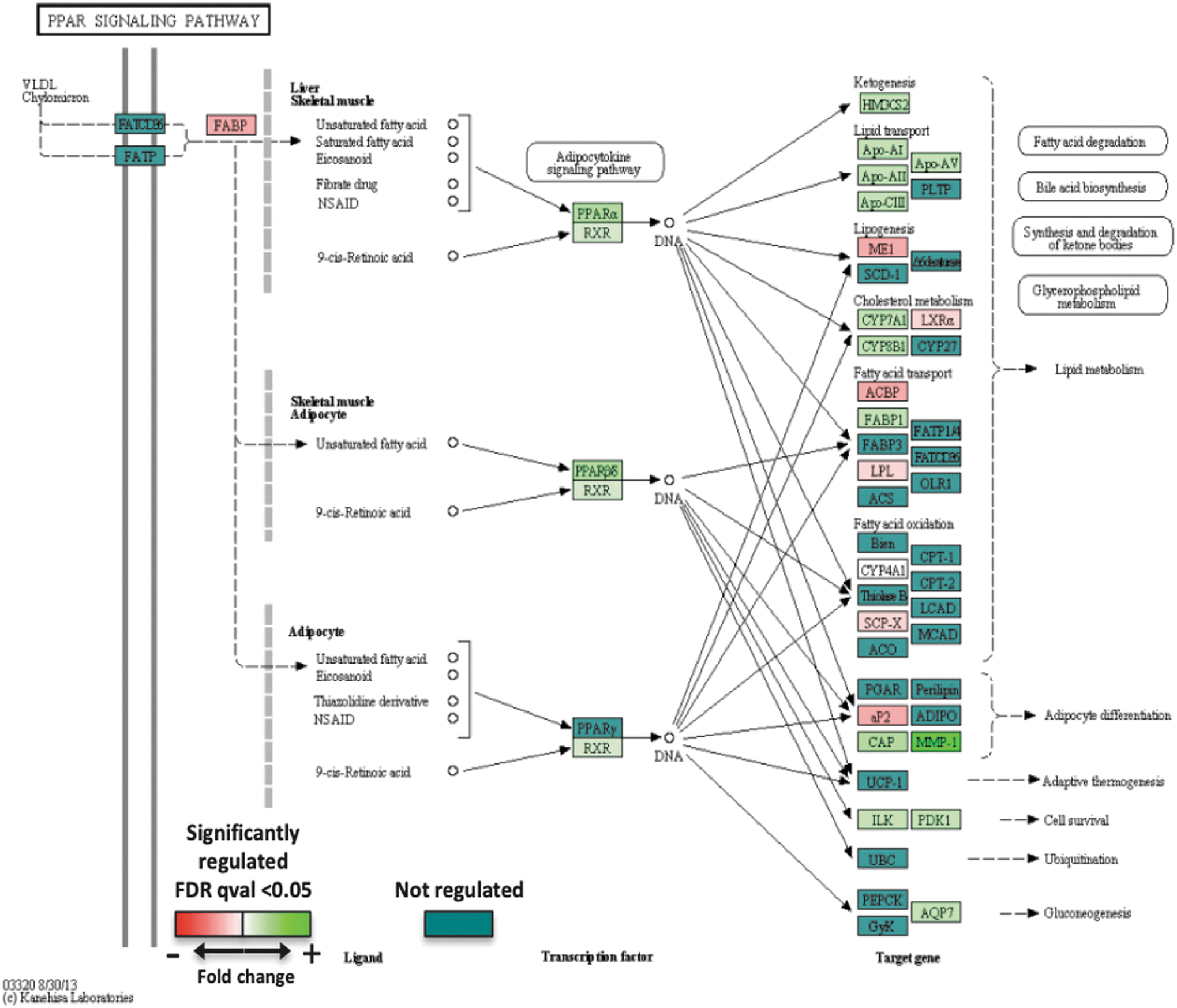
Activation of the PPAR pathway in primary cultures of human subcutaneous adipocytes in the presence of a miRNA 22–3p inhibitor (Kyoto Encyclopedia of Genes and Genomes [KEGG] resource).
Comparing the expression level of genes involved in the PPAR Signaling Pathway in the presence of KDM3A inactivation by knockout (publication by Tateishi et al. [20]) or KDM3A activation by miR-22-3p inhibition (our RNA Seq data), opposite effects of these two interventions were noted, thus suggesting that the antiobesity and beneficial metabolic effects of miR-22 inhibition involve the PPAR signaling pathway.
