
Abstract
Newly emerged highly pathogenic A/H7N9 viruses with pandemic potential are effectively transmitted from birds to humans and require the development of novel antiviral drugs. For the first time, we studied the in vitro and in vivo antiviral activity against A/H7N9 of oligodeoxyribonucleotides (ODNs), which were delivered into the cells in the proposed TiO2-based nanocomposites (TiO2∼ODN). The highest inhibition of A/H7N9 in vitro (∼400-fold) and efficient, sequence-specific, and dose-dependent protection (up to 100%) of A/H7N9-infected mice was revealed when ODN was targeted to the conserved terminal 3′-noncoding region of viral (−)RNA. After the treatment with ODN, the virus titer values in the lungs of mice decreased by several orders of magnitude. The TiO2∼ODN nanocomposite did not show toxicity in mice under the treatment conditions. The proposed approach for effective inhibition of the A/H7N9 can be tested against other viruses, for example, new emerging influenza viruses and coronaviruses with pandemic potential.
Introduction
Humanity is attacked annually by RNA-containing viruses. As a rule, these are Influenza A and B viruses and new coronaviruses that have recently covered almost the entire world. The fight against the influenza A virus (IAV) remains a significant problem because of mutations that lead to the appearance of new virulent subtypes. The new subtype of IAV, A/H7N9, which usually circulated among birds [1], was first identified by WHO in humans in China in 2013.
Whereas only low pathogenic avian A/H7N9 subtypes were detected during the first four waves, the fifth wave in 2017 has been the most severe with high mortality rates in humans (up to 50%) [2], thus developing into highly pathogenic avian influenza viruses [3]. Although A/H7N9 is effectively transmitted from birds to humans [4], the possibility of human-to-human transmission cannot be completely excluded [5]. The H7N9 virus continues to accumulate mutations, and its affinity for human respiratory epithelial sialic acid 2–6 receptor is increasing. Therefore, a pandemic is still possible. The latest data on the A/H7N9 virus are summarized in Wang et al. [6].
The A/H7N9 virus becomes highly pathogenic with pandemic potential and less sensitive to common old small-molecular drugs, for example, adamantane and oseltamivir [3,6–8]. New drugs, such as the inhibitor of PA endonuclease, baloxavir marboxil [9], were not very successful because A/H7N9 quickly becomes resistant to those drugs. Therefore, novel antiviral drugs that overcome existing limitations to treat A/H7N9 infection are urgently needed. A rational approach to the creation of effective antiviral drugs is the use of nucleic acid (NA) fragments, which can inhibit the expression of certain genes due to complementary interactions with the viral genome. The advantage of NA fragments is in their selective recognition and binding to complementary regions of target genes, thus preventing their functioning, and in acceptable toxicity profiles [10–12]. Some NA-based drugs are already approved or under clinical study [13].
The main limitation of using therapeutic NA is their poor penetration into cells. The search for effective delivery systems for NA-based drugs is an urgent task. Viral vectors, liposomes, cationic polymers of various nature, transport peptides, and other transport systems are used for cell transfection [14]. Despite the efforts made, the task cannot be considered complete.
In our previous work, we proposed and developed a new strategy for delivering DNA fragments to cells based on the use of TiO2•PL-ODN (further, TiO2∼ODN) nanocomposites consisting of delivery-promoting titanium dioxide nanoparticles and immobilized polylysine-containing oligodeoxyribonucleotides (PL-ODNs) [15–20].
It was shown that the same oligodeoxyribonucleotides (ODNs) delivered in the proposed nanocomposites to the virus-infected MDCK cells and targeted at conserved sites of IAV segment 5 of different subtypes (H3N2, H5N1, and H1N1) inhibited virus reproduction with high efficiency of 3–4 orders of magnitude [19]. These results were a serious motivation for further studies of the effect of oligonucleotides on highly pathogenic, including newly emerged, IAV subtypes.
In this work, we studied for the first time the in vitro and in vivo inhibition of the reproduction of the recently emerged pathogenic A/H7N9 virus with pandemic potential using ODNs targeted to conserved RNA regions of this virus.
Materials and Methods
Reactants
Chemicals were obtained from commercial suppliers. We used: RPMI-1640 medium; antibiotics (BioloT);
Oligonucleotides and nanocomposites
We used phosphodiester oligodeoxyribonucleotides (ODNs, Table 1), which were synthesized by the phosphoramidite method on an ASM-800 DNA synthesizer (Biosset) using phosphoramidite monomers (Glen Research).
