
Abstract
C promoter binding factor 1 (CBF1) (alias RBPJ) is a critical transcription factor involved in Notch signaling. The activation of Notch signaling through CBF1 maintains the angiostatic state of endothelial cells suppressing angiogenesis, that is, the formation of new blood vessels. Vascular endothelial growth factor (VEGF) induces angiogenesis by promoting the proteasomal degradation of CBF1, in addition to endothelial cell proliferation. To date, angiogenic inhibitors targeting VEGF have been successfully used in clinics for cancer and age-related macular degeneration. Most antiangiogenic drugs, however, only target VEGF or VEGF receptors. In this study, to expand the repertoire of antiangiogenic therapeutics, we developed 15 single-stranded deoxyribonucleic acid (ssDNA) aptamers capable of binding to CBF1 with high affinity (Kd; 10–300 nM). To this end, systematic evolution of ligands by the exponential enrichment (SELEX) method was applied. One of the CBF1-binding ssDNA aptamers, Apt-3, inhibited angiogenesis through the activation of Notch signaling in vitro. We found that Apt-3 directly interacted with the LAG1 domain of CBF1. We suggest that the Apt-3 ssDNA aptamer may contribute to the development of a novel angiogenic inhibitor, which does not target VEGF.
Introduction
Aptamers are a class of single-stranded deoxyribonucleic acid (ssDNA) or ribonucleic acid (ssRNA) oligonucleotides that specifically bind to target molecules. These agents have been recognized as promising novel therapeutics owing to a number of advantages (eg, short time/low costs of production, high affinity) [1,2]. The main strength of aptamers is the availability of a reliable and efficient method for their development, that is, the systematic evolution of ligands by exponential enrichment (SELEX) [3,4]. In general, aptamer candidates are designed as 70–100 nucleotide molecules that include a random region in the middle, flanked by fixed primer sequences. Therefore, in vitro SELEX selection starts by incubation of target molecules with a pool of 1014–1016 ssDNAs or ssRNAs [2]. This extremely large repertoire of single-stranded oligonucleotides enables to develop aptamers with high affinity and specificity for target molecules [1]. Pegaptanib, a vascular endothelial growth factor (VEGF)-specific RNA aptamer, has been clinically approved for age-related macular degeneration (AMD) therapy [5].
Angiogenesis is the process of new blood vessel formation from preexisting blood vessels. During angiogenesis, endothelial cells lining along the inside of blood vessels receive a variety of stimuli resulting in drastic changes of their characteristics [6]. Angiogenesis plays a critical role not only in physiological processes (eg, development, pregnancy, and tissue regeneration) but also in pathological processes (eg, inflammation, would healing, and tumor growth) [7]. Angiogenesis is regulated by a well-characterized mechanism consisting of a sort of “tug-of-war” between VEGF-dependent proliferative and Notch-dependent cytostatic signaling [8–10]. In angiostatic responses, endothelial cells receive signals generated from the interaction of plasmalemmal Notch receptors with Notch ligands, such as DLL4, which are expressed on the cell surface of neighboring endothelial cells [10]. The interaction between DLL4 and Notch receptors induces the γ-secretase-mediated cleavage of Notch intracellular domain (NICD) from Notch receptor [10]. Then, NICD translocates to the nucleus and forms complexes with various transcription factors and cofactors on target DNA sequences, leading to the upregulation of different target genes (eg, HEY1, HEY2, and HES) [11]. The activation of Notch signaling results in the inhibition of endothelial cell proliferation and in the maintenance of the angiostatic state of cells. Vice versa, VEGF signaling induces angiogenesis by reverting the Notch-dependent angiostatic state, in addition to activating cell proliferation [12]. Endothelial cell stimulation by VEGF through the VEGF receptor (VEGFR) upregulates the expression of Bcl6-associated zinc finger protein (BAZF), a substrate-recognition receptor for cullin-3 (CUL3)/RING ubiquitin E3 complex [11]. BAZF then recruits C promoter binding factor 1 (CBF1), one of the main transcription factors of Notch signaling, to the CUL3–E3 complex, leading to the proteasomal degradation of CBF1 [11]. VEGF-dependent CBF1 degradation turns off Notch-dependent angiostatic signaling, and angiogenesis can be initiated [11]. Specific disruption of CBF1 in endothelial cells strongly induces spontaneous angiogenesis and vascular sprouting in various mouse tissues [13]. Thus, CBF1 is a critical regulator of angiogenesis.
To date, antiangiogenic drugs targeting VEGF and its receptors have been developed. Humanized monoclonal antibodies against VEGF (eg, bevacizumab) and VEGFR tyrosine kinase inhibitors (eg, sunitinib) exhibit antitumor effects in colorectal cancer, lung cancer, and renal cell carcinoma [14]. The anti-VEGF RNA aptamer, pegaptanib, is currently used for the treatment of AMD [5]. Despite the good antiangiogenic effects of the VEGF inhibitors in vivo, adverse events (eg, hypertension, gastrointestinal perforation, and thrombotic events) and acquisition of drug resistance are major drawbacks for their clinical use [15–17]. The VEGF inhibitors lose their antiangiogenic activities by the long-term administration during therapy for ocular disorders [18]. Furthermore, all clinically available antiangiogenic therapeutics only target VEGF and VEGFR. In this study, we developed a CBF1-targeting ssDNA aptamer, termed Apt-3, which inhibited angiogenesis in vitro through the activation of Notch signaling.
Materials and Methods
Antibodies
The following antibodies were purchased from the indicated manufacturers: rabbit anti-CBF1 antibody (5313S, dilution 1:1,000 for western blotting, immunofluorescence; Cell Signaling Technology), rabbit anti-HEY2 antibody (10597-1-AP, dilution 1:100; Proteintech), mouse anti-GAPDH antibody (5A12, dilution 1:6,000; Wako), mouse anti-Myc antibody (9E10, dilution 1:1,000; Santa Cruz), mouse anti-FLAG antibody (M2, dilution 1:1,000; Sigma), goat Cy3-conjugated anti-rabbit IgG antibody (A10520, dilution 1:2,000; Molecular Probes), goat Alexa488-conjugated anti-mouse IgG antibody (A11001, dilution 1:2,000; Molecular Probes), HRP-conjugated anti-mouse IgG antibody (W4021, dilution 1:2,000; Promega), and HRP-conjugated anti-rabbit IgG antibody (W4011, dilution 1:2,000; Promega).
