
Abstract
Phosphorothioate antisense oligonucleotides (PS-ASOs) interact with proteins and can localize to or induce the formation of a variety of subcellular PS-ASO-protein or PS-ASO-ribonucleoprotein aggregates. In this study, we show that these different aggregates that form with varying compositions at various concentrations in the cytosol, nucleus, and nucleolus may undergo phase separations in cells. Some aggregates can form with both nontoxic and toxic PS-ASOs, such as PS bodies, paraspeckles, and nuclear filaments. However, toxic PS-ASOs have been shown to form unique nucleolar aggregates that result in nucleolar dysfunction and apoptosis. These include liquid-like aggregates that we labeled “cloudy nucleoli” and solid-like perinucleolar filaments. Toxic nucleolar aggregates may undergo solid-phase separation and in the solid phase, protein mobility in and out of the aggregates is limited. Other aggregates appear to undergo liquid-phase separation, including paraspeckles and perinucleolar caps, in which protein mobility is negatively correlated with the binding affinity of the proteins to PS-ASOs. However, PS bodies and nuclear filaments are solid-like aggregates. Importantly, in cells that survived treatment with toxic PS-ASOs, solid-like PS-ASO aggregates accumulated, especially Hsc70-containing nucleolus-like structures, in which modest pre-rRNA transcriptional activity was retained and appeared to mitigate the nucleolar toxicity. This is the first demonstration that exogenous drugs, PS-ASOs, can form aggregates that undergo phase separations and that solid-phase separation of toxic PS-ASO-induced nucleolar aggregates is cytoprotective.
Introduction
Inside cells, DNA, RNA, and proteins are organized spatially into distinct structures that facilitate various functions. Such separation is achieved in membraned organelles (eg, mitochondria, endoplasmic reticulum, and nucleus) and in membrane-less structures. Many membrane-less structures are well defined, including nucleolus, Cajal bodies, paraspeckles in the nucleus, and processing bodies (P-bodies) and stress granules in the cytoplasm. Membrane-less structures can form due to liquid-liquid phase separation (LLPS) or liquid-solid phase separation (LSPS) to generate distinct structures with concentrated components relative to the surrounding environments [1,2]. Although phase separation is required for normal biological processes, abnormal phase separation can be associated with disease [3].
Phase separation can be initiated by interactions of proteins with RNA or DNA as what occurs in the nucleolus, a site required for pre-rRNA transcription and processing [4,5], and during the formation of paraspeckles, which contain multiple RNA-binding proteins (eg, P54nrb, PSF, PSPC1, and FUS) and the long noncoding RNA NEAT1 [6,7]. For liquid-like structures, the composition and physical properties vary depending on cellular factors and environmental conditions. In cells, formation of these membrane-less structures is tightly regulated [8,9], and the structures are dynamic [1,2,10]. In certain cases, LSPS can occur. For example, the nucleolus has been shown to undergo LSPS upon prolonged heat shock, and to serve as a site for protein quality control [11,12]. Although phase separation in certain cellular organelles has been well characterized, such as LLPS of cytoplasmic P bodies, nuclear paraspeckles, and the nucleolus [4,13], little is known regarding the phase separation that occurs in cellular structures that are induced by or contain exogenous molecules such as antisense oligonucleotide (ASO) drugs.
ASOs can modulate gene expression through different posthybridization mechanisms [14–16]. For example, ASOs can reduce target RNA levels by directing RNase H1 cleavage or by triggering nonsense-mediated decay or no-go decay [17–19]. Protein levels can also be increased by using ASOs that modulate splicing or that target translation inhibitory elements [15,20–23]. The mechanism of action of an ASO depends on the chemical modifications and the binding sites in target RNAs. RNase H1-dependent ASOs, which are the focus of this study, are “gapmers” with 8 to 10 deoxynucleotides in the center to enable RNase H1 activation and 3 to 5 nucleotides at the ends modified with different 2′-modifications, such as 2′-methoxyethyl (2′-MOE), constrained ethyl (cEt), 2′-fluoro (2′-F), and locked nucleic acids (LNA) [24–27]. To enhance stability, tissue and cellular uptake, and pharmacological properties, ASOs are also modified with phosphorothioate (PS) backbone linkages.
Chemical modifications of ASOs not only affect ASO-RNA hybridization but also influence ASO-protein interactions. PS-modified ASOs (PS-ASOs) bind more tightly and more avidly to proteins than do ASOs with a phosphodiester backbone [28,29]. In addition, 2′-modifications with more hydrophobicity (eg, 2′-F, cEt, and LNA) tend to significantly increase protein binding affinity compared with more hydrophilic 2′-modifications (eg, 2′-MOE) [30–32]. Many proteins that bind to PS-ASOs are DNA- or RNA-binding proteins; some are chaperone proteins, including TCP1 complex and heat shock proteins [28,31,33–35]. Interactions with proteins can influence PS-ASO potency through effects on uptake and subcellular trafficking and distribution [28,36–40]. PS-ASO binding can also affect the fates of proteins such as their localization, stability, or interaction with other partners [30,41,42]. Thus, interactions with proteins can influence safety of PS-ASO therapeutics. Indeed, we have recently shown that most toxic PS-ASOs tend to bind more proteins more tightly than nontoxic PS-ASOs, leading to nucleolar mislocalization of paraspeckle proteins and nucleolar stress, resulting in apoptotic cell death [43–45].
When cells are transfected with high doses of PS-ASOs, toxic or nontoxic, the PS-ASOs can localize to different cellular structures, for example, cytoplasmic P-bodies, stress granules, and nuclear paraspeckles [34,41,42]. PS-ASOs are observed in these structures shortly after transfection (Fig. 1A), and localization is dependent on interactions with proteins [29,33,34,41,42]. After 1 to 4 h, PS-ASOs can induce the formation of distinct structures, such as nuclear PS bodies that contain chaperone protein TCP1β, a protein normally localized in the cytoplasm [33], and nuclear PS-ASO filaments that contain paraspeckle proteins, including PSF and P54nrb [34]. However, some structures are formed only with toxic PS-ASOs, for example, mislocalization of paraspeckle proteins to the perinucleolar cap and the nucleolus that is observed shortly after transfection of nearly all toxic PS-ASOs tested (Fig. 1A) [43]. The various PS-ASO-containing structures have distinct morphologies and protein components (Fig. 1A). However, the nature of phase separation in these various PS-ASO-containing structures has not been studied. As PS-ASO binding to cellular proteins may alter protein conformation or binding partners, especially for the paraspeckle proteins that contain low-complexity domains and bind tightly with PS-ASOs [32], it is possible that PS-ASOs alter the phase separation of proteins in PS-ASO-containing cellular structures.
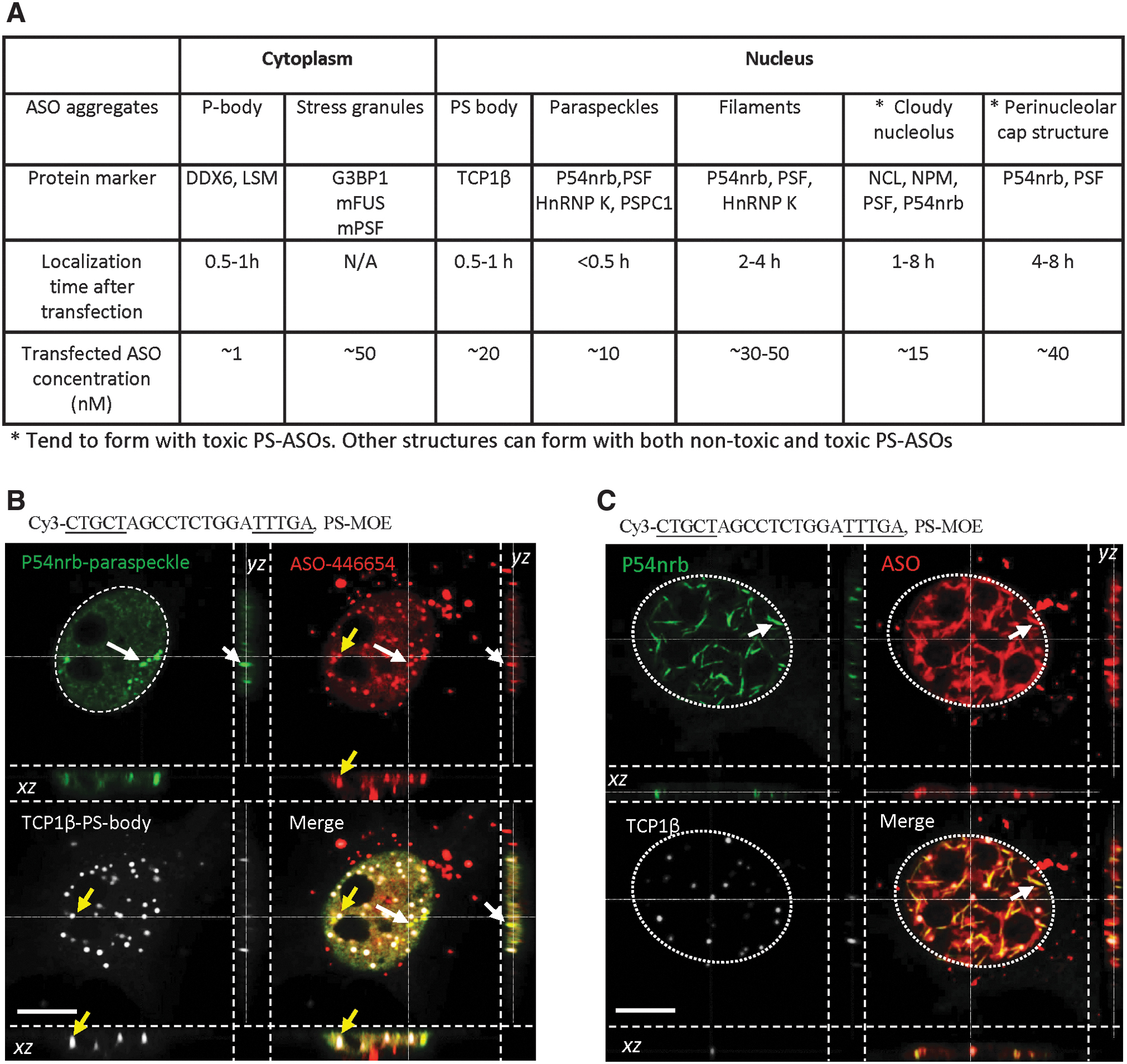
PS-ASOs form different subcellular structures after transfection into cells.