In vitro intracellular assessment of mitochondrial mass and activities, as well as EE, during miR-22 inhibition
In our model of primary culture of human subcutaneous adipocytes differentiated for 3 days, then transfected once with 50 nM of the miR-22-3p antagomir APT-110, and maintained in culture (n = 6 per condition), the following parameters were assessed on day 14:
Mitochondrial content/mass by MitoTracker (Fig. 4A)
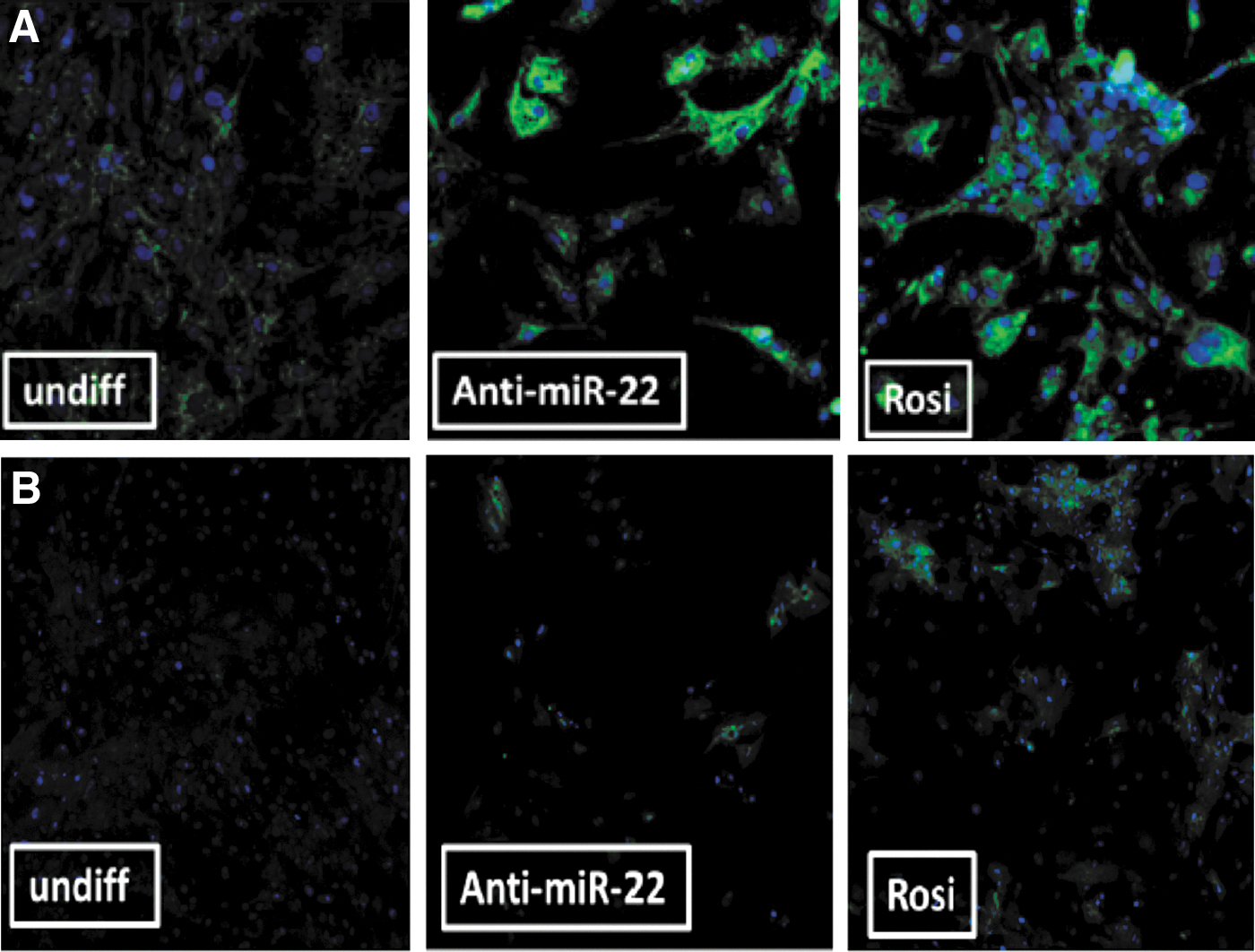
A significant increase of mitochondrial mass/activity was noted in the presence of the miR-22-3p antagomir, similar to that observed in the presence of a positive control (the PPARG agonist rosiglitazone, 100 nM throughout the 14-day experiment), although the cell density was significantly higher (three times more cells) in the presence of rosiglitazone, a well-known adipogenic agent. At day 14 of culture, about 64% ± 5% of cells seemed to have converted into “beige cells” with increased mitochondrial mass in the presence of miR-22-3p inhibition, similar to what was observed in the presence of rosiglitazone (61% ± 1%).