Plasmids
The CBF1 gene was amplified from a vector containing the human CBF1 cDNA [11] using the following pairs of primers: 5′-ATGGACCACACGGAGGGCTC-3′ (sense primer) and 5′-TTAGGATACCACTGTGGCTG-3′ (antisense primer). The polymerase chain reaction (PCR) products were introduced into the blunt ends of the pEU-N-bls vector (a king gift from Ms. Yuki Tanaka, Ehime University). The CBF1 was amplified using the following pairs of primers: 5′-ATGGACCACACGGAGGGCTC-3′ (sense primer) and 5′-GGATACCACTGTGGCTGTAG-3′ (antisense primer). The PCR products were introduced into the blunt ends of the pEU-C-FLAG vector (a king gift from Ms. Yuki Tanaka, Ehime University). The Myc-CBF1 was amplified using the following pairs of primers: 5′-ATGGAACAAAAACTCATCTCAGAAGAGGATCTGATGGACCACACGGAGGGCTC-3′ (sense primer) and 5′-TTAGGATACCACTGTGGCTG-3′ (antisense primer). The PCR products were introduced into the blunt ends of the CSII-CMV-MCS-IRES2-Bsd vector (a king gift from Dr. Hiroyuki Miyoshi, RIKEN). The LAG1 domain of CBF1 was amplified using the following pairs of primers: 5′-ATGGTACTTATTCTTCATGC-3′ (sense primer) and 5′-GTCAGCATTTTTCAATGACTGC-3′ (antisense primer). The PCR products were introduced into the blunt ends of the pEU-C-FLAG vector. CBF1 (ΔLAG1) was generated with the following pairs of primers: 5′-TTATGCATTGCCTCAGGAACAAAGG-3′ (sense primer) and 5′-TGTTTGATCCCCTCGCTCTTTTAAA-3′ (antisense primer). CBF1 (ΔBTD) was generated with the following pairs of primers: 5′-ACAATCATTAGCACAGATAAGGCAG-3′ (sense primer) and 5′-GTCAGCATTTTTCAATGACTGCTTC-3′ (antisense primer). CBF1 (ΔLAG1/BTD) was generated with the following pairs of primers: 5′-ACAATCATTAGCACAGATAAGGCAG-3′ (sense primer) and 5′-TGTTTGATCCCCTCGCTCTTTTAAA-3′ (antisense primer). CBF1 truncated mutants were generated with the following pairs of primers: 5′- TGCATTGCCTCAGGAACAAAGGTGG-3′ (Mutant 1-sense primer) and 5′-TGGGCAAAAAAACCTTTTTTCATTT-3′ (Mutant 1-antisense primer), 5′-TGCATTGCCTCAGGAACAAAGGTGG-3′ (Mutant 2-sense primer) and 5′- ACCATCGCGTTCCATTTGTTCTTTT-3′ (Mutant 2-antisense primer), 5′-TGCATTGCCTCAGGAACAAAGGTGG-3′ (Mutant 3-sense primer) and 5′- CTGCATTTCTTGGTCACTATTTCCT-3′ (Mutant 3-antisense primer), 5′- TGCATTGCCTCAGGAACAAAGGTGG-3′ (Mutant 4-sense primer) and 5′- CTTGTCTGAGTCAGATATATACAAT-3′ (Mutant 4-antisense primer), 5′- CGAAAGCACTTCATGTTGTCTGTAA-3′ (Mutant 5-sense primer) and 5′- TGTTTGATCCCCTCGCTCTTTTAAA-3′ (Mutant 5-antisense primer), 5′- AGCAAGCGGATAAAAGTCATCTCCA-3′ (Mutant 6-sense primer) and 5′- TGTTTGATCCCCTCGCTCTTTTAAA-3′ (Mutant 6-antisense primer). The His-CBF1-pEU vector is a king gift from Ms. Yuki Tanaka, Ehime University.
Production of recombinant proteins by wheat germ extracts
Transcription and translation for the production of biotinylated or FLAG-tagged recombinant human CBF1 proteins were performed using the WEPRO7240 Expression Kit (Cell Free Science) as reported previously [19].
AlphaScreen
The in vitro binding assays were performed as previously described using an AlphaScreen IgG (Protein A) Detection Kit (Perkin Elmer) [20] with slight modifications. Briefly, 10 μL of detection mixture containing FLAG-tagged CBF1 recombinant proteins, 25 nM 5′-terminally biotinylated aptamers (Eurofins Genomics), 100 mM Tris-HCl (pH 7.5), 0.01% Tween 20, 1 μg/mL anti-FLAG antibody (Wako), 1 mg/mL bovine serum albumin (BSA), 0.06 μL of streptavidin-coated donor beads, and 0.06 μL of anti-IgG acceptor beads were added to each well of a 384-well OptiPlate followed by incubation at room temperature for 1 h. Luminescence was detected by the AlphaScreen detection program using EnSpire (Perkin Elmer).
In vitro aptamer selection
The initial pool of ssDNA (5′-ATAGGAGTCGACCGACCAGANNNNNNNNNNNNNNNNNNNNNNNNNNNNNNGTGCGTCTACATCTAGACTC-3′) was purchased from Tsukuba Oligo Service, in which a mixture of the four phosphoramidites with an A:C:G:T ratio of 3:3:2:2 was used to obtain an even distribution of the four bases at each position in the random region (N) [21]. In the first round, 4.6 nmol (2.8 × 1015 molecules) of the initial ssDNA pool in 50 μL of 1 × S-buffer (100 mM Tris-HCl (pH 8.0), 100 mM NaCl, 5 mM KCl, 1 mM MgCl2, 0.1% Tween 20, 1 mg/mL BSA) was incubated at room temperature for 30 min with streptavidin-coated magnetic (SAv-mag) beads (Promega) that were prewashed with 1 × S-buffer, and then the supernatant was recovered to remove ssDNA bound to SAv-mag beads. This negative selection was repeated thrice. The supernatant in the third negative selection was incubated at room temperature for 30 min with SAv-mag beads on which 100 pmol of Biotin-CBF1 had been immobilized. After the beads were washed thrice with 200 μL of 1 × S-buffer, the bound ssDNA-CBF1 complexes were denatured by heating at 95°C for 5 min with 50 μL of 1 × E-buffer [1 × sample buffer with 10% 3-mercapto-1,2-propanediol (Wako), 4 M urea, 5 mM ethylenediaminetetraacetic acid (EDTA)]. The eluted ssDNA in the supernatant was purified with a QIA Nucleotide Removal Kit (QIAGEN) and then moderately amplified by PCR using Tks Gflex DNA Polymerase (Takara Bio) with the following primers: a forward primer (F-pri: 5′-ATAGGAGTCGACCGACCAGA-3′) and a 5′ phosphorylated reverse primer (pR-pri: 5′-pGAGTCTAGATGTAGACGCAC-3′). The PCR condition was (98°C for 10 s, 60°C for 15 s, 68°C for 10 s)/cycle. The PCR products (purified with the purification kit in the fifth and subsequent rounds) were treated with 10 U of λ exonuclease (NEB) at 37°C for 2 h to digest the 5′ phosphorylated antisense strand. The resulting ssDNA aptamer pool was purified by negative selection in the next round. The process of purification leads to refolding of the aptamer pool. The selection cycle was repeated 14 times with progressively increasing stringency (specifically, gradually decreasing concentrations of Biotin-CBF1 and ssDNA) (Table 1). To minimize the amplification bias, number of the PCR cycle and volume of the PCR template were also gradually reduced as the selection cycle proceeded (Table 1).
The Concentrations of Biotin-CBF1 and ssDNA and PCR Conditions for Amplification of the ssDNA Pool in Each SELEX Round
CBF1, C promoter binding factor 1; PCR, polymerase chain reaction; SELEX, systematic evolution of ligands by the exponential enrichment; ssDNA, single-stranded deoxyribonucleic acid.