In this study, we characterized, for the first time, the phase separation of different PS-ASO-containing structures in cells, using 1,6 hexanediol-mediated disruption and fluorescent recovery after photobleaching (FRAP) approaches. PS-ASO-containing cytoplasmic P-bodies, nuclear paraspeckles, toxic PS-ASO-induced perinucleolar caps, and cloudy nucleoli were found to be liquid like. PS bodies, nuclear PS-ASO filaments, and PS-ASO-containing cytoplasmic granules formed with mutant FUS and mutant PSF proteins appear to be solid-like aggregates. In addition, we found that 6 to 24 h after transfection, toxic PS-ASOs tend to localize to perinucleolar filaments and nucleolus-like structures, which are solid-like aggregates. Interestingly, in cells that survived toxic PS-ASO treatment, we observed enrichment in solid-like PS-ASO structures, especially nucleolus-like structures, suggesting that the formation of solid-like structures sequesters toxic PS-ASOs to allow cells to survive.
Materials and Methods
Phosphorothioate ASO
PS-ASOs or siRNAs used in this study are listed in Supplementary Table S1. PS-ASOs were synthesized in-house using standard protocols [46]. siRNA was purchased from IDT.
Plasmids
Wild-type GFP-PSF plasmid was purchased from Genescript (OHu23607C), GFP-FUS, GFP-FUS-P525L, and GFP-ΔNLS-PSF were described previously [41]. GFP-NPM1 (RG203344) and GFP-P54nrb (RG226567) were purchased from Origene. GFP-DDX6 was described previously [42].
Antibodies
Antibodies against TCP1β (ab92746), FUS (ab23439), G3BP1 (ab56574), GAPDH (ab8245), Lamp1 (ab25630), NPM1 (ab24412), NCL (ab13541), NOP58 (ab155969), 5-bromouridine (BrU; ab6326), HSP90 (ab53497), and γH2AX (ab26350) were from Abcam. Antibodies against P54nrb (sc-376865), PSF (sc-374502), and RPA194 (sc-46699) were purchased from Santa Cruz Biotechnology. Anti-HSP70 antibody was from Enzo (ADI-SPA-820-F). Anti-R-loop antibody was from Kerafast (ENH001). RNase H1 and PS-ASO antibodies were raised at home and described previously [33,47]. Anti-rabbit secondary antibodies conjugated to AF488 (ab150077) and AF647 (ab150079) and anti-mouse secondary antibodies conjugated to AF488 (ab150113) and AF647 (ab150115) were purchased from Abcam.
Cell culture and transfection
HeLa and MHT cells were grown in DMEM supplemented with 10% FBS at 37°C in an incubator with 5% or 8% CO2. One day before transfection, cells were seeded at ∼70% confluency and incubated overnight. Transfection of PS-ASOs was performed in Opti medium using 4 μL/mL Oligofectamine 2000 (Life Technologies), based on manufacturer's instructions. Medium was replaced with prewarmed complete DMEM 4 h after transfection. For live-cell imaging, the medium was replaced immediately before imaging with prewarmed FluoroBrite DMEM live-cell imaging medium or Opti-medium (Life Technologies). Transfection of plasmids (1.5 μg plasmid per 10-cm dish) was performed using Oligofectamine 3000 based on manufacturer's instructions. At 24 h after plasmid transfection, cells were reseeded into glass-bottom dishes and were incubated overnight before PS-ASO treatment or imaging.
For reduction of ras-related nuclear protein (RAN), the siRNA targeting RAN (Santa Cruz Biotechnology, SC-36382) was transfected into HeLa cells at 5 nM final concentration for 36 h, and cells were reseeded in glass-bottom dishes and incubated overnight. Cells were then transfected with PS-ASO and, after 6 h, were subjected to immunofluorescent staining and imaging. Reduction of RAN mRNA was confirmed by quantitative real-time polymerase chain reaction using TaqMan primer probe sets described previously [33].
Drug treatment
For live-cell imaging, cells in glass-bottom dishes were placed under the confocal microscope (Olympus) and 10% 1,6-hexanediol (1,6-HD; Sigma) was added, followed by imaging. For immunofluorescent staining, cells were treated with 10% 1,6-HD for different times at 37°C in Opti medium before live-cell imaging or immunofluorescent staining. For 5,6-dichlorobenzimidazole 1-β-D-ribofuranoside (DRB; Sigma) or actinomycin D (ActD; Sigma) treatment, cells were incubated with 100 μg/mL DRB or 1 μg/mL ActD at 37°C for 45 min before imaging or subsequent treatments.
Cell viability assay
HeLa cells were either mock treated or transfected with 75 nM toxic PS-ASO (ASO ID 464917) for different times. Cells were then incubated with 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt (WST-8; Sigma) at 37°C in 8% CO2 incubator for 1 h. The percentage of viable cells was determined based on the ratio of absorbance at 450 nm to that at 600 nm as measured using a TECAN infinite M200 plate reader. Biological triplicates were analyzed.
Immunofluorescent staining and confocal imaging
Cells grown in glass-bottom dishes were treated with or without PS-ASOs or different drugs before immunofluorescent staining. For nascent RNA labeling, cells were incubated with 1 mM BrU (Sigma) at 37°C for 30 min before staining. Immunofluorescent staining was performed essentially as described previously [33]. Briefly, cells were washed with 1 × phosphate-buffered saline (PBS), fixed with 4% paraformaldehyde at room temperature (RT) for 30 min, permeabilized using 0.1% Triton in PBS at RT for 4 min, and blocked using block buffer (1 mg/mL bovine serum albumin in PBS) at RT for 30 min. Next, cells were incubated with primary antibodies (1:100–1:200 in block buffer) at RT for 2 h or at 4°C overnight. After washing, cells were incubated with secondary antibodies (1:200 in block buffer) at RT for 1 h. After washing, antifade mount buffer with DAPI (Life Technologies) was added, and cells were covered with cover slides. Images were obtained using an Olympus confocal microscope (FV-1000). For three-dimensional (3D) imaging, 23 to 35 Z-sections were taken at ∼0.1 μm depth. Three-dimensional movies were created using FV-3000 software (Olympus).
For live-cell imaging, cells grown in glass-bottom dishes with opti-MEM medium or FluoroBrite DMEM live-cell imaging medium were imaged at 37°C in an environmental chamber using an Olympus FV-1000 confocal microscope with a 100 × oil objective lens. Recordings were made with free-run for 80 cycles. Images and movies were generated using the FV-3000 software (Olympus).
FRAP assay
Under microscopy, an area of interest of a cell was photobleached using maximum laser power. Images were recorded immediately with free-run for 100–200 cycles. Next, signal intensities in the photobleached area and an adjacent, unbleached control area were measured using FV-3000 software, and the ratios of fluorescent intensities at different times after photobleaching between the photobleached area and the control area were calculated. The time required for maximum recovery (plateau) was determined.
Results
PS-ASOs form distinct subcellular structures when transfected into cells
Previously, we have shown that PS-ASOs, when delivered into cells by transfection at various concentrations, can localize to, or trigger the formation of, different subcellular structures [40]. Some structures are common for both nontoxic and toxic PS-ASOs as tested with multiple sequences of PS-ASOs, such as cytoplasmic P-bodies and ribonucleoprotein (RNP) granules, nuclear PS bodies, paraspeckles, and nuclear filaments [33,34,41–43]. However, some structures are specific to toxic PS-ASOs. For example, almost all toxic PS-ASOs tested tend to localize to the nucleolus and induce the nucleolar mislocalization of paraspeckle proteins, forming distinct nucleolar structures such as perinucleolar caps and diffused nucleolus [43], termed, in this study, as cloudy nucleolus.
Protein markers have been identified for these various structures as summarized in Fig. 1A. PS-ASOs localize to these structures at different times after PS-ASO transfection. Among these structures, paraspeckle proteins are particularly interesting, as they were found in different forms of PS-ASO aggregates, including paraspeckles, nuclear PS-ASO filaments, cloudy nucleolus, perinucleolar caps, and cytoplasmic granules formed with mutant FUS [34,41,43]. In addition, PS-ASOs can also form nuclear PS bodies that contain TCP1β, but not paraspeckle proteins [33,34,48].