UCP-1 protein expression by immunofluorescence (Fig. 4B).
A significant increase of UCP1 immunostaining was noted in the presence of the miR-22-3p antagomir. The number of “UCP1 positive” cells was higher in the presence of the mir-22-3p antagomir (85% ± 3%, P ≤ 0.0001) than in the presence of rosiglitazone (58% ± 2%).
In vitro extracellular assessment of EE during miR-22 inhibition of human subcutaneous adipocytes in culture
Supplementary Figure S3A shows the OCR at baseline, then after addition of oligomycin (to assess ATP production and proton leak), FCCP (to assess maximum respiration), and antimycin A+rotenone (to assess reserve capacity and nonmitochondrial respiration) in control condition (BSA) and in the presence of rosiglitazone. Under basal conditions, rosiglitazone-treated adipocytes had higher OCRs than control adipocytes (fourfold). Rosiglitazone treatment increased proton leak (uncoupling), ATP turnover, maximum respiration (fourfold), reserve capacity, and nonmitochondrial respiration (Supplementary Fig. S3B), similar to the profile recently reported by Bartesaghi et al. [40]. To assess oxidation of endogenous and exogenous fatty acids in the presence of rosiglitazone, Seahorse measurements were performed in the presence of rosiglitazone alone (utilization of endogenous fatty acids), rosiglitazone+palmitate (utilization of exogenous fatty acids), or rosiglitazone+palmitate+etomoxir (blockade of utilization of exogenous fatty acids). Supplementary Figure S3C, D shows that the effects of rosiglitazone on OCR are mainly due to the utilization of endogenous fatty acids.
Under the same conditions, the effects of miR-22-3p inhibition on OCR of human adipocytes in culture are different (Supplementary Fig. S3E, F). Under basal conditions, miR-22-3p inhibitor-treated adipocytes had higher OCRs than control adipocytes (twofold) while maximum respiration is increased by 33%. The addition of palmitate to miR-22-3p inhibition induced a dramatic increase of nonmitochondrial respirations, as well as proton leak, suggesting that the miR-22-3p inhibitor acts through utilization of both endogenous and exogenous fatty acids, leading to enhanced respiratory chain uncoupling.
In vitro testing of miR-22-3p antagomirs ability to derepress luciferase expression in HeLa cells cotransfected with a miR-22-expression plasmid and a miR-22-3p luciferase reporter gene
Supplementary Figure S4 shows that the inhibition of miR-22-3p reporter luminescence by miR-22 expression was very robust, ∼30-fold. Negative controls (anti-miR-133 and inhibitor) did not significantly derepress luciferase expression by miR-22, as predicted. Several miR-22-3p antagomirs derepressed luciferase expression in a dose-dependent manner, with maximal derepression of about 30% of controls. No cytotoxicity was observed in all conditions tested.
Beneficial metabolic effects of miR-22 inhibition in mouse model of diet induced obesity
The metabolic effects of miR-22 inhibition were assessed in several studies in diet induced obesity (DIO) C57BL/6J male mice of various ages [30]. Three studies are presented in the following sections.
Reduction of weight gain induced by high fat diet in young adult mice treated for 8 weeks with a miR-22 antagomir
A study of miR-22-3p inhibition was conducted in the initial phase of DIO in 6-week-old mice receiving saline, a scrambled miRNA antagomir or a miR-22-3p antagomir (Exiqon; 15 mg/kg). As shown in Fig. 5A, there was a steady increase of body weight over the 8-week duration of the study in all three groups related to the ad libitum consumption of a 60% high fat diet. However, body weight gain over time was lower in the miR-22-3p antagomir group, reaching statistical significance as of week 4 of treatment. At the end of 8 weeks of high fat diet (14-week-old mice), the miR-22-3p antagomir group gained 29% less body weight than the saline group and 25% less body weight than the scrambled miRNA group (P ≤ 0.05).