Sequencing
Adapter DNA sequences that are required for sequencing were added to each side of the purified PCR products in the 14th round by PCR with another forward primer (5′-CCATCTCATCCCTGCGTGTCTCCGACTCAGATAGGAGTCGACCGACCAGA-3′) and another reverse primer (5′-CCTCTCTATGGGCAGTCGGTGATGAGTCTAGATGTAGACGCAC-3′). The PCR products were agarose gel-purified with an illustra GFX PCR DNA and Gel Band Purification Kit (GE Healthcare) and then sequenced with a next-generation sequencer, Ion PGM (Thermo Fisher Scientific), according to the manufacturer's protocol. Of the 198,955 read sequences, 17,140 were randomly chosen for clustering into several groups with the same sequence. Table 2 shows the sequences of the 15 most abundant ssDNA aptamers.
List of the CBF1-Bound ssDNA Aptamers Developed in this Study
The sequence of negative control-dA30 (NC-dA30) aptamer used in this study is also shown. The percentage of abundance in 14th pool is a ratio for the number of reads/total reads (17,140 reads).
n.d, not detected.
Determination of dissociation constants
Different amounts (0.3–15 pmol) of an ssDNA aptamer (Eurofins Genomics) were individually incubated at room temperature for 30 min with 0.3 pmol of Biotin-CBF1 immobilized on SAv-mag beads in 30 μL of 1 × S-buffer. After the beads were washed thrice with 50 μL of 1 × S-buffer, the bound aptamer-CBF1 complexes were denatured with 50 μL of 1 × E-buffer as described above. Each eluted aptamer was purified and then quantified by qPCR using a GoTaq qPCR Master Mix (Promega) with F-pri and pR-pri (or Rm-pri: GAGTCTAGATGTAGACGCCAC for Apt-14 and Apt-15). A series of serially diluted aptamers dissolved in 1 × E-buffer and purified were used for the calibration line. To determine the dissociation constant (Kd), the data were fitted to the equation f = fmax × [Apt]/([Apt] + Kd), where f is the complex formation rate, fmax is the maximum formation rate, and [Apt] is the concentration of free aptamer, by the nonlinear least squares method [21].
Surface plasmon resonance
Surface plasmon resonance (SPR) experiments to determine dissociation constants were performed using a Biacore T200 (Cytiva) according to the manufacturer's instructions at Translational Research Center, Fukushima Global Medical Science Center. The Sensor Chip SA, Amine Coupling Kits, and HBS-EP+ buffers were purchased from Cytiva. Production and purification of His-tagged recombinant human CBF1 protein were performed using the WEPRO7240H Expression Kit (Cell Free Science) according to the manufacturer's instructions. The purified His-tagged human CBF1 protein and 5′-terminally biotinylated Apt-3 (Eurofins Genomics) were used as the analyte and ligand, respectively. Biacore T200 Evaluation Software 2.0 (Cytiva) was used for single-cycle kinetics analyses.
Cell culture
Human umbilical vein endothelial cells (HUVECs) were obtained from Lonza. HUVECs were grown at 37°C with 5% CO2 in the endothelial growth medium EGM-2 (Lonza). All experiments were performed with HUVECs at passages 2–4. HEK293T cells were maintained at 37°C with 5% CO2 in Dulbecco's modified Eagle's medium (Wako) supplemented with 10% fetal bovine serum (FBS), 20 U/mL penicillin, and 100 μg/mL streptomycin. Activation of stimulator of interferon genes (STING) by treatment of HUVECs with cyclic guanosine monophosphate-adenosine monophosphate (cGAMP) was performed as described previously [22]. Activation of Toll-like receptor 9 (TLR9) by treatment of HUVECs with CpG-ODN-2006 (10 μM; Enzo Life Sciences) or GpC-ODN-2137 (10 μM; Enzo Life Sciences) was performed as described previously [23]. Recombinant His-tagged human DLL4 protein (R&D Systems) was resuspended at a concentration of 1 μg/mL in phosphate-buffered saline (PBS) containing 1% BSA. Tissue culture plates were incubated with DLL4 (1 μg/mL) or BSA at 4°C for 24 h. Plates were warmed to 37°C, and the DLL4 or BSA solutions were aspirated before seeding HUVECs (0.4 × 105/well in 12-well plates). The cells were treated with aptamers on the next day. The RNA and cell lysates were harvested at 30–96 h postseeding.
Transfection of plasmid, siRNA, and aptamer
For transfection of plasmids into HEK293T cells, GeneJuice (Millipore) was used according to the manufacturer's instructions. At 48 h post-transfection, the cells were processed for subsequent experiments. Transfections of siRNAs (10 nM) or aptamer (2 μM; Eurofins Genomics) into HUVECs were performed using RNAiMAX (Invitrogen) according to the manufacturer's instructions. Subsequent experiments were performed at 6 or 72 h post-transfection.
Lentiviral expression
Transient expression of Myc-CBF1 in HUVECs was induced through lentiviral infection as previously described [24]. The CSII-CMV-MCS-IRES2-Bsd, pCAG-HIVgp, and pCMV-VSVG-RSV-Rev vectors were kind gifts from Dr. Hiroyuki Miyoshi (RIKEN).
siRNAs
The following validated siRNA duplex oligomers were purchased and used for knockdown experiments: GAGCGGCCUCCACCUAAACGACUUA (siCBF1 #1, Invitrogen), GGGAAGCUAUGCGAAAUUAUUUAAA (siCBF1 #2, Invitrogen), and GAAAGCACUUCAUGUUGUCUGUAAA (siCBF1 #3, Invitrogen). Control siRNAs were purchased from Sigma (SIC-001).
Western blotting and immunoprecipitation
Western blotting and immunoprecipitation were performed as previously described [25]. The biotinylated proteins were detected using the VECTASTAIN ABC Kit (Funakoshi).
Pull-down assay
HUVECs were lysed in IP buffer (25 mM Tris-HCl pH 7.4, 150 mM NaCl, 1% NP-40, 1 mM EDTA, 5% glycerol) containing cOmplete protease inhibitors (Roche). After incubation of cell lysates on ice for 10 min, the cell lysates were centrifuged at 10,000 g for 10 min at 4°C. The resulting supernatants were incubated with 5′-terminally biotinylated aptamers (1 μM; Eurofins Genomics) for 2 h at 4°C followed by incubation with prewashed Dynabeads M280 Streptavidin (Invitrogen) for 1 h at 4°C. The beads were washed thrice with IP buffer.