To better understand the nature of these PS-ASO-containing structures, HeLa cells were transfected with a nontoxic, Cy3-labeled gapmer PS-ASO (ASO ID 446654), which has 2′-MOE-modified nucleotides flanking the internal deoxy segment. After 5 h, cells were stained for TCP1β and P54nrb, markers of PS bodies and paraspeckles, respectively. Confocal imaging revealed that PS-ASOs were localized in both PS bodies and paraspeckles as expected [34]. In these samples, PS bodies and paraspeckles were spatially distinct structures with no obvious physical connections (Fig. 1B and Supplementary Movie S1), consistent with previous reports [34]. In addition, in ∼5% of cells, PS-ASOs formed nuclear filaments that contained P54nrb and were spatially separated from PS bodies as determined by 3D imaging (Fig. 1C and Supplementary Movie S2). Consistently, a GFP-tagged paraspeckle protein PSF also colocalized with PS-ASOs in paraspeckles and nuclear filaments in live cells (Supplementary Fig. S1A, B). Although the fluorophore used for ASO labeling may potentially affect the chemical properties of PS-ASOs [49,50], the formation of PS bodies and nuclear filaments does not require the fluorophore, as PS bodies and nuclear filaments could be formed in cells transfected with unlabeled PS-ASOs (Supplementary Fig. S1C), consistent with our previous observations [33,34].
ASO-containing paraspeckles are liquid-like structures and the type of 2′-modification influences protein mobility in paraspeckles
To determine the nature of phase separation of PS-ASO-containing paraspeckles, HeLa cells expressing GFP-PSF were treated with different concentrations of 1,6-HD. 1,6-HD disrupts hydrophobic interactions and is commonly used to disrupt structures with liquid-phase separation, but not structures with strong interactions like the case of solid-phase separation [51]. Consistent with previous reports that endogenous paraspeckles are liquid-like structures [52], 1,6-HD treatment with all concentrations tested disrupted the paraspeckle structures within about 10 s in live cells (Supplementary Fig. S2). In subsequent experiments, 10% 1,6-HD was used.
In cells transfected with the Cy3-labeled, 2′-MOE-modified PS-ASO (ASO ID 446654), GFP-PSF colocalized with PS-ASOs in paraspeckle structures, and these structures were quickly disrupted upon 1,6-HD addition (Fig. 2A), suggesting that PS-ASO-containing paraspeckles are also liquid like. To confirm this, FRAP was performed. Proteins within liquid-like structures exchange with the surrounding environment, leading to protein signal recovery after photobleaching [53]. Indeed, GFP-PSF signal in the bleached PS-ASO-containing paraspeckle recovered and plateaued within 80s when normalized to a control paraspeckle without photobleaching (Fig. 2B, C).

PS-ASO paraspeckles are liquid-like structures.
Previously, we have demonstrated that binding of gapmer PS-ASOs to proteins is influenced by the 2′-modification [31], with 2′-F-modified PS-ASOs binding tighter to proteins than 2′-cEt- or 2′-MOE-modified PS-ASOs. For example, gapmer PS-ASOs with 2′-F, cEt, or MOE modifications bind PSF with dissociation constants of ∼0.7, 2.7, and 3.7 nM, respectively [32]. To determine whether the 2′-modification of the PS-ASOs affects PSF mobility in paraspeckles, Cy3-labeled PS-ASOs were synthesized to replace 2′-MOE modification of the PS-ASO (ASO ID 446654) used in previous experiments with either 2′-cEt (ASO ID 598987) or 2′-F (ASO ID 626825) modifications. Results from FRAP assay showed that the signal of GFP-PSF was recovered in paraspeckle structures within 85 and ∼140 s for the cEt-modified and the 2′-F-modified PS-ASOs, respectively (Supplementary Fig. S3A–D). The average PSF recovery time in paraspeckles in cells transfected with the 2′-F PS-ASO (144.0 ± 3.6 s) was significantly longer than recovery time in 2′-cEt PS-ASO- (84.7 + 2.1 s) or 2′-MOE PS-ASO-treated cells (76.0 ± 2.6 s) (Supplementary Fig. S3E). These results indicate that affinity of the PS-ASO for protein is correlated with protein mobility in membrane-less structures.
PS-bodies appear to be solid-like structures
PS-ASOs can also trigger the formation of nuclear PS bodies that contain TCP1β and appear as round-shaped foci. PS body formation requires 10 or more PS backbones, but is independent of ASO sequence, 2′-modifications, or fluorophore labeling [33,34,42,48]. We noted that upon 1,6-HD treatment, PS-ASO-stained PS bodies were not disrupted (Fig. 2A), suggesting that PS bodies may be solid-like aggregates. As GFP tagging of TCP1β at either N- or C-termini blocked localization to PS bodies (data not shown), we further characterized the nature of PS bodies using the Cy3-labeled, PS-MOE ASO (ASO ID 446654). The PS bodies were larger at 24 h than at 4 h after transfection (Supplementary Fig. S4A). In live cells, differential interference contrast (DIC) microscopy revealed that PS bodies are spherical (Supplementary Fig. S4B). PS-ASO-stained PS bodies were not disrupted by 1,6-HD treatment even when treatment lasted 144 s (Supplementary Fig. S4C). In addition, in HeLa cells transfected with the Cy3-labeled PS-ASO (ASO ID 446654), TCP1β-stained PS bodies were resistant to a 5-min 1,6-HD treatment (Supplementary Fig. S4D, E). Similarly, TCP1β-stained foci induced by unlabeled 2′-MOE-modified PS-ASO (ASO ID 116847) were also resistant to 1,6-HD treatment (Supplementary Fig. S4F, G). Together, these results suggest that PS bodies are solid-like structures.
Previously, we found that in cells depleted of RAN, a protein required for nuclear transport, PS body-like structures form in the cytoplasm, which also contain TCP1β and not other TCP1 complex proteins, similar to the nuclear PS bodies [33]. In HeLa cells expressing GFP-PSF, but depleted of RAN using a siRNA, PS bodies formed in the cytoplasm after PS-ASO transfection, as shown by the presence of TCP1β-stained foci (Supplementary Fig. S5A, B). Such cytoplasmic foci were resistant to 5-min 1,6-HD treatment, although paraspeckle structures were disrupted by this treatment (Supplementary Fig. S5C). This suggests that the cytoplasmic PS bodies are also solid-like aggregates.
Nuclear PS-ASO filaments are solid-like structures
Next, we characterized the nature of phase separation in PS-ASO-induced nuclear filaments, which are formed in a small number of cells at high PS-ASO concentrations. These filaments contain PSF and P54nrb [34]. Cy3-labeled PS-ASO (ASO ID 446654) was transfected for 5 h into GFP-PSF-expressing Hela cells, and live-cell imaging was performed. Nuclear filaments containing PS-ASO and GFP-PSF protein were observed in the nuclei, but not nucleolus, of certain cells (Fig. 3A, upper panel). Unlike paraspeckles, the PS-ASO-containing filaments were not disrupted by 1,6-HD treatment (Fig. 3A, lower panel). The PS-ASO-filament structures formed at earlier time (3 h after transfection) were also resistant to 1,6-HD treatment for 2 or 8 min (Supplementary Fig. S6A, B). By DIC imaging, the filaments protrude from the fixed cells (Supplementary Fig. S6C). In a FRAP assay performed in GFP-PSF-expressing cells after transfection with Cy3-labeled PS-ASO (ASO ID 446654), no recovery after photobleaching was observed in the filament structures (Fig. 3B, C). Similarly, no substantial GFP-PSF signal recovery was observed in nuclear filaments induced by the unlabeled PS-ASO counterpart (Supplementary Fig. S6D), indicating no or little protein mobility in these nuclear filaments. This observation is characteristic of the minimal exchange with surrounding environment expected for solid-phase separation. Together, these results indicate that PS-ASO-induced nuclear filaments are solid-like structures.

Nuclear PS-ASO-containing filaments are solid like.
In cytoplasm, PS-ASO-containing P-bodies are liquid like, whereas PS-ASO-containing granules formed with mutant paraspeckle proteins are solid-like structures
PS-ASOs can localize to cytoplasmic P-bodies [42,54], which are involved in mRNA degradation and storage [55]. Consistent with previous reports that P-bodies undergo liquid-phase separation [56], 1,6-HD treatment disrupted the majority of P-bodies (marked with GFP-DDX6) in HeLa cells not transfected with PS-ASOs (Supplementary Fig. S7A). PS-ASO-containing P-bodies were also disrupted by 1,6-HD treatment (Supplementary Fig. S7B), suggesting that PS-ASO-containing P-bodies have similar LLPS properties as naive P-bodies. We note that, although most P-bodies were disrupted by 1,6-HD treatment, a few GFP-DDX6 foci appeared to be more resistant, suggesting that either those structures are not bona fide P-bodies or those structures underwent different phase separation upon 1,6-HD treatment. Understanding the nature of such structures requires further investigation.
Previously, we have shown that PS-ASOs can also localize to cytoplasmic granules formed with mutant FUS (FUS-P525L), a paraspeckle protein, or with a mutant PSF lacking a nuclear localization signal (ΔNLS-PSF) [41]. In HeLa cells that transiently expressed the GFP-FUS-P525L, cytoplasmic granules were not disrupted even after a 10-min treatment with 1,6-HD (Supplementary Fig. S8A), consistent with previous reports that FUS mutant protein can form solid-phase separation [57]. Although some GFP-FUS-P525L proteins localized as dot-like structures in the cytoplasm, such structures are not late endosomes/lysosomes as they did not significantly colocalize with Cy3-labeled PS-ASO (ASO ID 446654) when the PS-ASO was added to cell medium without transfection reagent (Supplementary Fig. S8B). In the absence of transfection reagent, PS-ASOs mainly localize to late endosomes/lysosomes [36,58]. Upon transfection, however, PS-ASOs colocalized with GFP-FUS-P525L granules, and such granules were resistant to 1,6-HD treatment (Supplementary Fig. S8C). These results indicate that the FUS-P525L granules, with or without PS-ASO, are solid-like structures.