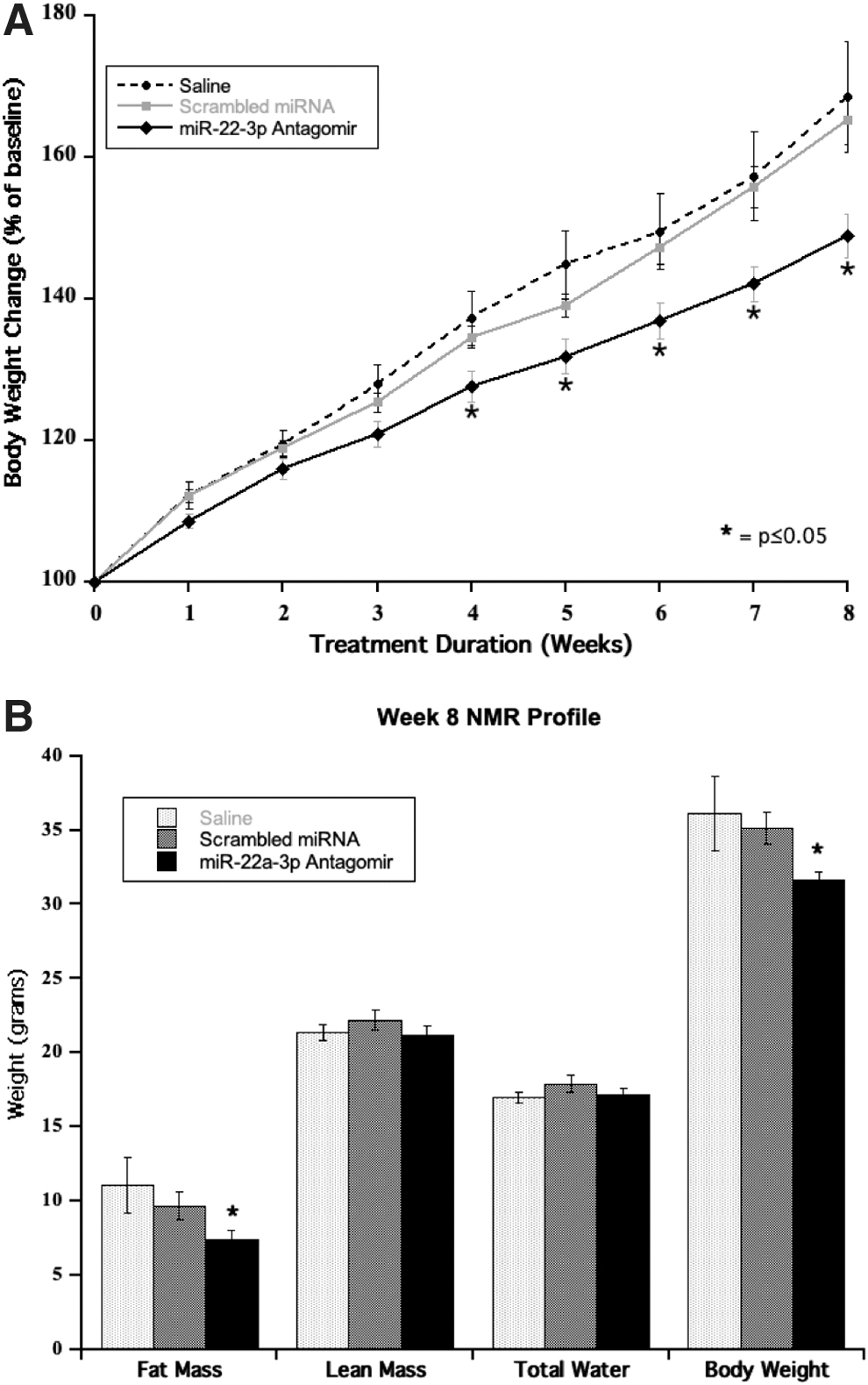
The reduced weight gain in the miR-22-3p antagomir group was not explained by a lower food consumption as this group seemed to eat more food over the 8 weeks of treatment (50.83 ± 0.06 g, 51.88 ± 0.10 g, and 58.55 ± 0.15 g over 8 weeks in the saline, scrambled miRNA antagomir, and miR-22-3p antagomir groups, respectively, P ≤ 0.01).
The miR-22-3p antagomir group showed a significant reduction of fat mass and total body weight by NMR analysis at week 8 of the study, whereas its lean and total water fractions were similar to those of the control groups (Fig. 5B).
In addition, the miR-22-3p antagomir group displayed a significant improvement of random nonfasting blood glucose (226 ± 12 mg/dL vs. 289 ± 9 mg/dL, P ≤ 0.0015), serum insulin (6.3 ± 1.09 ng/mL vs. 10 ± 2 ng/mL, P ≤ 0.01), and serum cholesterol (126 ± 4 mg/dL vs. 175 ± 14 mg/dL, P ≤ 0.0011) versus the saline group at week 8. Liver function tests were within normal ranges in all groups.