Oligodeoxyribonucleotides Targeted at Different Regions of (−)RNA and (+)RNA of Influenza A Virus Segment 5
ODN9 and ODN10 were used as negative controls. ODN9 is complementary to the AUG-containing region of Bovine viral diarrhea virus (strain Oregon C24V, GenBank). Random ODN10 was taken from the work of Ge et al., who studied the effect of morpholino oligonucleotides against IAV [22].
Numbering of nucleotides are given for (+)RNA.
Noncoding region.
Coding region.
IAV, influenza A virus; ODN, oligodeoxyribonucleotide.
The synthesis of polylysine-containing oligodeoxyribonucleotides (PL-ODNs) and nanocomposites TiO2∼ODN consisting of anatase nanoparticles and PL-ODN conjugates were fulfilled as described in Levina et al. [15]. The capacity of the nanocomposites for oligonucleotides was 20 and 60 nmol/mg for the in vitro and in vivo experiments, respectively.
Antiviral activity of nanocomposites in vitro
The experiments were fulfilled as described in Levina et al. [23]. In short, the MDCK cells were initially infected with A/H7N9 at a multiplicity of infection (MOI) of 0.1 TCID50/cell. After the virus adsorption (room temperature, 1 h) and the removal of the excess of the virus, the tested samples in RPMI-1640 medium without trypsin (100 μL/well; the final concentration, 5 μg/mL for the nanocomposite or 0.1 μM for ODN) were applied to the infected MDCK cells. The cells were incubated for 4 h (37°C, 5% CO2, and 100% humidity), followed by washing with the same medium. The RPMI-1640 medium containing trypsin was added to each well (100 μL), and the cells were incubated for 48 h. Serial 10-fold dilutions (10−1–10−8) of the virus-containing liquid from each well were applied to MDCK cells for 48 h to evaluate the virus titer. The presence of the virus was visually determined by the hemagglutination reaction with a 1% suspension of chicken erythrocytes.
Antiviral activity of nanocomposites in vivo
The outbred ICR mice (13–15 g; FBRI Vector) were kept under standard conditions; water and balanced food were provided without limitation. All experiments were conducted according to international rules for research using experimental animals (UFAW Handbook), followed guidelines for the maintenance and use of laboratory animals [24], and were approved by the Bioethics Committee (IRB) of the State Research Center of Virology and Biotechnology “Vector” (Department of the Federal service for supervision of consumer protection and human welfare, Russia).
The mice were divided into control and experimental groups of 10 mice each. The nanocomposite (TiO2∼ODN8) was administrated intraperitoneally at a single dose/mouse of 0.04, 0.08, and 0.12 mg of TiO2 bearing ODNs (60 nmol/mg) in 100 μL of physiological solution (∼1, 2, and 3 mg of ODN per 1 kg of body weight). In an hour, the anesthetized mice were infected intranasally with 10 LD50 of A/H7N9. Six hours later, the mice were again treated with the nanocomposite at the same dose. The procedure was repeated twice a day for the next 4 days. The mice were monitored for 16 days. The TiO2∼ODN10 nanocomposite and TiO2 nanoparticles at a single dose/mouse of 0.04 and 0.08 mg were used as controls under the above regime. Reference preparation oseltamivir (Hoffmann-La Roche) was administered orally at a dose of 100 μL (1.5 mg/mL, 10 mg/kg).
To evaluate the virus titer in the lungs of mice, they were divided into one control and four experimental groups of six mice each. The control mice were A/H7N9 infected but not treated with the samples. The experimental mice were infected and treated with TiO2∼ODN1 and TiO2∼ODN10 at a single dose of 0.04 and 0.08 mg/mouse as described above. The virus titer in the lungs was evaluated on the third and sixth day postinfection taken by two mice from each group. The lung tissues from sacrificed mice were homogenized in the RPMI-1640 medium at 1:10 (w/v) that contained penicillin (100 mU/mL) and streptomycin (100 μg/mL). Homogenates were centrifuged (10,000 rpm, 15 min) to pellet debris. Serial 10-fold dilutions of the lung homogenate supernatant were added to 96-well plates (0.1 mL for each well, 4 wells for each dilution). Each well contained MDCK cell monolayer in the RPMI-1640 medium with antibiotics without serum. MDCK cells were grown in 96-well plates in the RPMI-1640 medium containing 5% inactivated calf serum (HyClone), penicillin (100 mU/mL), and streptomycin (100 μg/mL) until the formation of a monolayer (100,000 cells/mL, 100 μL/well). After incubation of the cells with lung homogenates (2 days, 37°C, 5% CO2), the virus-containing solution (50 μL) was transferred to the wells of a round-bottom plate, followed by the addition of a 1% suspension of chicken erythrocytes (50 μL) in each well, and incubation for 40 min at room temperature. The presence of the virus was visually determined by the hemagglutination reaction.