Next, we characterized the nature of cytoplasmic GFP-ΔNLS-PSF granules. Such cytoplasmic granules did not contain FUS or P54nrb (Supplementary Fig. S9A, B) and did not co-stain with G3BP1 (Supplementary Fig. S9C), suggesting that the foci containing mutant PSF are not typical stress granules as these contain G3BP1 [59]. In addition, the GFP-ΔNLS-PSF-containing granules do not colocalize with Lamp1 (Supplementary Fig. S9D), a late endosome/lysosome marker.
The GFP-ΔNLS-PSF-containing cytoplasmic granules were resistant to 1,6-HD treatment (Supplementary Fig. S10A), suggesting solid-like aggregates. In support of this, FRAP results showed that the GFP-PSF signal did not recover to more than 20% of control after photobleaching (Supplementary Fig. S10B). Cy3-labeled, PS-MOE ASO (ASO ID 446654) transfected into cells colocalized with the cytoplasmic ΔNLS-PSF granules (Supplementary Fig. S10C), and less than 10% of the control level of GFP-PSF signal was recovered after photobleaching of these PS-ASO-containing granules (Supplementary Fig. S10D). Similarly, the Cy3-labeled PS-ASO with 2′-F modification (ASO ID 626825) colocalized with the GFP-ΔNLS-PSF cytoplasmic granules, and such granules were also resistant to 1,6-HD treatment (Supplementary Fig. S10E). These data indicate that PS-ASO-containing granules formed with mutant paraspeckle proteins are solid-like structures; however, weak but detectable protein exchange may still occur.
Toxic PS-ASO-induced cloudy nucleoli are liquid-like structures
In addition to the above-described PS-ASO-containing structures, we recently found that shortly after transfection, almost all toxic gapmer PS-ASOs, and not nontoxic PS-ASOs, localize to nucleolus together with paraspeckle proteins, and these aggregates contribute to PS-ASO toxicity [43]. We thus characterized the nature of phase separation of these toxic PS-ASO-induced structures. As expected, transfection of HeLa cells with a toxic, Cy3-labeled, PS-cEt ASO (ASO ID 813223) for 4 h caused nucleolar mislocalization of P54nrb (Fig. 4A). Similar nucleolar mislocalization of P54nrb or PSF has been observed with multiple toxic PS-ASO sequences modified with 2′-cEt, MOE, or LNA [43]. Such structures appear to be liquid like, as treatment with 1,6-HD for 1 min cleared P54nrb from the nucleoli, as shown by immunofluorescent staining (Fig. 4B). Similarly, GFP-PSF also mislocalized to the nucleolus in live cells upon transfection with this toxic PS-ASO (ASO ID 813223) (Fig. 4C, upper panel). The nucleolar structures containing the toxic PS-ASO had irregular shapes and were disrupted by 1,6-HD treatment within 8 s (Fig. 4C, lower panel), suggesting that such structures are liquid like. Interestingly, GFP-PSF was observed in distinct, unevenly distributed foci within a cloud of PS-ASO signal (Supplementary Movie S3). We refer to these irregularly shaped structures as cloudy nucleoli.

Toxic PS-ASO-containing cloudy nucleoli are liquid-like structures.
A FRAP assay was performed in GFP-PSF-expressing HeLa cells 4 h after transfection with the Cy3-labeled toxic PS-ASO (ASO ID 813223) (Fig. 4D–F). The GFP-PSF signal had recovered to maximum within ∼80 s, indicating that like paraspeckles, the cloudy nucleoli are liquid like. Similarly, transfection of a Cy3-labeled, LNA-modified version of the toxic PS-ASO (ASO ID 1468683) also led to GFP-PSF localization to cloudy nucleoli, in which the GFP-PSF protein was able to recover after photobleaching (Supplementary Fig. S11). The nucleolar mislocalization of paraspeckle proteins was not due to the fluorophore labeling of the PS-ASOs, since multiple unlabeled toxic PS-ASOs, but not nontoxic PS-ASOs, also caused nucleolar mislocalization of GFP-PSF (Supplementary Fig. S12A), consistent with our previous observations [43]. In addition, GFP-PSF in a cloudy nucleolus formed with an unlabeled toxic PS-ASO (464917) had a recovery time similar to that with the Cy3-labeled, toxic PS-ASO counterpart (ASO ID 813223) (Supplementary Fig. S12B, C and Fig. 4F).
In addition to toxic PS-ASOs, double-stranded siRNAs have been reported to localize to the nucleolus [60]. Indeed, after transfection, fluorophore-labeled siRNAs modified with PS backbone and 2′-OMe or F at certain positions were enriched in nucleoli, but did not recruit paraspeckle protein PSF (Supplementary Fig. S13A). The siRNA-containing nucleolus still displays protein mobility, as demonstrated by FRAP assay in cells expressing GFP-NPM1, a nucleolar protein (Supplementary Fig. S13B–D). However, the recovery of GFP-NPM1 is slower in the presence (∼275 s) than the absence (∼122 s) of siRNAs (Supplementary Fig. S13E, F). These results imply that siRNA-containing nucleoli are most likely liquid-like structures, and the presence of siRNA reduced protein mobility.
Toxic PS-ASOs can cause the formation of perinucleolar cap structures that are liquid like
In addition to the cloudy nucleoli that are formed in majority of cells within 0.5 to 1 h after transfection of toxic PS-ASOs [43], we noted that 4 to 6 h after transfection with the Cy3-labeled toxic PS-ASO (ASO ID 813223), the PS-ASO and GFP-PSF could localize to perinucleolar caps in a small number of cells (Fig. 5A). GFP-PSF in such perinucleolar caps could recover after photobleaching (Fig. 5A, B). Perinucleolar cap localization of GFP-PSF was not due to the labeling dye of the PS-ASO, as similar GFP-PSF localization could be observed with the unlabeled toxic PS-ASO counterpart (ASO ID464917) or with different unlabeled toxic PS-ASOs (Supplementary Fig. S14A). The recovery times of GFP-PSF in the perinucleolar caps are comparable for the toxic PS-ASO with or without Cy3 label (Fig. 5B, Supplementary Fig. S14B, C). These results suggest that the toxic PS-ASOs containing perinucleolar caps are liquid-like structures.
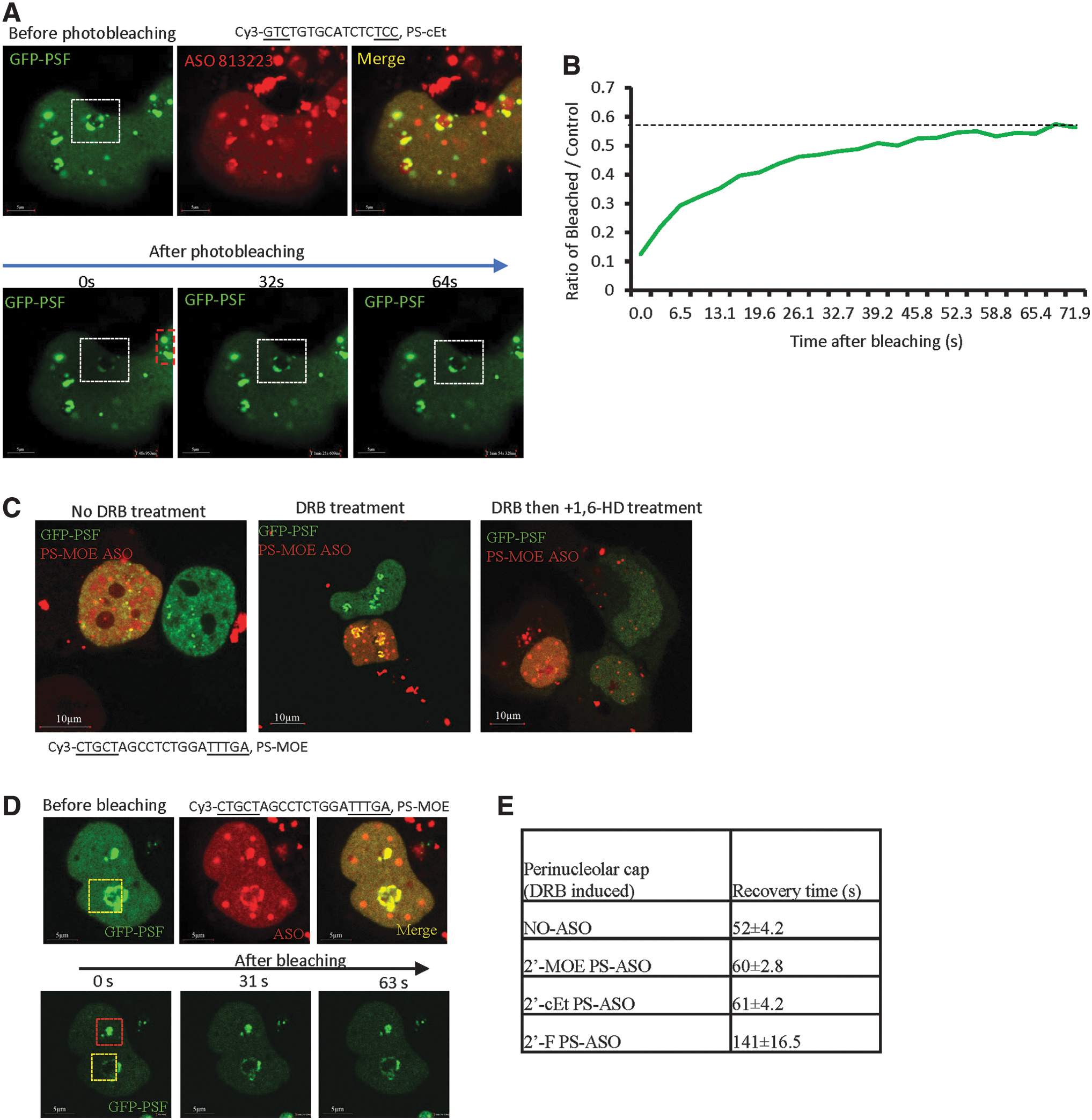
Toxic PS-ASO-induced perinucleolar caps are liquid-like structures.