Tissues were collected at the end of the study. There was no difference in heart, liver, and spleen weights across the three groups. The group treated with the miR-22-3p antagomir displayed a significant reduction of inguinal (0.577 ± 0.055 g vs. 1.074 ± 0.194 g, −46%, P ≤ 0.032), perirenal (0.489 ± 0.053 g vs. 0.924 ± 0.107 g, −47%, P ≤ 0.017), and subscapular fat (0.127 ± 0.009 g vs. 0.381 ± 0.101 g, −67%, P ≤ 0.004) depots in comparison to the saline group on the same 60% high fat diet.
Prevention of obesity induced by high fat diet in mature adult mice treated for 8 weeks
A study of miR-22-3p inhibition was conducted in the established phase of DIO in 13-week-old C57BL/6J adult male mice who were started on 60% high fat diet ad libitum at 6 weeks of age. At the end of 8 weeks of active treatment (22-week-old mice), the mice receiving the miR-22-3p antagomir (Exiqon; 15 mg/kg) kept a stable weight (35.51 ± 0.76 g at week 8 of treatment vs. 34.98 ± 0.54 g at week 0), whereas the mice on the 60% high fat diet receiving saline injections kept gaining weight, reaching a 30% weight increase at week 8 of treatment (45.85 ± 0.93 g at week 8 vs. 35.14 ± 0.47 g at week 0, P ≤ 0.0001) (Fig. 6A). The mice that switched back to a normal chow at week 12 of age initially dropped their body weight for 2 weeks because of reduced consumption of less palatable food and then displayed an age-related moderate weight gain (+8%) at the end of the study (32.08 ± 0.61 g at week 8 vs. 29.71 ± 0.40 g at week 0, P ≤ 0.004) while receiving saline injections (Fig. 6B).

There was no difference in food consumption between the saline and miR-22-3p antagomir groups on 60% fat diet (70.57 ± 1.51 g and 69.98 ± 1.35 g over 8 weeks of treatment in the saline and miR-22-3p antagomir groups, respectively) that could have explained the body weight differences. As expected, the group on 10% fat regular chow and saline injections ate less food (49.71 ± 0.23 g).
Body composition was measured by NMR at weeks 0 and 8 of active treatment. At initiation of treatment, body weight and fat mass were lower in the group that has been switched from 60% fat diet to 10% fat diet a week before. At week 8, fat mass was 29% lower in the miR-22-3p antagomir group on 60% fat diet than that in the group on saline and 60% fat diet (10.50 ± 0.71 g vs. 14.75 ± 1.36 g, P ≤ 0.0015). Lean mass was similar across the three groups at week 8 of treatment.
In addition, the miR-22-3p antagomir group on 60% fat diet displayed a significant improvement of random nonfasting blood glucose versus the group on saline and 60% fat diet (203 ± 4 mg/dL vs. 282 ± 11 mg/dL, P ≤ 0.0001) at the end of the study (week 22). Surprisingly, the blood glucose at week 22 of the miR-22-3p antagomir group on 60% fat diet was lower compared with the group on saline that was switched back to 10% fat diet at week 13 (203 ± 4 mg/dL vs. 253 ± 10 mg/dL, P ≤ 0.0017), possibly reflecting the metabolic insult (60% fat diet) these animals experienced between weeks 6 and 13 of age before they were switched back to normal chow. Also at week 22, serum insulin (2.56 ± 0.22 ng/mL vs. 6.99 ± 0.71 ng/mL, P ≤ 0.001) and serum cholesterol (109 ± 1 mg/dL vs. 188 ± 7 mg/dL, P ≤ 0.0001) were improved in the miR-22-3p antagomir group on 60% fat diet versus the saline group on 60% fat diet, reaching values similar to those of the group on 10% fat diet (insulin: 1.80 ± 0.16 ng/mL, cholesterol: 117 ± 5 mg/dL). Liver function tests were within normal ranges in all groups.
Body temperature was measured with a rectal probe during weeks 1, 4, and 8 of active treatment. No difference was recorded between the three groups. At the end of the study, body temperature was 35.51°C ± 0.11°C, 35.80°C ± 0.17°C, and 36.04°C ± 0.12°C in the 10% fat saline, 60% fat saline, and 60% fat miR-22-3p antagomir groups, respectively (P = 0.742, Kruskal–Wallis Rank Sum test). This suggests that the weight loss observed in the miR-22-3p antagomir group was not caused by hyperthermia.