Real-time polymerase chain reaction
Total RNAs were extracted from prostate cancer cells using ISOGEN II (Nippon Gene) according to the manufacturer's protocol. The total RNA (1 μg) was used for cDNA synthesis using High Capacity RNA-to-cDNA Master Mix (Applied Biosystems). RT-PCR was carried out (FastStart Universal SYBR Green Master ROX; Roche) on the ABI 7500 RT-PCR system (Applied Biosystems) using the following pairs of primers:
5′-TGAGTTCGGCTCTAGGTTCCA-3′ (HEY1 sense primer),
5′-GCGCTTCTCAATTATTCCTCTCC-3′ (HEY1 antisense primer),
5′-CAACCCCTTGTCGCCTCTC-3′ (HEY2 sense primer),
5′ CCGTGGATGGCATTCGGAG-3′ (HEY2 antisense primer),
5′-GCTGACTTATGCATTGCCTCAGGA-3′ (CBF1 sense primer),
5′-CCACTGCTGTGAACTGGCATGAAA-3′ (CBF1 antisense primer),
5′- GACCAACAAGTGTCTCCTCCAAA-3′ (IFNβ sense primer),
5′- AGCAAGTTGTAGCTCATGGAAAGAG -3′ (IFNβ antisense primer),
5′- AGGTGCAGTTTTGCCAAGGA-3′ (IL-8 sense primer),
5′- TTTCTGTGTTGGCGCAGTGT -3′ (IL-8 antisense primer),
5′-CATGAGAAGTATGACAACAGCCT-3′ (GAPDH sense primer),
5′-AGTCCTTCCACGATACCAAAGT-3′ (GAPDH antisense primer).
Immunofluorescence staining
Cells were fixed with 4% paraformaldehyde (PFA) in PBS for 30 min at room temperature and permeabilized with 0.1% Triton X-100 in PBS for 15 min at room temperature. For staining of endogenous CBF1, cells were fixed with 10% trichloroacetic acid (TCA) in PBS for 15 min at 4°C and permeabilized with 0.05% saponin in PBS for 5 min at room temperature. After blocking with 3% BSA in PBS for 30 min at room temperature, the cells were incubated with primary antibodies and then with secondary antibodies conjugated to fluorophores. To stain nuclei, the fixed cells were treated with Hoechst 33342 (dilution 1:2,000; Molecular Probe) at room temperature for 1 h. The 5′-terminally Cy3-labeled aptamers were purchased from Eurofins Genomics.
Tube formation assay
Aptamers were introduced into HUVECs (0.4 × 104 cells/well of 96 well plates) seeded on the reconstituted Type I-A collagen gel (Nitta Gelatin). At 6 h postaptamer delivery, the cells were packed with Type I-A collagen gel followed by the addition of VEGF-A (50 ng/mL: R&D Systems) in 0.15% serum-containing EBM-2 medium. At 66 h postaddition of VEGF-A, cells were incubated with Calcein-AM (DOJINDO) for 1 h, and then the tube area was measured with FIJI software (NIH). HUVECs were treated with bevacizumab (0.125 μg/mL: MedChemExpress) for 66 h before the addition of Calcein-AM.
Water-soluble tetrazolium salt-1 assay
HUVECs were seeded in 96-well plates (0.4 × 104 cells/well), and the cells were treated with aptamers on the next day. After 66 h, 10 μL of Cell Proliferation Reagent WST-1 (Sigma) was added to the growth medium. The plates were incubated for 30 min, and the absorbance at 420 nm was measured.
Confocal microscopy
Confocal microscopy was performed using the A1R laser confocal microscope (Nikon) with 60 × 1.27 Plan-Apochromat water immersion lens. Images were analyzed with FIJI (NIH). For quantitation of tube formation assay, the image thresholding for its binarization was performed using FIJI (NIH).
Results
Selection of ssDNA aptamers for recombinant CBF1 protein by SELEX
SELEX is a well-established method for the selection of aptamers [26]. We thus used this method to develop CBF1-bound ssDNA aptamers (Fig. 1). We prepared an initial pool of ssDNAs containing a central random sequence of 30 nucleotides, flanked by specific sequences of 20 nucleotides at the 5′ and 3′ termini. The negative selection was performed by incubating the initial ssDNA pool (2.8 × 1015 molecules) with streptavidin-coated beads (SAv beads), followed by supernatant collection. The obtained supernatant was incubated with biotinylated recombinant human CBF1 protein (Biotin-CBF1)-bound SAv beads (Supplementary Fig. S1). The biotinylated CBF1 protein was efficiently captured from the wheat germ extract translation system (Supplementary Fig. S1). After washing, ssDNA-CBF1 complexes on SAv beads were denatured by heating in the presence of sodium dodecyl sulfate (SDS), reducing reagents, and urea. The eluted ssDNA was then amplified by PCR. We used a 5′ phosphorylated reverse primer because the amplified 5′ phosphorylated antisense strands could be digested by treatment with λ exonuclease. The obtained ssDNA aptamer pool was used in the next round of negative selection. The selection cycle was repeated 14 times with gradually decreasing concentrations of both Biotin-CBF1 and ssDNA to increase the stringency of interaction between CBF1 and ssDNA pools (Table 1). The number of the PCR cycle and volume of the PCR template were also decreased gradually to minimize the amplification bias (Table 1). After the 14th round, the PCR products were subjected to sequencing with a next-generation sequencer. Of the 198,955 read sequences, 17,140 were chosen for clustering into several groups with the same sequence. The abundance of each aptamer in 14th pool was calculated as a percentage of each number of reads in the 17,140 reads (Table 2). The 15 most abundant ssDNA aptamers that bound to recombinant human CBF1 protein were selected (Apt-1 to Apt-15; Table 2). As shown in Table 2 and Supplementary Fig. S2, the developed ssDNA aptamers bound to CBF1 with high affinity (Kd ≈ 10–300 nM). The Kd of selected 15 aptamers (Apt-1 to Apt-15) did not correlate to their abundance (Table 2). It is likely that a variety of factors (eg, existence of other ssDNA and amplification bias) during SELEX affected the abundance of low-affinity aptamers.