The localization of paraspeckle proteins to perinucleolar caps has been observed previously upon transcription inhibition [61]. In addition, although nontoxic PS-ASOs do not localize to perinucleolar caps, transcription inhibition can cause relocalization of nontoxic PS-ASOs and paraspeckle proteins to perinucleolar caps [34]. We thus characterized the nature of phase separation in such structures. In the absence of PS-ASOs, inhibition of RNA polymerase II transcription by 5,6-dichloro-1-beta-ribofuranosyl benzimidazole (DRB) treatment caused GFP-PSF localization to perinucleolar caps, which could be disrupted by 1,6-HD treatment (Supplementary Fig. S15A), and the GFP-PSF signal in such structure could be recovered after photobleaching (Supplementary Fig. S15B, C). In the presence of a nontoxic PS-ASO (ASO ID 446654), DRB treatment induced mislocalization of paraspeckle proteins and PS-ASOs to perinucleolar caps, and these caps were also disrupted by 1,6-HD treatment (Fig. 5C). Similar results were observed when transcription was inhibited using ActD (Supplementary Fig. S15D, E). Together, these results suggest that the perinucleolar caps, with or without toxic PS-ASOs, are liquid-like structures. Interestingly, the GFP-PSF recovery time in DRB-induced, 2′-MOE- or 2′-cEt-modified PS-ASO-containing perinucleolar caps after photobleaching was ∼60 s, whereas the recovery time for 2′-F-modified PS-ASO counterparts in such structures was ∼140 s (Fig. 5D, E and data not shown). These observations are consistent with the trend in paraspeckles and further indicate that tighter protein binding reduces protein mobility.
Toxic PS-ASOs can cause the formation of perinucleolar filaments that are solid-like structures
In addition to the cloudy nucleoli and perinucleolar caps identified previously, in this study, we found that toxic PS-ASOs can also induce the formation of other nucleolus-related structures. Five to 8 h after transfection with a Cy3-labeled, toxic PS-ASO (ASO ID 813223), PS-ASO-containing filaments were observed in the nucleolar area in around 5–10% of cells, as shown by co-staining with NPM1 (Supplementary Fig. S16A). Like the nuclear filaments that distribute across the nucleoplasm, the toxic PS-ASO-induced perinucleolar filaments also contained P54nrb, as determined by immunofluorescent staining (Fig. 6A), and also contained PSF, as shown by the colocalization of GFP-PSF with PS-ASOs in live cells (Fig. 6B, upper panels).
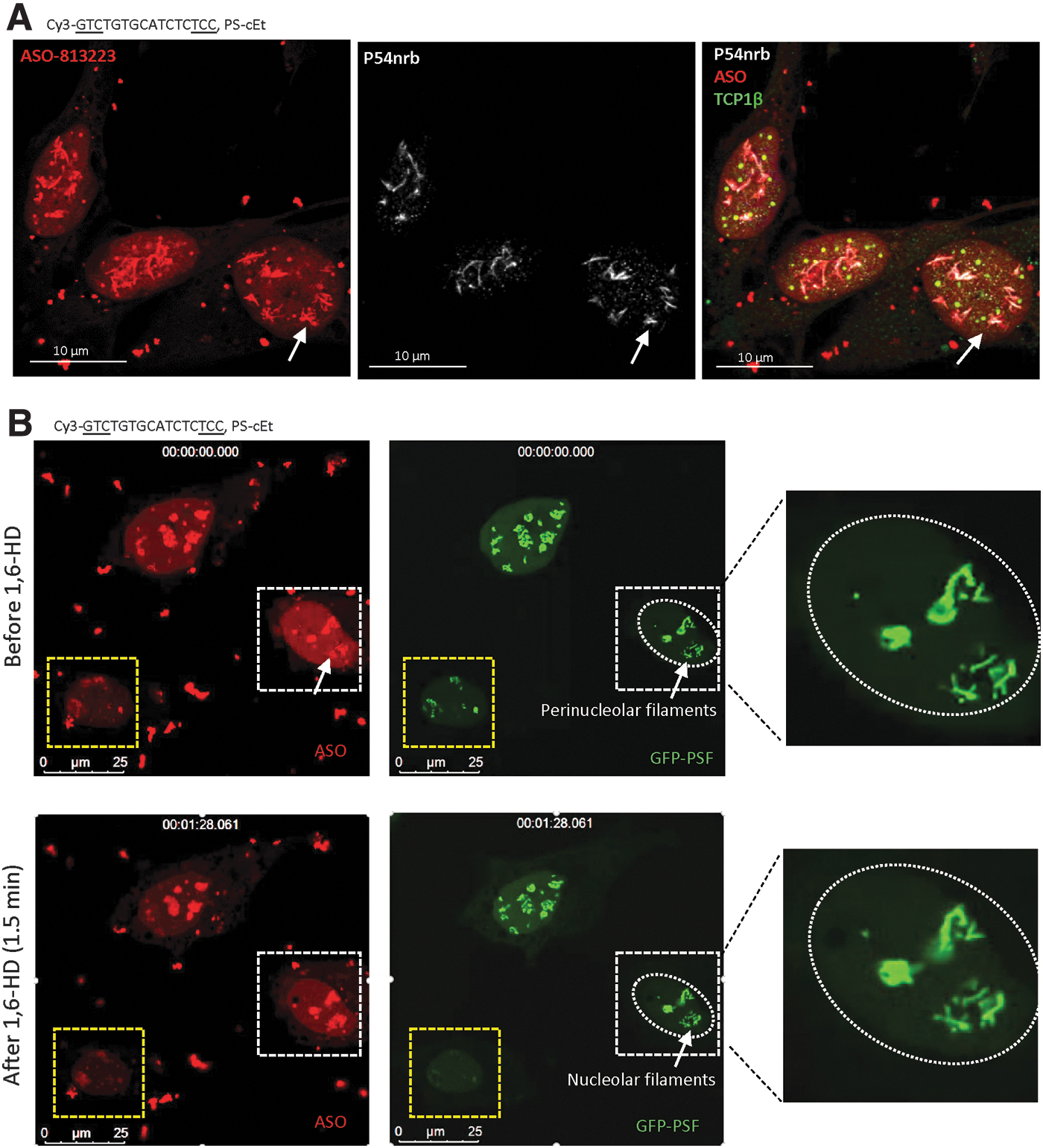
Perinucleolar filaments that contain toxic PS-ASOs are solid-like structures.
Similar to the PS-ASO-containing nuclear filaments, the perinucleolar filaments that include the toxic PS-ASO (ASO ID 813223) were also resistant to 1,6-HD treatment (Fig. 6B, lower panel). In the same experiment, perinucleolar cap structures in one cell were disrupted by 1,6-HD treatment. This result suggests that the toxic PS-ASO-containing perinucleolar filaments are solid like. Consistently, no GFP-PSF signal was recovered in perinucleolar filaments after photobleaching (Supplementary Fig. S16B, C). In addition, formation of perinucleolar filaments with GFP-PSF was also observed with unlabeled toxic PS-ASOs of different sequences or 2′-modifications (Supplementary Fig. S16D), and no obvious GFP-PSF signal recovery was observed after photobleaching in perinucleolar filaments formed by an unlabeled toxic PS-ASO (Supplementary Fig. S16E). Together, these results indicate that the perinucleolar filaments induced by toxic PS-ASOs are solid-like structures.
Toxic PS-ASOs can localize to nucleolus-like structures that are solid-like aggregates
As longer treatment of toxic PS-ASOs led to the change of cloudy nucleolar structure at 2–4 h to perinucleolar cap and perinucleolar filaments at 4–8 h, we examined whether toxic PS-ASOs can form additional structures at a later time. Although transfection with toxic PS-ASOs caused severe death, a small number of cells survived even at 48 h after transfection with the unlabeled toxic PS-ASO (ASO ID 464917) (Supplementary Fig. S17). Next, HeLa cells were transfected with the same toxic PS-ASO labeled with Cy3 (ASO ID 813223), and the PS-ASO localization pattern was monitored over time (Fig. 7A). The toxic PS-ASOs mainly localized in cloudy nucleoli at 3 h after transfection, and in some cells, perinucleolar filaments were observed at 6 h after transfection. At these early time points, PS-ASOs were mostly observed in the nucleoplasm.