Tissues were collected at the end of the study (Table 1). There was no difference of liver and spleen weights across the three groups. The 60% high fat diet group treated with the miR-22-3p antagomir displayed a size reduction of inguinal (−48%), perirenal (−34%), and subscapular fat (−38%) depots compared to the 60% high fat diet group treated with saline (all P ≤ 0.001).
Organ/Tissue Weights in Mature Adult Mice (22-Week-Old Mice) Treated for 8 Weeks
P ≤ 0.0001, miR-22-3p antagomir group on 60% fat diet group versus saline group on 60% fat diet.
SEM, standard error of the mean.
A similar study of miR-22-3p inhibition was conducted in 13-week-old C57BL/6J adult male mice, which were treated with saline or AptamiR's proprietary miR-22-3p antagomir APT-110 (15 mg/kg) for 6 weeks. The selection of the 15 mg/kg dose was based on body weight and fat mass recorded during a dose–response study testing six doses of APT-110 from 1.25 to 30 mg/kg. At the end of 6 weeks of active treatment (20-week-old mice), the body weight of the mice receiving the miR-22-3p antagomir APT-110 was similar to their body weight at randomization (36.16 ± 0.67 g at week 6 of treatment vs. 36.08 ± 0.74 g at week-1), whereas the mice on the 60% high fat diet receiving saline injections kept gaining weight, reaching a 16% weight increase at week 6 of treatment (41.73 ± 1.07 g at week 6 vs. 36.05 ± 0.79 g at week 0, P ≤ 0.0001). On day 43 of active treatment, the mice were fasted for 4 h, and blood glucose was measured by glucometer. Blood glucose was 231 ± 9 mg/dL in the saline group and 189 ± 12 mg/dL in the miR-22-3p antagomir APT-110 group (P ≤ 0.0236).
Blood was also collected at the time of necropsy. At that time, nonfasting blood glucose was 343 ± 9 mg/dL in the saline group and 210 ± 12 mg/dL in the miR-22-3p antagomir APT-110 group (P ≤ 0.0001). Blood cholesterol was 164 ± 6 mg/dL in the saline group and 110 ± 5 mg/dL in the miR-22-3p antagomir APT-110 group (P ≤ 0.0001).
Tissues were collected at the end of the study. There was no difference in heart and liver weights between the two groups. The 60% high fat diet group treated with the miR-22-3p antagomir displayed a weight reduction of inguinal (−36%), perirenal (−29%), and subscapular fat (−39%) depots compared to the 60% high fat diet group treated with saline (Table 2). Browning