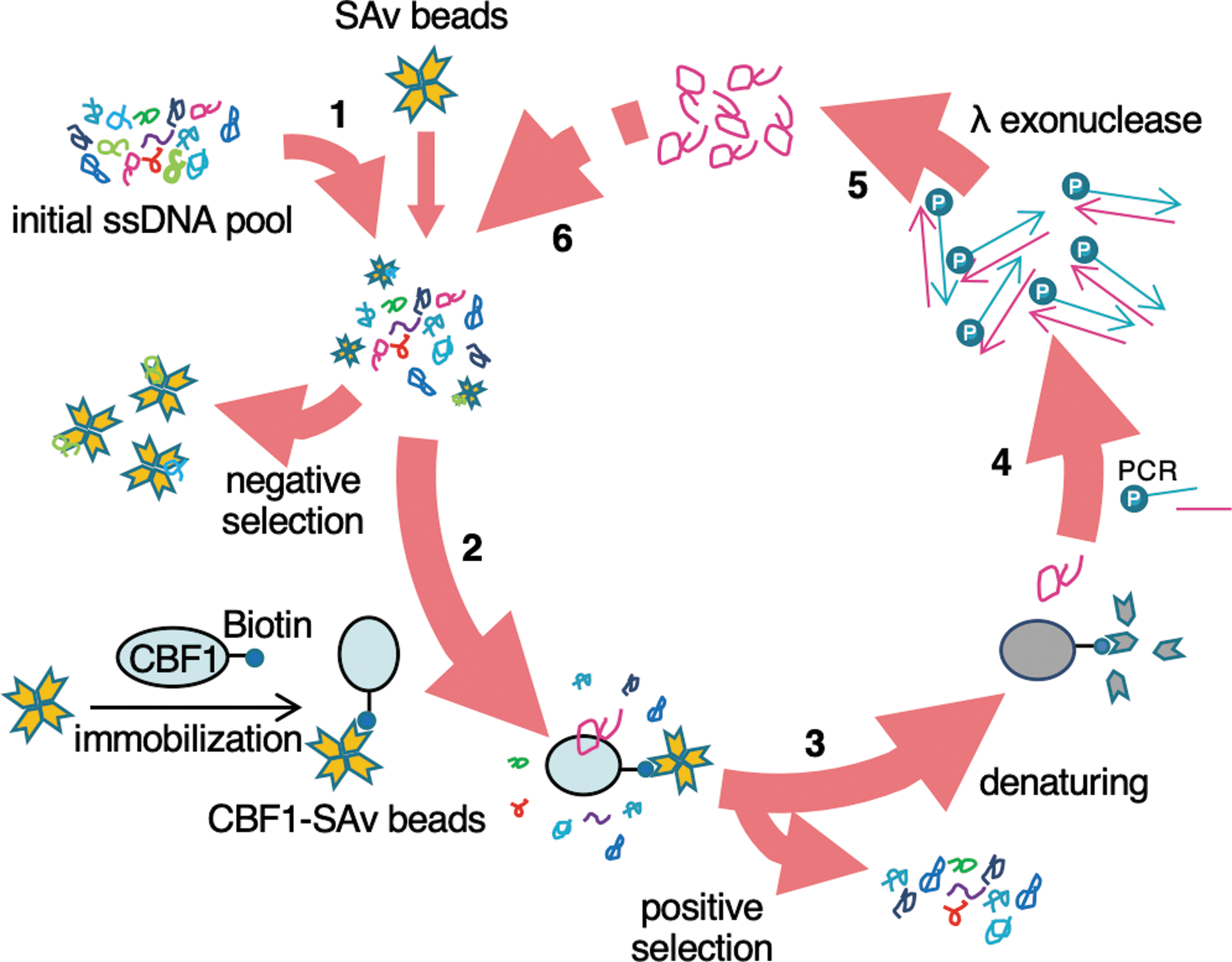
In vitro selection of CBF1-targeting single-stranded DNA aptamers. The scheme of SELEX in this study is shown. A total of 4.6 nmol (2.8 × 1015 molecules) of ssDNA pool was screened in the first step. The selection cycle was repeated 14 times. SAv: streptavidin. See also Materials and Methods section. 1, 6: Negative selection of ssDNAs that bind to SAv beads. 2: Positive selection of ssDNAs that bind to CBF1-immobilized SAv beads. 3: Denaturation and purification of the CBF1-bound ssDNAs. 4: Amplification of the purified CBF1-bound ssDNAs by PCR. 5: Treatment of the PCR products with λ exonuclease followed by purification to obtain an enriched ssDNA pool for the next round. CBF1, C promoter binding factor 1; PCR, polymerase chain reaction; SELEX, systematic evolution of ligands by the exponential enrichment; ssDNA, single-stranded deoxyribonucleic acid.
We next compared the sequences and secondary structures of obtained aptamers. As shown in Supplementary Fig. S3, most of the obtained 15 aptamers can be classified into three groups (Apt-1, 3, 6; Apt-14, 15; Apt-2, 7, 8, 9, 10, 12) with high homology within each group. Apt-4, 5, 11, and 13 do not possess similar sequences compared to other aptamers (Supplementary Fig. S3). The RNAstructure web server [27], which is also available for ssDNA secondary structure prediction, predicted that all the CBF1-bound ssDNA aptamers (Apt-1 to Apt-15) form secondary structures (Supplementary Figs. S4–S8). These data suggested that the aptamers bind to CBF1 in a shape-dependent manner. Within a group containing Apt-1, 3, 6, we noticed that Apt-1 does not contain T in a stem-loop at the central region, which would reduce the stability of the stem-loop compared to that of Apt-3 and Apt-6 (Supplementary Figs. S3 and S4). The lower affinity of Apt-1 to CBF1 compared with Apt-3 and Apt-6 may be accounted for this structural difference of the stem-loop.
We then verified intracellular introduction of aptamers to human umbilical vein endothelial cells (HUVECs). To this aim, we used Cy3-labeled Apt-1 as a representative. As shown in Supplementary Fig. S9, Cy3-labeled Apt-1 was introduced into HUVECs using RNAiMAX transfection reagents. The aptamers were well colocalized with CBF1 in intracellular puncta and nuclei (Supplementary Fig. S9). It is likely that the Apt-1/CBF1 complex was transported into nuclei according to the nuclear localization signals of CBF1. In contrast, Cy3-labeled negative control-dA30 (NC-dA30) aptamers were localized at intracellular puncta without colocalization with CBF1 at nuclei in HUVECs (Supplementary Fig. S9). These data suggested that Apt-1 selectively interacted with CBF1 at nuclei.
Effect of CBF1-targeting ssDNA aptamers on in vitro angiogenesis
To evaluate the effects of the selected CBF1-binding ssDNA aptamers (Apt-1 to Apt-15) in human endothelial cells, we performed a tube formation assay, mimicking angiogenesis in vitro [28]. As shown in Fig. 2A and B, treatment of HUVECs with Apt-1, 3, 4, 5, or 6 significantly reduced the tube area to ∼30%–50% compared to controls. The tube area was decreased by ∼70% compared to controls by treatment of HUVECs with a clinically-used angiogenic inhibitor, bevacizumab (a humanized anti-VEGF monoclonal antibody) (Fig. 2A, B). Other CBF1-binding ssDNA aptamers that exhibited no antiangiogenic activities served as additional negative controls. These data suggested that Apt-1, 3, 4, 5, and 6 possessed antiangiogenic effects. The water-soluble tetrazolium salt (WST)-1 assay showed that treatment of HUVECs with Apt-3 significantly inhibited cell proliferation, whereas other tested aptamers did not affect endothelial cell proliferation (Fig. 2C). Although Apt-2, 7, 8, 9, 10, 11, 12, 13, 14, and 15 can interact with recombinant CBF1 protein (Table 2), they affected neither tube formation nor cell proliferation of HUVECs (Fig. 2A–C), suggesting that binding of those aptamers to CBF1 did not affect its molecular functions in cells. From the aptamer-structural standpoint, although Apt-14, 15 partially possess the same sequences as those of Apt-1, 3, 6, the predicted secondary structures of Apt-14, 15 are clearly different from those of Apt-1, 3, 6 (Supplementary Figs. S3–S5). These observations suggested that the conserved sequence of Apt-1, 3, 6 together with their specific secondary structure might be critical for the antiangiogenic activities. We then focused on the five CBF1-targeting ssDNA aptamers (Apt-1, 3, 4, 5, and 6) that inhibited angiogenesis in vitro.