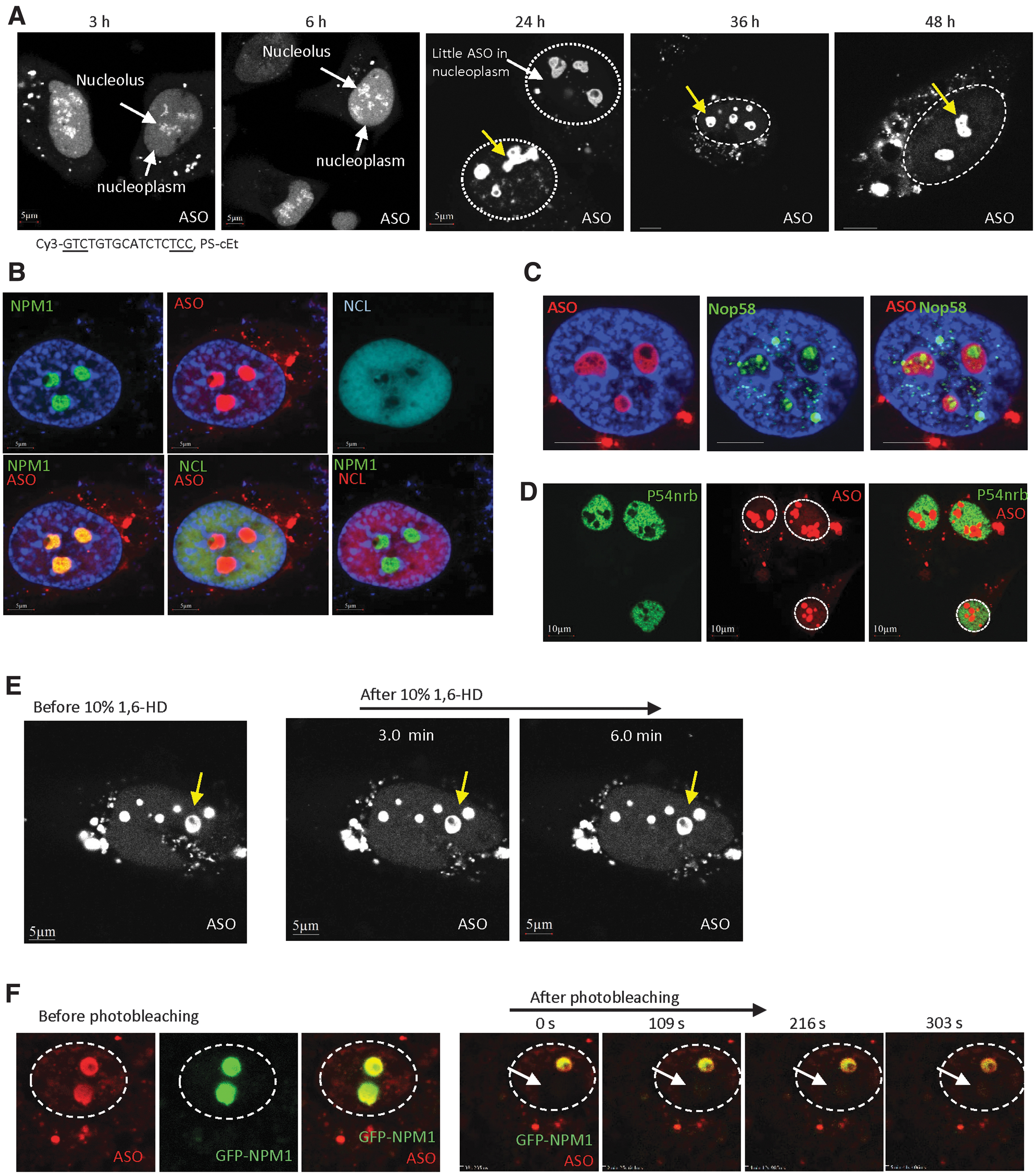
Toxic PS-ASOs can form large nucleolus-like structures at a later time, which undergo LSPS.
Transfection of the Cy3-labeled toxic PS-ASO also caused severe cell death, especially after 24 h of treatment (not shown), yet a small number of cells survived. Interestingly, in cells that were alive at 24 h after transfection, PS-ASOs were observed predominantly in a few large nuclear aggregates (1–3 μm diameter) and were excluded from the nucleoplasm (Fig. 7A). The aggregates contained hollow areas, suggesting that different complexes or structures exist inside the PS-ASO-containing aggregates. Similar localization pattern was observed in cells that survived 36 and 48 h after transfection, implying that such toxic PS-ASO aggregates are relatively stable. Similar structures were also found in HeLa cells transfected with toxic cEt-modified PS-ASOs of different sequences or an LNA-modified toxic PS-ASO (Supplementary Fig. S18A), but were not observed in cells transfected with nontoxic PS-ASOs (Supplementary Fig. S18B). In addition, this structure was able to form in cells transfected with as low as 20 nM toxic PS-ASOs (Supplementary Fig. S18C). The formation of such aggregates of toxic PS-ASOs was also observed in mouse MHT cells, suggesting that it is not unique to a single cell type (Supplementary Fig. S18D).
As the size, number, and morphology of these large PS-ASO aggregates were similar to that of nucleoli, we stained cells transfected with the Cy3-labeled toxic PS-ASO with NPM1 and NCL, protein markers of nucleoli; NPM1 and NCL colocalized in the nucleoli of cells without efficient PS-ASO transfection (Supplementary Fig. S19A), as expected. In cells with large PS-ASO aggregates, NPM1, but not NCL, colocalized with the PS-ASO aggregates (Fig. 7B). This suggests that such toxic PS-ASO aggregates are related to nucleoli, but lack NCL protein, which was relocalized to the nucleoplasm in cells transfected with the toxic PS-ASO. The large PS-ASO aggregates formed at 24 h after transfection were evenly co-stained with NPM1 (Supplementary Movie S4). NPM1, but not NCL, also colocalized with the toxic PS-ASO at perinucleolar cap regions (Supplementary Fig. S19B). Although the toxic PS-ASO-containing cloudy nucleoli observed at 6 h after transfection also contained NPM1, but not NCL (Supplementary Fig. S19C, D), NPM1 did not evenly colocalize with PS-ASOs, but appeared as distinct foci within the cloudy nucleoli (Supplementary Fig. S19D).
To further confirm that the large PS-ASO-aggregates are related to nucleoli, we stained cells that were and were not transfected with the Cy3-labeled toxic PS-ASO (ASO ID 813223) for NOP58, an snoRNP protein that normally accumulates in the nucleoli and Cajal bodies (Supplementary Fig. S19E) [62]. Although NOP58 did not exactly colocalize with toxic PS-ASOs, it localized as distinct foci inside PS-ASO-containing aggregates, especially in the hollow areas that lacked PS-ASO staining (Fig. 7C). These results further indicate that the tight PS-ASO aggregates are related to the nucleoli. We thus refer to these PS-ASO aggregates as nucleolus-like structures. Interestingly, although paraspeckle proteins P54nrb and PSF are present in many PS-ASO aggregates, these proteins were not detected in the nucleolus-like structures (Fig. 7D and data not shown), further suggesting that protein composition can change in different PS-ASO aggregates.
To evaluate the nature of phase separation of the nucleolus-like structures, HeLa cells were transfected for 24 h with the Cy3-labeled toxic PS-ASO (ASO ID 813223). 1,6-HD treatment failed to disrupt the PS-ASO-stained nucleolus-like structures (Fig. 7E). In addition, in GFP-NPM1-expressing HeLa cells transfected with the toxic PS-ASO, recovery of GFP-NPM1 signal was not detected in such structures after photobleaching (Fig. 7F). These data suggest that the toxic PS-ASO-containing nucleolus-like structures are solid-like aggregates.
Cells that survive toxic PS-ASO treatment form solid-like structures and have active RNA transcription
As cells that survived treatment with toxic PS-ASOs had PS-ASO-containing nucleolus-like structures that are solid like, it is possible that the formation of these solid-like aggregates protects cells from toxic PS-ASO-induced death. To evaluate this possibility, we quantified the percentages of cells containing liquid-like structures (eg, cloudy nucleoli) and containing solid-like structures (eg, nuclear filaments, perinucleolar filaments, and nucleolus-like structures) over time after transfection. We reasoned that if solid-like aggregates are protective, cells with solid-like PS-ASO structures should accumulate over time. Indeed, at early time points after transfection with the Cy3-labeled toxic PS-ASO (ASO ID 813223), around 72.5% of cells contained cloudy nucleoli, and the population of such cells declined over time to 12.6% and 2.5% at 24 and 48 h after transfection, respectively (Fig. 8A). However, the percentage of cells with nucleolus-like structures increased from 0% at 6 h to 63% at 24 h, and to 87.5% at 48 h. These results suggest that cells with the cloudy nucleoli either died or survived by converting liquid-like structures to solid-like structures.