of subscapular tissue was present in the miR-22-3p antagomir with a percentage of WAT calculated (19% ± 1% vs. 47% ± 3% in the saline group, P ≤ 0.0004). Histologic examination of fat tissues at the end of the study (hematoxylin and eosin staining) revealed no evidence of cellular toxicity or death, but a significant reduction of adipocyte cell size in the inguinal (subcutaneous) and subscapular regions, but not of the perirenal (visceral) region, which is known for its low level of UCP1 expression (Fig. 7).
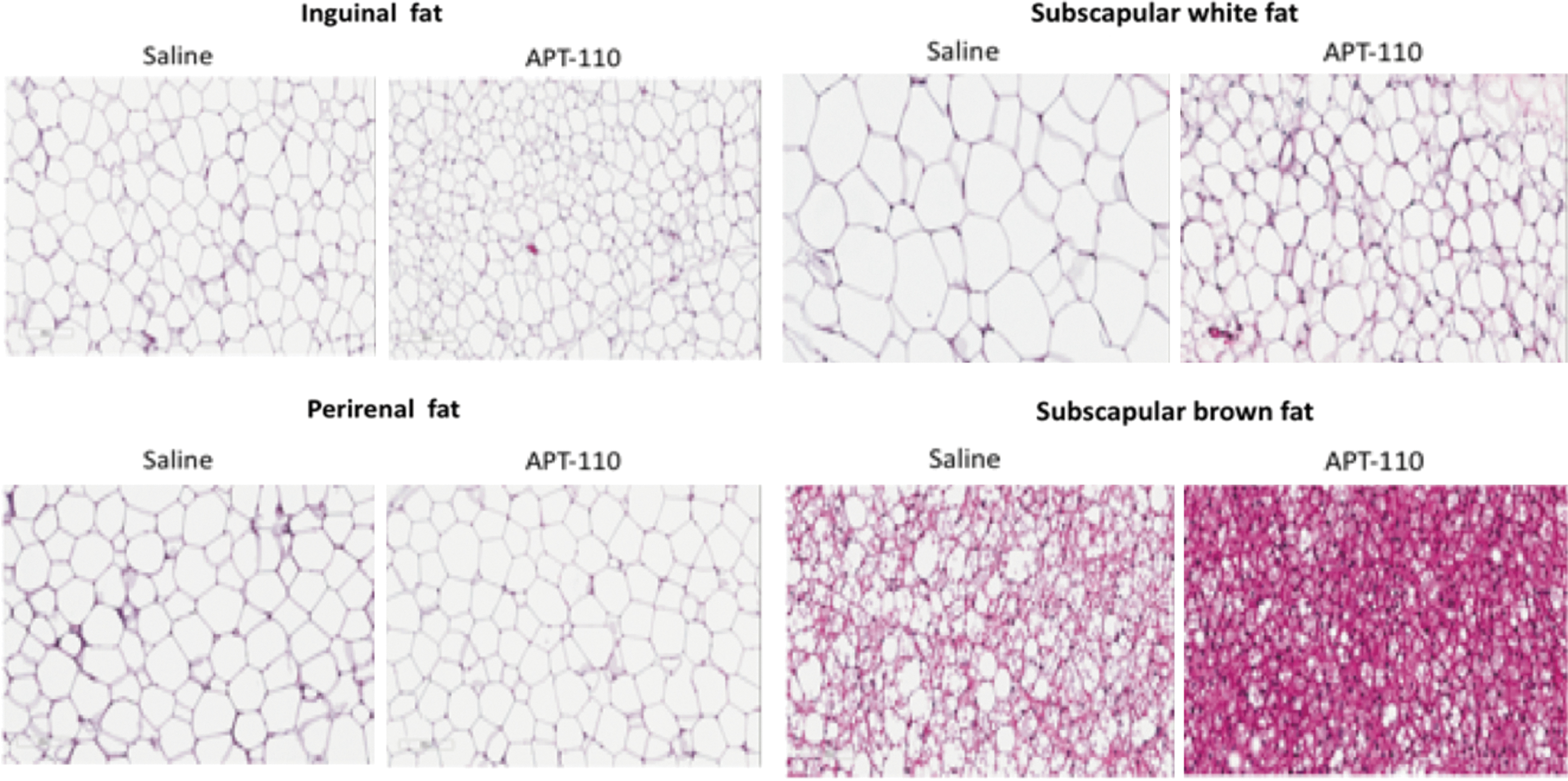
Hematoxylin and eosin staining of various fat tissues of 20-week-old C57Bl/J6 adult on 60% high fat diet alone (saline) or in the presence of the miR-22-3p antagomir APT-110 (APT-110) collected at the end of 6 weeks of treatment started at week 14 of age. Average cell perimeter (pixels)—inguinal fat: 279 ± 7 (saline group), 233 ± 15 (miR-22 antagomir group), P < 0.02; perirenal fat: 333 ± 4 (saline group), 326 ± 13 (miR-22 antagomir group), P = 0.5726; and subscapular white fat: 428 ± 28 (saline group), 327 ± 20 (miR-22 antagomir group), P < 0.02. For subscapular brown fat, the smaller size of the cells prevented an accurate measurement of their cell perimeter. Instead, the percentage of fat was calculated. It was 47% ± 3% in the saline group versus 19% ± 1% in the miR-22 antagomir group (P ≤ 0.0004).
Organ/Tissue Weights in Mature Adult Mice (20-Week-Old Mice) Treated for 6 Weeks
Discussion
miRNAs are involved in many metabolic functions (adipogenesis, obesity, browning of adipocytes, insulin resistance, lipids, and glucose metabolism) and have been shown to be dysregulated in various cardiometabolic diseases [41–46]. As a new class of therapeutic agents, the mechanism of action of miRNA analogs (one drug-multiple targets concept) makes them particularly attractive to treat complex diseases like diabesity and NAFLD/NASH [27,47,48]. Furthermore, recent elegant studies have shown that adipose tissue constitutes a major source of circulating exosomal miRNAs regulating gene expression in distant tissues like liver and muscle where they modulate insulin sensitivity and inflammation [33,49,50]. Administration of a high fat diet in mice increases the amount of 10 miRNAs in circulating exosomes, including miR-22-3p that modulates glucose and lipid metabolism in mice [34].