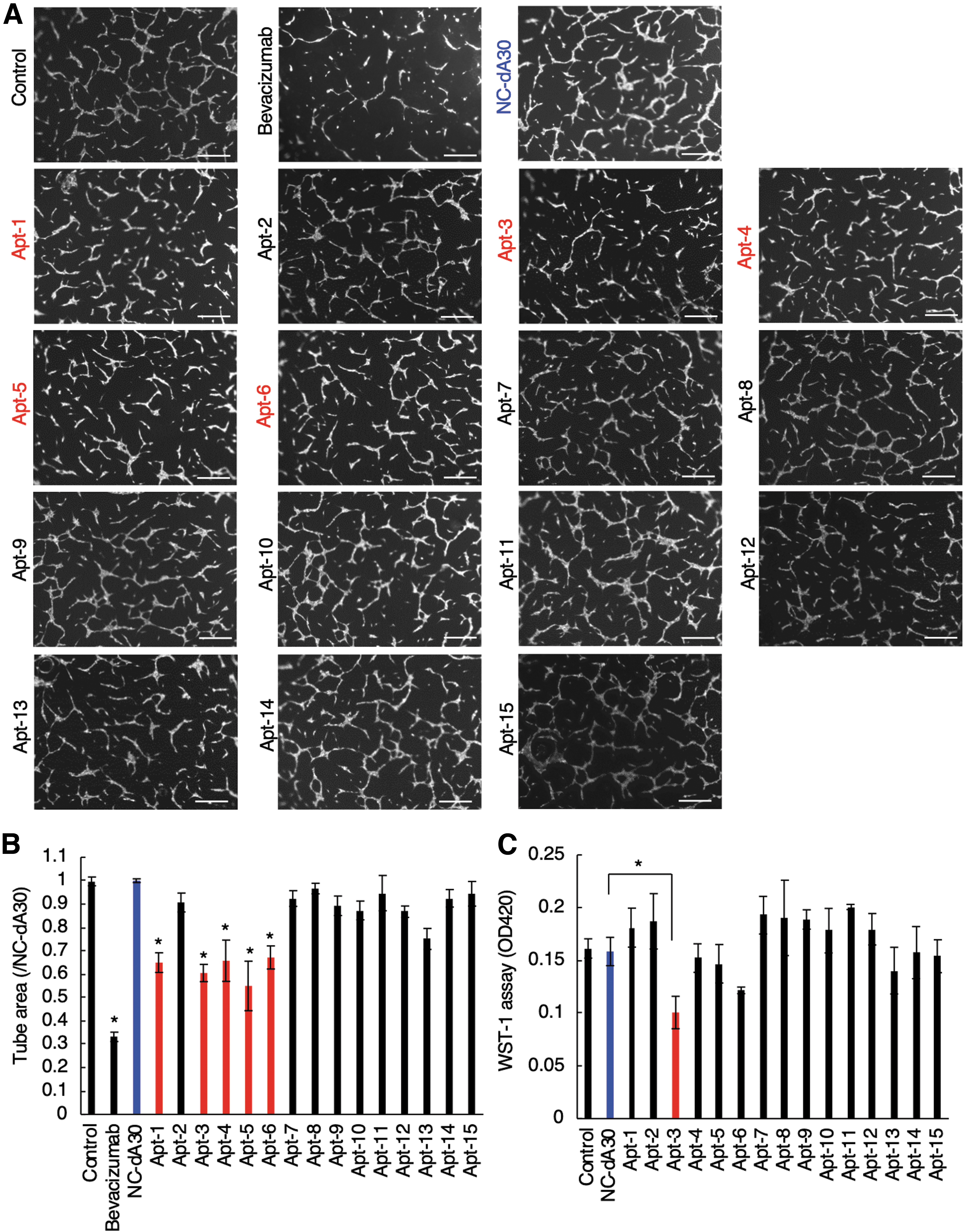
Effects of CBF1-targeting ssDNA aptamers on tube formation and proliferation of HUVECs.
The cyclic GMP-AMP synthase (cGAS)/STING pathway is necessary for type I interferon response against pathogen-derived intracellular ssDNA [29]. The recognition of CpG DNA derived from pathogens by TLR9 also induced immune responses [30]. To examine if the antiangiogenic CBF1-targeting ssDNA aptamers (Apt-1, 3, 4, 5, and 6) activated the cGAS/STING and CpG/TLR9 pathways in HUVECs, we examined type I interferon and interleukin responses, respectively. We first confirmed that the treatment with cyclic GMP-AMP (cGAMP) to HUVECs drastically increased the level of interferon β (IFNβ) mRNA, indicating the existence of a functional cGAS/STING pathway in HUVECs (Supplementary Fig. S10A). The stimulation of HUVECs with CpG-ODN-2006 upregulated the mRNA level of IL-8 as reported previously [23] (Supplementary Fig. S10B). As shown in Supplementary Fig. S10A and B, Apt-1, 3, 4, 5, and 6 affected the level of neither IFNβ nor IL-8 mRNA. These data indicated that Apt-1, 3, 4, 5, or 6 activated the neither cGAS/STING nor CpG/TLR9 pathway in endothelial cells.
A CBF1-targeting-ssDNA aptamer (Apt-3) upregulates HEY2 gene expression
Since Apt-1, 3, 4, 5, and 6 inhibited tube formation in HUVECs (Fig. 2A, B) and previous studies reported that CBF1-dependent Notch signaling suppresses angiogenesis [11,13], we speculated that the above CBF1-targeting aptamers would enhance CBF1 activity. As shown in Fig. 3A–C, neither protein nor mRNA levels of CBF1 were affected by treatment of HUVECs with antiangiogenic aptamers (Apt-1, 3, 4, 5, and 6). Nuclear localization of endogenous CBF1 was detected in aptamer-treated HUVECs, as well as in negative control NC-dA30 aptamer-treated HUVECs (Fig. 3D). These data suggest that Apt-1, 3, 4, 5, and 6 could enhance the activities of CBF1 without affecting its expression and cellular localization. A Notch ligand, DLL4, interacts with a Notch receptor, Notch1, on endothelial surface followed by sequential shedding of Notch1 leading to the release of Notch intracellular domain (NICD) into the cytosol [10]. NICD then translocates to the nucleus and forms a complex with CBF1 on promoter region of targeted genes resulting in the activation of their transcriptions [10]. We thus examined the effects of antiangiogenic aptamers (Apt-1, 3, 4, 5, and 6) on the regulation of CBF1-targeting genes, HEY1 and HEY2. Knockdown efficiency of CBF1 was confirmed in HUVECs (Fig. 3E). The expression level of HEY1 and HEY2 mRNA was elevated by ∼10 and 300 times, respectively, by the DLL4 stimulation, which was drastically decreased by CBF1 knockdown in HUVECs (Fig. 3F, G). These data indicated the CBF1 dependency on upregulation of HEY1 and HEY2 mRNA in the presence of DLL4 in HUVECs. Experiments with biological replicates showed that, among the tested antiangiogenic aptamers, treatment with Apt-3 slightly increased the level of HEY1 mRNA and significantly increased that of HEY2 mRNA in the presence of DLL4 stimulation (Fig. 3H, I). Together with the upregulation of HEY2 mRNA, the protein expression of HEY2 in DLL4-stimulated HUVECs was increased approximately threefold by treatment with Apt-3 compared to control aptamers (Fig. 3J). These data suggested that Apt-3 could activate CBF1. In contrast, the significant upregulation of HEY1 and HEY2 mRNA was not observed in HUVECs treated with Apt-1, 4, 5, 6 (Fig. 3H, I). Those aptamers may inhibit in vitro angiogenesis through the HEY1/HEY2-independent pathways that is not critical for cell proliferation. The only difference between Apt-3 and Apt-6 is one cytosine residue at the first position of the random region (Supplementary Figs. S3 and S4), suggesting critical roles of the specific single-stranded region of Apt-3 in upregulation of HEY2 mRNA. Because activation of Notch signaling suppresses cell growth [31], increased expression of HEY2 mRNA by Apt-3 may account for the inhibition of endothelial cell proliferation by treatment with Apt-3 (Fig. 2C). The interaction between biotinylated Apt-3 and endogenously-expressing CBF1 was detected in HUVECs (Fig. 3K). As shown in Fig. 3L, we confirmed the dose dependency of Apt-3 in inhibitory efficacy of angiogenesis in HUVECs (IC50 = 0.025 μM). SPR analysis indicated that Apt-3 directly interacted with CBF1 at a Kd value of 39 ± 1.2 nM (Fig. 4 and Supplementary Fig. S11). The value was close to a Kd value calculated by the binding curve based on PCR (34 nM, Table 2), suggesting the reliability of Kd values shown in Table 2. Collectively, these data suggested that Apt-3 binding to CBF1 resulted in CBF1 activation in human endothelial cells.