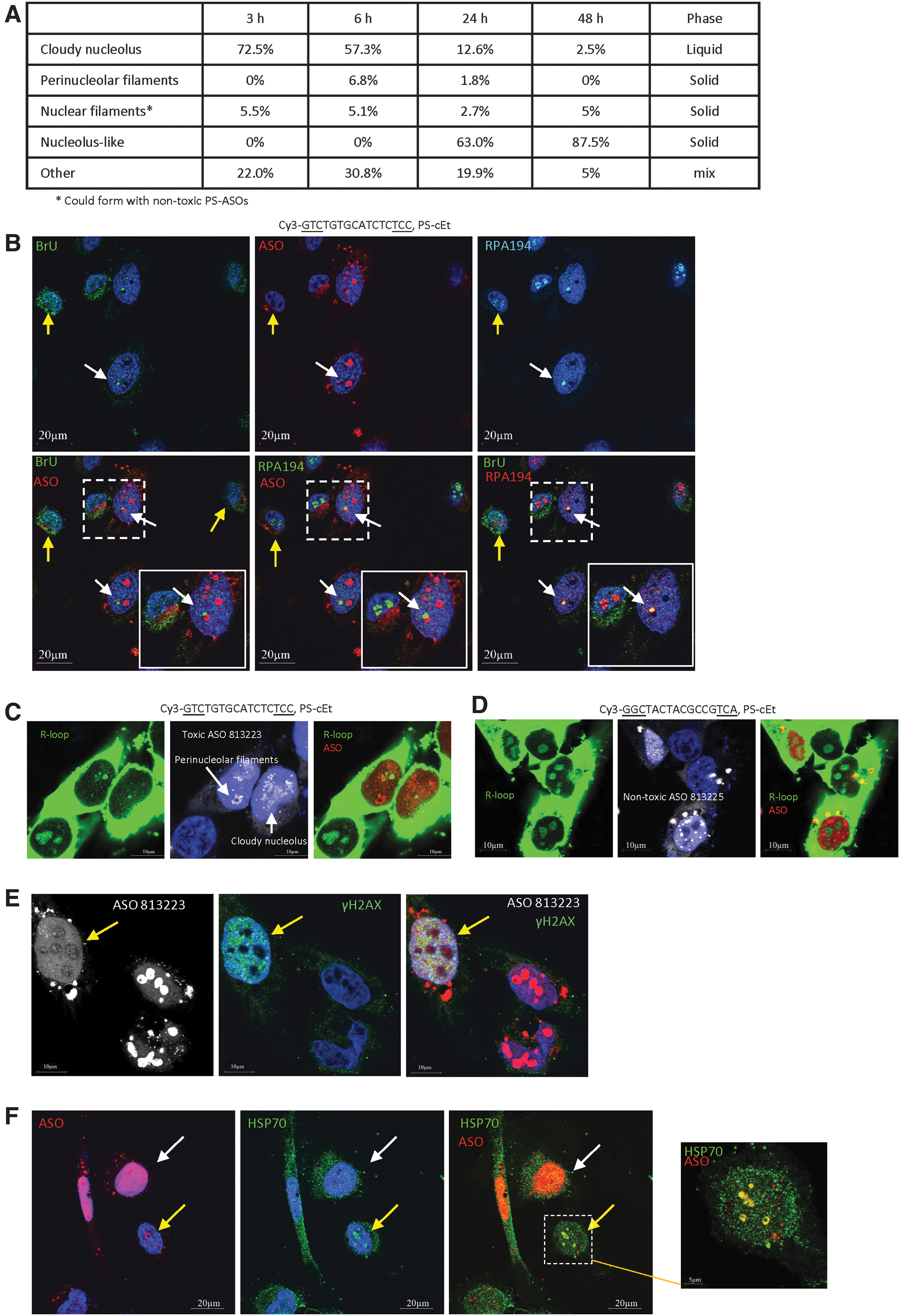
The nucleolus-like PS-ASO structures may provide protective roles.
As the nucleolus is the site of transcription mediated by RNA polymerase I (Pol I), we evaluated the transcriptional activity in cells containing the nucleolus-like PS-ASO structures. At 24 h after transfection with the Cy3-labeled toxic PS-ASO (ASO ID 813223), remaining live cells were incubated with BrU to label nascent RNA, which was subsequently detected by immunofluorescent staining. Compared with cells with no or little nuclear PS-ASO signal due to inefficient transfection, cells with nucleolus-like structures or PS-ASO filaments, both of which are solid like, had weaker nascent RNA signal, but one or two bright BrU-stained foci (Supplementary Fig. S20A, B), suggesting robust transcriptional activity at one or two sites. However, in cells that contain strong nucleoplasmic PS-ASO signal and liquid-like cloudy nucleolar structure, very weak BrU signal was detected, suggesting weaker transcription activity even compared with cells containing nucleolus-like PS-ASO structures (Supplementary Fig. S20C).
The bright BrU foci in cells with nucleolus-like PS-ASO structures colocalized with RPA194 (Fig. 8B), a Pol I transcription factor, suggesting that the BrU staining signal represents Pol I transcription sites that contain nascent pre-rRNA. The sites of transcription were adjacent to, although not exactly colocalized with, the nucleolus-like PS-ASO structures (Fig. 8B, Supplementary Fig. S20D, and Supplementary Movie S5). In addition, RNase H1, an enzyme required for R-loop resolution, was detected in the nucleolus and colocalized with RPA194 in cells not transfected with PS-ASO (Supplementary Fig. S20E). However, RNase H1 was detected adjacent to the toxic PS-ASO-containing nucleolus-like structures and was also adjacent to, but not exactly colocalized with, RPA194 in PS-ASO-transfected cells (Supplementary Fig. S20F). This suggests that the spatial arrangement for transcription and R-loop resolution around the nucleolus-like PS-ASO-containing structures differs from that in control cells.
Next, we evaluated transcription activity in cells containing a few different PS-ASO structures at 3 or 6 h after transfection with the Cy3-labeled toxic PS-ASO (ASO ID 813223) by staining for the R-loop that is formed by co-transcriptional base pairing of RNA with DNA. We note that the R-loop antibody used also nonspecifically stains rRNAs in the cytoplasm [63]. In cells without significant nuclear PS-ASO signal, a strong R-loop signal was detected at nucleoli (Fig. 8C and Supplementary Fig. S20G), consistent with robust Pol I transcription. In cells containing cloudy nucleoli, the R-loop signal in the nucleus was very weak. In contrast, in cells containing solid-like perinucleolar filaments, decent R-loop signal was observed in the nucleus, consistent with BrU labeling of nascent RNA (Fig. 8C and Supplementary Fig. S20B). As a control, transfection with nontoxic PS-ASOs did not substantially affect R-loop signal (Fig. 8D). These results suggest that cells containing solid-like toxic PS-ASO structures have more RNA Pol I transcription activity than cells containing the cloudy nucleolar structures that are liquid like.
Our observations suggest that cells containing solid-like PS-ASO structures such as nucleolus-like structures may be less stressed than cells containing liquid-like PS-ASO structures. To evaluate this possibility, HeLa cells were transfected with the Cy3-labeled toxic PS-ASO (ASO ID 813223) for 24 h, and stained with γH2AX, a marker of double-stranded DNA break that occurs during apoptosis. In cells with nucleolus-like PS-ASO structures, the γH2AX staining signal was much weaker than that in cells containing cloudy nucleoli and strong nucleoplasmic PS-ASO signal (Fig. 8E). At 6 h after transfection of the toxic ASO, cells with cloudy nucleoli also showed strong nuclear staining of γH2AX foci, whereas in a cell without ASO transfection, no decent nuclear γH2AX staining was observed (Supplementary Fig. S21). These results, together with the results of transcription activity, suggest that cells with solid-like PS-ASO structures are less damaged than cells with liquid-like cloudy nucleolar structures.
The nucleolus-like PS-ASO structure contains HSP70 protein
Formation of solid-like structures may be protective, for example, by extracting toxic PS-ASOs from the soluble biological environment. Recent studies have demonstrated that under heat shock, the nucleolus can undergo phase separation and serves as a site for protein refolding by recruiting heat shock protein HSP70, which normally does not localize to the nucleolus [11,12]. These observations promoted us to examine whether HSP70 is present in the nucleolus-like PS-ASO structures, which may contain misfolded proteins due to PS-ASO-protein interactions and phase separation. Indeed, HSP70 was detected to colocalize with Cy3-labeled, cEt-modified toxic PS-ASOs in the nucleolus-like structures in cells (Fig. 8F). HSP70 did not evenly colocalize with the toxic PS-ASOs in the nucleolus-like structures; rather, it was present as distinct foci in these aggregates (Supplementary Movie S6). A similar HSP70 localization pattern was also observed in cells transfected with the same toxic PS-ASO, but lacking the Cy3 dye (Supplementary Fig. S22A). We note that the PS-ASO antibody failed to stain the nucleolus-like structures, likely due to poor accessibility to the solid structure. In addition, the same Cy3-labeled toxic PS-ASO sequence containing LNA modification (Supplementary Fig. S22B) or other Cy3-labeled toxic PS-ASOs also caused nucleolar localization of HSP70 (Supplementary Fig. S22C), suggesting a common feature for toxic PS-ASOs.
However, in cells containing high levels of nucleoplasmic toxic PS-ASOs, HSP70 was mainly detected in the cytoplasm (Fig. 8F). In addition, HSP70 was not present in cloudy nucleoli, perinucleolar caps, PS bodies, or PS-ASO filaments (Supplementary Fig. S22D, E), although the latter two structures are solid like. Even though transfection of a toxic PS-ASO caused HSP70 localization to the nucleolus-like structures, the levels of HSP70 protein were not substantially affected (Supplementary Fig. S22F). As a control, another heat shock protein HSP90 did not relocate to the nucleolus-like structures in cells transfected with the Cy3-labeled toxic PS-ASO (ASO ID 813223) (Supplementary Fig. S22G). These results suggest that HSP70 protein is specifically recruited to the nucleolus-like structures in cells that survive toxic PS-ASO transfection.
Discussion
Protein-phase separation has been shown to play important roles in various biological processes by providing higher order organization of functional compartments [2]. In this study, we characterized, for the first time, the nature of phase separation of various cellular structures that contain exogenous PS-ASOs. Our results showed that upon transfection into cells at high concentrations, PS-ASOs can affect phase separation in various membrane-less structures in a time- and dose-dependent manner. Nuclear paraspeckles, cytoplasmic P-bodies, and toxic PS-ASO induced cloudy nucleoli and perinucleolar cap structures appear to be liquid like. Solid-phase separation, in contrast, is a characteristic of PS bodies, nuclear filaments, and toxic PS-ASO-specific perinucleolar filaments and nucleolus-like structures.