Our initial experimental and in silico work with various miR-22-3p antagomirs suggested that miR-22 is a good target to investigate in the context of obesity and related metabolic disorders. Our in vitro experiments in human adipocytes and in vivo studies in mice suggest that miR-22-3p inhibition:
modulates the expression of several metabolic genes such as KDM3A, KDM6B, PPARA, PPARGC1B, and SIRT1
stimulates lipid metabolism, fatty acid oxidation, and thermogenesis in human subcutaneous adipocytes
reduces the accumulation of fat in the mouse model of DIO as tested in animals of various ages
modifies the phenotype of adipocytes with cellular size reduction, increase of mitochondrial mass/activity, and UCP1 expression (the “Browning Effect”)
increases EE
has a dramatic effect on glucose homeostasis and insulin sensitivity
reduces levels of circulating cholesterol
alters favorably the release of adipokines (reduction of pro-inflammatory leptin while anti-inflammatory adiponectin remains steady)
All these metabolic benefits were observed despite the fact that the animals kept eating the same amount of high fat diet, no hyperthermia was recorded, and physical activity remained unchanged. The effects of miR-22-3p inhibition on energy homeostasis take two weeks to develop, reflecting the mechanisms(s) of action of oligonucleotide therapeutic agents. This profile is clearly different from the one of central acting agents modulating appetite and/or satiety whose effects mainly occur during the first 2 weeks of treatment [51]. It now remains to establish if the same beneficial effects can be observed in human clinical trials.
Our findings are in agreement with findings from other investigators. In transgenic mice with enhanced cardiac expression of miR-22, the expression of PPARA, PPARGC1A, and SIRT1 (all involved in energy substrate metabolism) was repressed [52]. Our findings represent the opposite pattern of what was observed in KDM3A knockout mice in regard to the expression of genes involved in PPAR signaling, fatty acid oxidation, and thermogenesis [20]. In addition, the expression of SIRT1 in visceral adipose tissue of obese subjects was correlated negatively with the expression of miR-22-3p [53]. In H9c2 cardiomyocytes, a miR-22 mimic aggravated, whereas a miR-22 inhibitor significantly attenuated ischemia/reperfusion-induced mitochondrial damage [54]. Silencing of SIRT1 abolished the protective effect of miR-22 inhibition against ischemia/reperfusion-induced mitochondrial dysfunction and cell injury in cardiomyocytes. Finally, miR-22 role in gluconeogenesis, lipid metabolism, EE, and fat mass was recently confirmed by several independent academic groups [55–58].
Investigators have studied the role of several miRNAs in obesity, diabetes, and lipid disorders as recently summarized [59–61]. For instance, Dimmeler and colleagues reported that a locked nucleic acid-modified anti-miR-21 reduced body weight, as well as adipocyte size and serum triglycerides, over the course of 18 weeks of treatment of leptin receptor deficient (db/db) mice [62].
The concept of modulation of energy-expending adipocytes in human adults is now firmly established [63–66]. Increase of EE as an effective treatment of obesity and related disorders is appealing and the target for drug research and development [13,67–69]. A 15% increase of EE is believed to be sufficient to safely achieve a significant weight and fat mass reduction while providing meaningful improvement of metabolic parameters [7,70–72]. However, one should proceed with caution when modulating EE in view of the 2,4-dinitrophenol (DNP) experience [73]. Several approaches to “browning of white adipocytes” are currently tested, including activation of thyroid hormone receptors [74], activation of the beta3-adrenergic receptors [75,76], development of 2,4-DNP derivatives targeting the liver [77], niclosamide analogs that are intestinally absorbed [78], food ingredients [79], and transplantation of BAT [80]. Interestingly enough, double and triple combinations of incretins (gut hormones) are currently developed for the treatment of type 2 diabetes mellitus and/or obesity [81–86].
Footnotes
Acknowledgments
The authors thank the following individuals: Dr. Huntington F. Willard, President and Director of the Marine Biological Laboratory, for his scientific input and in-depth discussions. Dr. Daniel Ricquier, Professor at University Paris Descartes and member of the French Academy of Sciences, for sharing his expertise on thermogenesis and his critical reading of the article. Dr. Eric Clauser, Director of the INSERM School, for his in-depth review of the data and article.
Author Disclosure Statement
Ownership: M.T. is the Founder and a Shareholder of AptamiR Therapeutics, Inc. Intellectual property: M.T. is the inventor of several patents assigned to AptamiR Therapeutics, Inc. C.E declares no conflicts of interest.
Funding Information
All funding for this R&D work came from AptamiR Therapeutics, Inc.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.