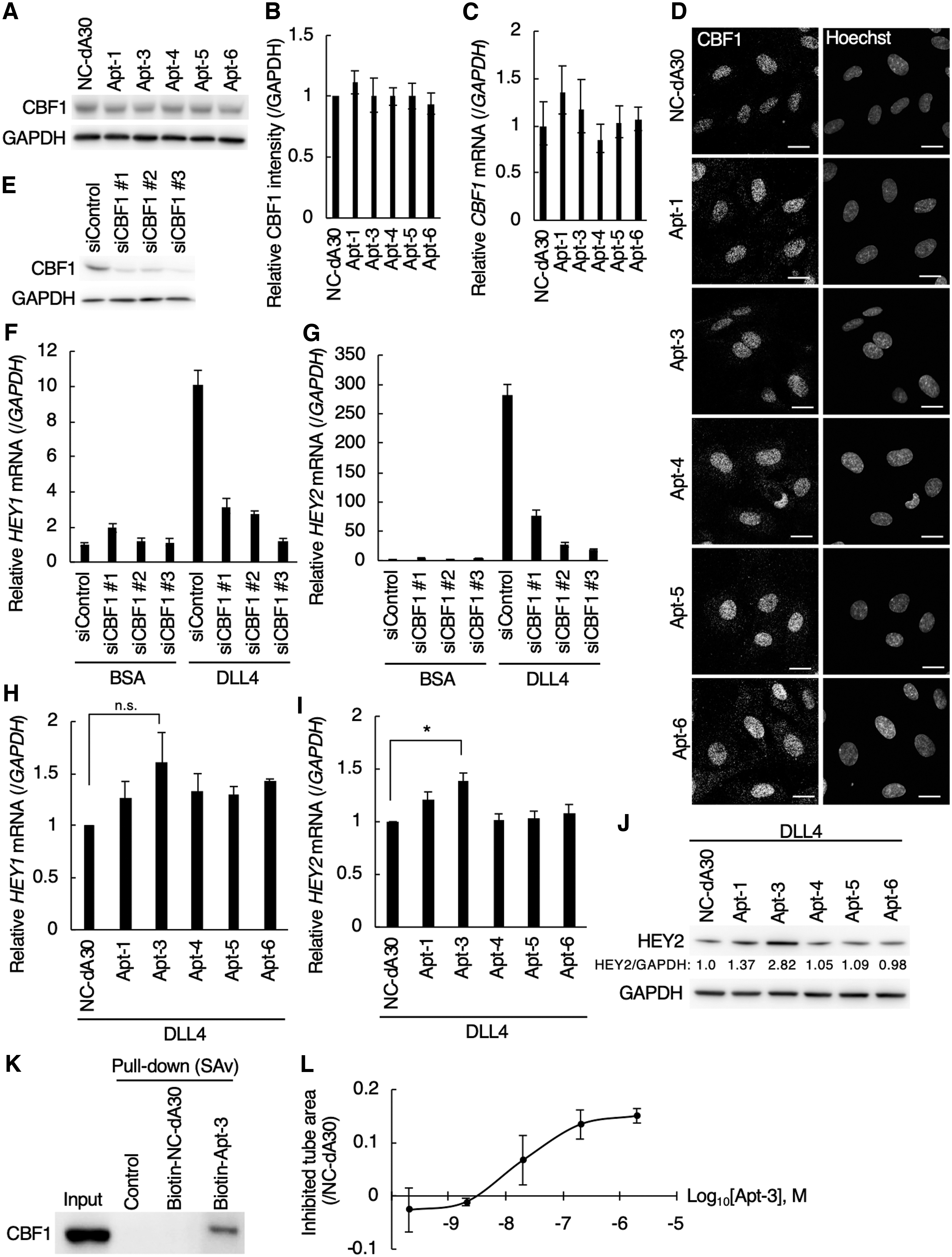
Site of action of the CBF1-targeting ssDNA aptamers (Apt-1, 3, 4, 5, 6).
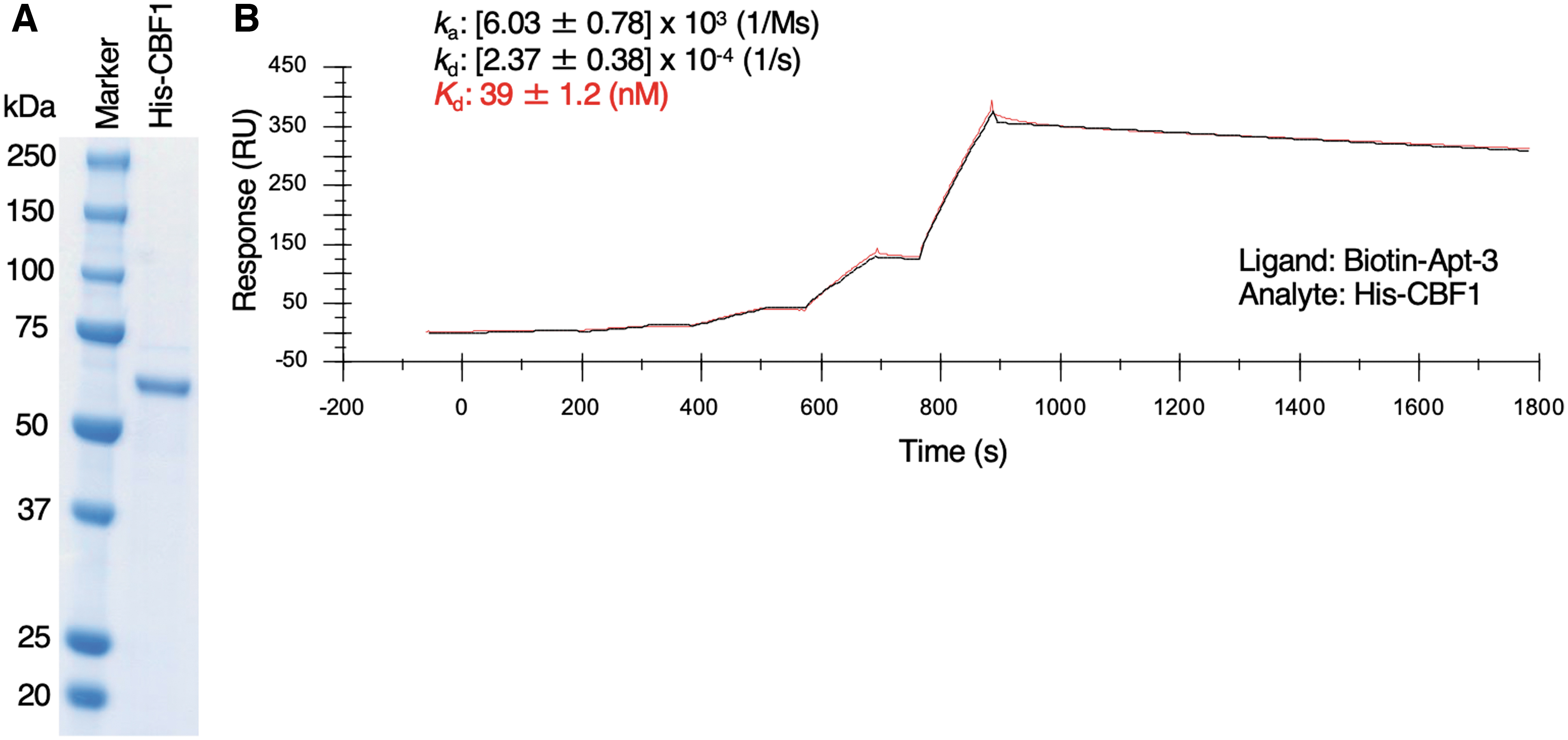
SPR single-cycle kinetics of interaction between Apt-3 and CBF1.