Statistical analysis
Statistical analysis was carried out using the Statistica 7.0 program. The significance of the percent survival was determined by a Chi-square (χ2) analysis. The Student's t-test and the Mann–Whitney U-test were used to analyze the virus titer in the lungs [25]. The indicators of life expectancy are presented as M ± I95, where M is the arithmetic mean and I95 is the confidence interval. Comparison of these indicators for mice in different groups was performed using the Mann–Whitney U-test. The differences were considered statistically significant at P ≤ 0.05.
Results
Inhibition of A/H7N9 in vitro
First, we studied the antiviral activity of the proposed TiO2∼ODN nanocomposites in the A/H7N9-infected MDCK cells. We used 19–21-meric ODNs targeted at four selected conserved regions of both (−)RNA and (+)RNA of IAV segment 5 encoding the nucleoprotein (NP) (Table 1). As before [23], we chose the four most conserved target regions in segment 5 of the IAV genome, that is, the 5′-noncoding region (2–22), the region containing the initiation AUG codon (34–54), the coding region (1498–1516), and the noncoding region at the 3′ end of (+)RNA (1544–1564). Some examples of the sequences of the chosen regions in segment 5 of different A/H7N9 strains taken from the GenBank database are presented in Table 2 in comparison with the same regions of the H5N1, H3N2, and H1N1 subtypes, which were studied in our previous works [19,23]. The data strongly suggest that the studied regions in all IAV subtypes and strains are truly conserved. Only the AUG-containing region contains insignificant mutations. Therefore, the use of preselected oligonucleotides to affect segment 5 of the A/H7N9 virus was justified.
Sequences of Complementary DNA [Corresponding to (+)RNA] of the Chosen Regions of Segment 5 of Different Influenza A Virus Subtypes and Strains from GenBank
Complete 5′ and 3′ sequences of different strains (1,565 nt), except strains i (1,544 nt) and m and n (1,497 nt) are presented; the chosen conserved regions are in bold and underlined; the initiation codon is not underlined; point mutations are in gray.
Strains: H5N1(a), A/chicken/West Java(Sbg)/29/2007; H5N1(b), A/duck/Shandong/093/2004; H3N2(c), A/swine/Guangdong/113/2002; H3N2(d), A/Bilthoven/16190/1968; H1N1(e), PR/8/34; H1N1(f), A/swine/Beijing/21/2008; H7N9(g), A/blue-wingedteal/Guatemala/CIP049-02/2008; H7N9(h), A/chicken/Hangzhou/174/2013; H7N9(i), A/Zhejiang/DTID-ZJU10/2013; H7N9(j), A/turkey/Minnesota/1/1988; H7N9(k), A/Kunming/KMCDC-YHY/2017; H7N9(l), A/chicken/Guangdong/J2/2017; H7N9(m), A/duck/Anhui/SC702/2013; H7N9(n), A/chicken/Anhui/AH395/2017.
The A/H7N9 inhibition with oligonucleotides targeted to (−)RNA of IAV appeared to be more efficient as compared with oligonucleotides targeted to (+)RNA (comp. gray and black columns in Fig. 1). The highest inhibition of the IAV reproduction was in the case of oligonucleotides ODN1 and ODN2 (columns 1 and 2; inhibition by a factor of ∼400). ODN1 is targeted to the terminal 3′-noncoding region of (−)RNA, and ODN2 is targeted to the neighboring region, which is complementary to the AUG-containing region in (+)RNA. ODN6 targeted to the (+)RNA region that contained the initiation AUG codon (column 6), exhibited slightly less effect (∼200-fold). The TiO2∼ODN3 and TiO2∼ODN5 nanocomposites showed 60-fold and 25-fold inhibition, respectively (columns 3 and 5). The other tested oligonucleotides (columns 4, 7, and 8) and the control samples, that is, TiO2∼ODN9, TiO2∼ODN10 (columns 9 and 10), and the mixture of TiO2 nanoparticles with the most efficient ODN1 (column 11) showed the antiviral activity similar to the control (column 12).