PS-ASOs can interact with different cellular proteins in ways that are affected by PS-ASO sequence, the numbers and positions of PS backbone linkages, and the numbers, positions, and types of 2′-modifications [31,32,40,43]. The interactions between PS-ASOs and proteins can lead to the localization of PS-ASOs to different existing subcellular structures and can result in the formation of new structures [29,40]. Both toxic and nontoxic PS-ASOs can localize to P-bodies and mutant FUS or mutant PSF granules that are cytoplasmic, and PS-bodies, paraspeckles, and nuclear filaments that are present in the nucleus. Since safe PS-ASOs can also form paraspeckle-like structures, PS bodies, and nuclear filaments upon transfection, formation of such nuclear structures appears to be well tolerated and does not contribute to toxicity. However, some structures are specific to toxic PS-ASOs, including cloudy nucleoli, perinucleolar caps, perinucleolar filaments, and nucleolus-like structures. Formation of these nucleolar aggregates can contribute to PS-ASO toxicity by triggering protein mislocalization and degradation, nucleolar stress, and apoptosis, as we recently demonstrated [43]. The nucleolar mislocalization of paraspeckle proteins detected in cell culture strongly correlates with PS-ASO toxicity in vivo, indicating that the same mechanism contributes to toxicity in cells and in animals [43]. Thus, protein mislocalization in cell culture can also be used to predict PS-ASO toxicity in vivo.
Consistent with the protein binding properties, formation of PS-ASO aggregates is affected by the chemistry and sequence of ASOs. PS backbone modification plays a dominant role in enhancing protein binding compared with PO backbone ASOs [45], and more than 12 PS is required for substantial protein binding [31,33]. The increased protein binding by PS modification relative to PO backbone is likely due to extra contact of the sulfur in PS backbone with additional amino acids or even with the same amino acid in the bound protein, as determined by crystallographic studies of PC4 protein complexed with PS-ASOs [64]. The formation of certain structures, especially PS bodies, is PS dependent and reduction of PS numbers to less than 10 abolished PS body formation [33]. In addition, PS-ASO sequence as well as 2′-modification can also significantly affect protein binding and aggregate formation. Toxic PS-ASO sequences tend to bind more proteins more tightly than nontoxic PS-ASOs, leading to the formation of nucleolar aggregates [43]. On the other hand, more hydrophobic 2′-modifications tend to enhance protein binding and may cause protein mislocalization or degradation, like the case of 2′-F-modified PS-ASOs [30]. It is possible that tighter binding of PS-ASOs to proteins may alter protein conformation and protein-protein interactions, reducing protein mobility and ultimately causing phase separation. Moreover, the concentration of PS-ASOs can also affect the formation of certain aggregates. For example, nuclear filaments require high PS-ASO concentration to form, whereas PS-ASO localization to P bodies could be observed at low concentration.
Although these various structures were initially detected with fluorescence-labeled PS-ASOs, similar structures could also form with unlabeled PS-ASOs, as detected previously and also in this study by confocal imaging of marker proteins in different structures, including PS bodies [33] (Supplementary Fig. S1), nuclear filaments (Supplementary Fig. S1), cloudy nucleolus [43] (Supplementary Fig. S12), perinucleolar cap (Supplementary Fig. S14), perinucleolar filaments (Supplementary Fig. S16), and nucleolus-like structures (Supplementary Fig. S22). The results indicate that the formation of these structures was not due to the presence of the fluorescent dye.
Consistent with previous findings that endogenous paraspeckles undergo LLPS [13], we found that PS-ASO-containing paraspeckles are also liquid like as shown by the observations that these structures were disrupted by 1,6-HD treatment and that GFP-PSF protein was recovered after photobleaching. Interestingly, although the times required for maximum GFP-PSF recovery were similar in paraspeckles with or without PS-ASO modified with 2′-MOE (∼76 s), longer times were required for GFP-PSF recovery in paraspeckles containing 2′-F-modified PS-ASOs (∼144 s). Similar results were also observed with the PS-ASO-containing perinucleolar cap structures induced by transcription inhibition. These recovery times correlated with the binding affinity of the PS-ASO with different 2′-modifications to proteins: The 2′-F-modified PS-ASO binds PSF with an affinity ∼5-fold tighter than the 2′-MOE-modified PS-ASO counterpart [30,32]. These data suggest that tight interactions between PS-ASO and protein reduce protein mobility in membrane-less structures.
At high concentrations, PS-ASOs can induce the formation of nuclear filaments that contain paraspeckle proteins such as PSF and P54nrb. The PS-ASO filaments are characterized as solid like, as they are resistant to 1,6-HD treatment and show no GFP-PSF recovery after photobleaching. These results suggest that the paraspeckle proteins in filaments have different binding properties, partners, or conformations than those in paraspeckles. Although the mechanism of initiation of PS-ASO filament formation is unclear, it is possible that the filaments originate from smaller complexes that have already undergone phase separation. In support of this hypothesis, mutant FUS and mutant PSF proteins can form cytoplasmic granules that are solid like. After transfection, PS-ASOs can also localize to these cytoplasmic granules. As the cytoplasmic GFP-ΔNLS-PSF granules do not contain P54nrb and FUS, the formation of such granules may alter the binding properties of PSF, leading to LSPS.
In addition, we found that PS bodies, which are formed within 0.5 to 1 h after transfection of cells with PS-ASO, are solid like. PS bodies are tight structures with round shape that can grow in size over time. PS bodies contain chaperone protein TCP1β, but not RNA binding proteins P54nrb and PSF [34]. As TCP1β binds to PS-ASOs with an affinity more than 100-fold weaker than the affinities of PSF and P54nrb for PS-ASOs [32], it is unclear how PS-ASOs form such a structure with a distinct protein composition. It is possible that other proteins in PS bodies have high binding affinity for PS-ASOs or that the PS body proteins undergo conformational changes upon binding to PS-ASOs to form tight interactions with other components, triggering solid-phase separation.
In the presence of toxic PS-ASOs, PSF and P54nrb mislocalize together with PS-ASOs to the nucleolus within 30 min after transfection and within 5 min after electroporation [43]. These paraspeckle protein-containing cloudy nucleolar structures colocalize with NPM1 and are liquid like. At 4 to 8 h after transfection, toxic PS-ASOs and paraspeckle proteins can localize in certain cells to perinucleolar caps that are also liquid like. However, at 6–8 h after transfection, perinucleolar filaments containing toxic PS-ASO and paraspeckle proteins can form in certain cells that are solid like. These results suggest that the toxic PS-ASOs induce tighter interactions over time among components in these structures.
At 24 h after transfection with toxic PS-ASOs, a small number of cells survived. In most of the surviving cells, we observed nucleolus-like structures that contained toxic PS-ASO and NPM1, but not NCL. The nucleolus-like PS-ASO structures are solid like, and in cells with these structures, the levels of PS-ASOs in the nucleoplasm and in other liquid-like PS-ASO structures were dramatically reduced compared to cells without nucleolus-like PS-ASO-containing structures. These observations suggest that over time, toxic PS-ASOs are removed from the nucleoplasm or other liquid-like structures and sequestered in solid-like aggregates.
The altered phase separation from liquid phase to solid phase of PS-ASO-containing structures over time suggests that toxic PS-ASOs are actively eliminated from the more dynamic biological environment. This type of sequestration was proposed previously to account for the formation of cellular “crystals” that may serve as a storage or detoxication process [65–67]. The formation of solid-like PS-ASO structures may protect cells from toxic PS-ASOs that otherwise would disrupt normal biological processes. The potential protective role of the solid-like PS-ASO structures is supported by the observation that cells with sequestration of toxic PS-ASOs in solid-like structures had more active RNA Pol I-mediated transcription and were less damaged compared to cells with liquid-like PS-ASO structures.
The presence of HSP70 in the nucleolus-like PS-ASO structures suggests that some protein components in those structures are misfolded, and that the recruitment of HSP70 may result in refolding of these proteins to restore normal functions. In support of this hypothesis, recent reports have shown that under heat shock, HSP70 is recruited to the nucleolus [11,12]. This process, which also involves NPM1, has been proposed as a protein quality control mechanism [12]. Upon prolonged heat shock, nucleoli undergo LSPS, and protein refolding does not occur. Currently, it is unclear whether a protein quality control mechanism is involved in cells transfected with toxic PS-ASOs, although the presence of Hsc70 and NPM1 in nucleolus-like PS-ASO structures favors such a possibility.
In summary, our study showed that PS-ASOs can locate to and induce the formation of different subcellular structures that undergo phase separation. These findings explain why the interactions between PS-ASOs and proteins can affect PS-ASO efficacy and toxicity, and suggest that PS-ASOs could be employed to alter phase separation of cellular structures as a therapeutic strategy or as a research tool to better understand biological processes. Moreover, these findings, together with our previous studies on PS-ASO-protein interactions, suggest strategies to improve PS-ASO drug performance by modulating PS-ASO-protein interactions in cells through medicinal chemistry efforts.
Footnotes
Acknowledgments
We wish to thank Tim Vickers, Adam Pollak, Punit Seth, Michael Migawa, and Michael Oestergaard for stimulating discussions; Karla Bernardo for technique assistance; and Tracy Reigle for help in figure preparation.
Author Disclosure Statement
All authors are employees of Ionis Pharmaceuticals, Inc.
Funding Information
This study was supported by an internal funding from Ionis Pharmaceuticals.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.