Apt-3 targets the C-terminal region of the LAG1 domain of CBF1
We finally sought to identify the CBF1 domain(s) interacting with Apt-3. The Pfam protein database [32] indicated that CBF1 possesses a LAG1 domain (Lin-12 and Glp-1 phenotype domain) located at 48th–178th aa (amino acid) and a BTD domain (β-trefoil DNA-binding domain) located at 179th–328th aa in its amino acid sequence (Fig. 5A). Recombinant FLAG-tagged CBF1 proteins (full length or domain deleted) were produced using wheat germ extract [33], as shown in Fig. 5B, and then subjected to cell-free based AlphaScreen assay [34] to verify their direct interaction with biotinylated Apt-3. Enhanced AlphaScreen signal confirmed the interaction between CBF1 (full length and BTD domain deleted) and Apt-3 (Fig. 5C). In contrast, deletion of the LAG1 domain completely abolished this interaction (Fig. 5C). Enhanced AlphaScreen signal was also detected between recombinant LAG1 domain and Apt-3 (Fig. 5B, C). These data suggested that Apt-3 directly interacted with the LAG1 domain of CBF1. We next produced six truncated CBF1 mutants, each possessing 21 or 22 amino acids of the LAG1 domain without overlapping each other (Fig. 5A, D). Of these, mutant 6 interacted with Apt-3 with efficiency comparable to that of full length CBF1 (Fig. 5E). Collectively, these data suggested that Apt-3 directly and specifically targeted a sequence spanning residues from 158 to 178 (SKRIKVISKPSKKKQSLKNAD) in the LAG1 domain of CBF1.

LAG1 domain of CBF1 interacts with Apt-3.
Discussion
CBF1 is a critical transcription factor in Notch signaling and, therefore, a promising therapeutic target [35]. To date, a CBF1 inhibitor, termed RBPJ INhibitor-1 (RIN1), has been developed [36]. RIN1 is a small chemical compound that inhibits the interaction between CBF1 and NICD, suppressing the growth of hematologic cancer cells and promoting skeletal muscle differentiation [36]. In this study, we developed a CBF1-binding ssDNA aptamer, Apt-3, which inhibited angiogenesis of HUVECs. Notably, treatment of HUVECs with Apt-3 increased HEY1 and HEY2 expression in the presence of DLL4 stimulation. These data suggested that Apt-3 was a CBF1 activator and an inhibitor of angiogenesis. The appropriate type of CBF1 modulation, that is, inhibition (by RIN1) or stimulation (by Apt-3) will depend on the clinical context. From the clinical standpoint, the inability of Apt-3 to activate the cGAS/STING and CpG/TLR9 pathways is beneficial (Supplementary Fig. S10), as it avoids the risk of side effects due to type I interferon responses such as those related to autoimmune disorders. The current main limitation to the therapeutic application of aptamers is related to their delivery to target molecules in cells and tissues [2]. Notably, while pegaptanib, an RNA aptamer that is approved for AMD, interacts with extracellular VEGF and can easily reach its target, Apt-3 interacts with CBF1 in nuclei. Strategies promoting Apt-3 delivery to the cell nucleus, such as the use of lipid carriers, will have to be developed to allow for efficient delivery of this molecule to its target, CBF1.
The double-stranded DNA sequence that interacts with CBF1 is 5′-CGTGGGAA-3′/5′-TTCCCACG-3′ [37]. We noticed that Apt-11 and Apt-13 possess GTGGGAA and TTCCCACG, respectively, which are likely to be partially located in double-stranded regions (Supplementary Figs. S3 and S8). Although Apt-5 possesses TTCCCA in its sequence, the region seems not to form a double-strand structure (Supplementary Fig. S8). Other aptamers do not possess the CBF1-bound double stranded DNA sequence (Supplementary Fig. S3). Together, these predictions suggest that antiangiogenic aptamers (Apt-1, 3, 4, 5, 6) would not at least hamper the promoter recognition of CBF1.
X-ray crystallography has revealed that CBF1 contains a conserved structural core that is largely composed of β strands, located in the LAG1 and BTD domains [38]. The BTD domain interacts with various crucial coregulators of CBF1 (eg, NICD [39], RITA1 [40], SPEN [41]). Another component of the CBF1 protein complex, MAML1, interacts with the LAG1 domain of CBF1, leading to CBF1-dependent gene expression [42]. In this study, we found that the CBF1-targeting ssDNA aptamer, Apt-3, interacted with the C-terminal region of the LAG1 domain (Fig. 5E). The CBF1 region found to interact with Apt-3 (amino acid residues 158–178) forms a β-strand that establishes contacts with the DNA chains [43]. The binding of Apt-3 to the C-terminal β-strand in the LAG1 domain may cause conformational changes, which promote CBF1 interaction with its target DNA sequences.
Footnotes
Acknowledgments
The authors thank Ms. Mami Chosei, Ms. Yuki Tanaka, and Drs. Shinji Fukuda and Tomohisa Sakaue (Ehime University) for providing their technical assistance, Drs. Kojiro Mukai and Tomohiko Taguchi (Tohoku University) for providing reagents, and Dr. Takao Inoue (National Institute of Health Sciences) and Mr. Kota Tange (NOF Corporation) for their helpful information.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by JSPS KAKENHI grant no. 18K15718 to M.T-.K., JSPS KAKENHI grant no. 18K15244 to M.M., Takeda Science Foundation to Proteo-Science Center, and JSPS KAKENHI grant no. 19K22564 to A.O. and S.H.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.