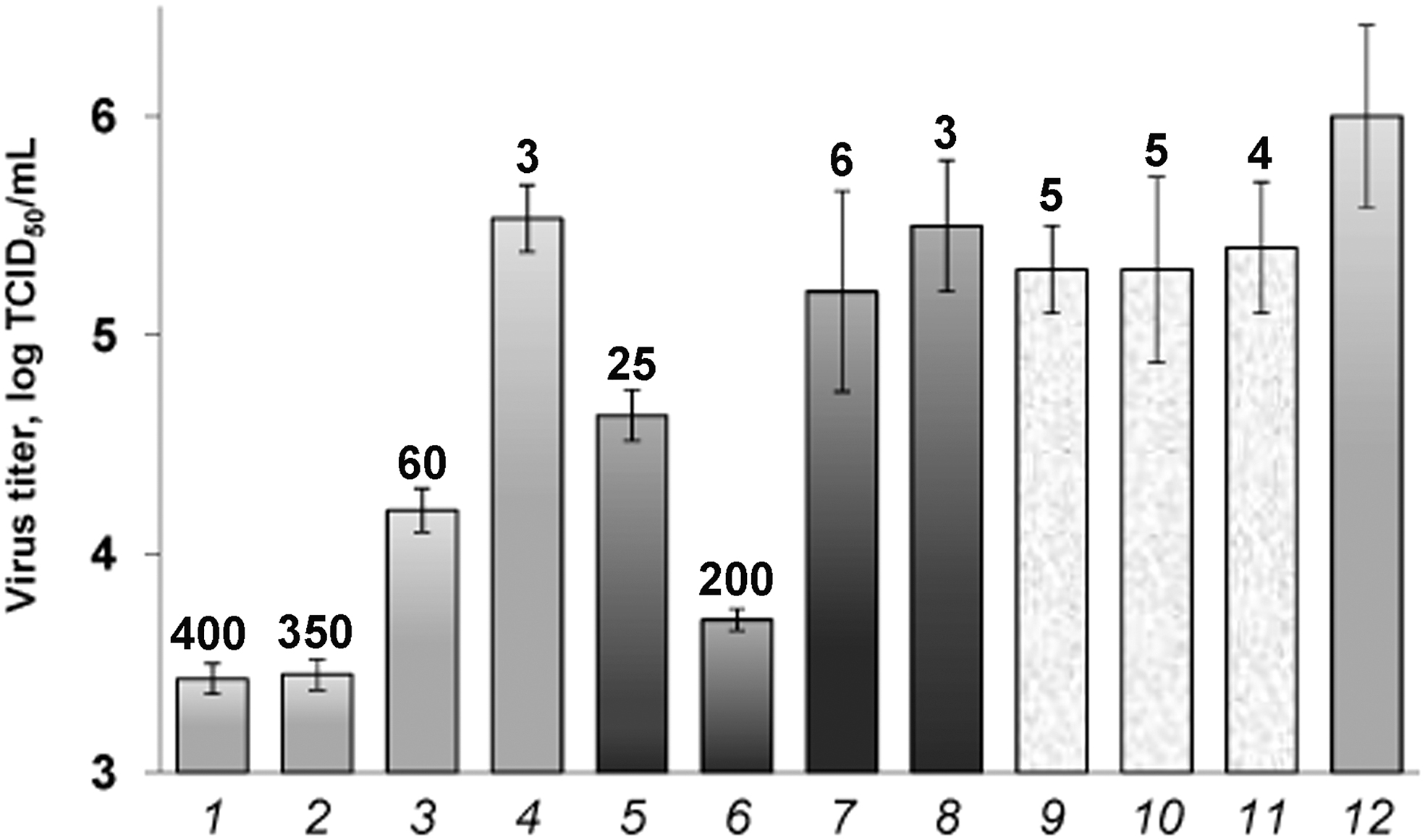
Inhibition of A/H7N9 subtype with the TiO2∼ODN nanocomposites in infected MDCK cells. 1, TiO2∼ODN1; 2, TiO2∼ODN2; 3, TiO2∼ODN3; 4, TiO2∼ODN4; 5, TiO2∼ODN5; 6, TiO2∼ODN6; 7, TiO2∼ODN7; 8, TiO2∼ODN8; 9, TiO2+ODN1; 10, TiO2∼ODN9; 11, TiO2∼ODN10; 12, virus control without sample. The numbers above the columns are the values of n-fold inhibition of IAV replication in comparison with the virus control. IAV, influenza A virus; ODN, oligodeoxyribonucleotide.
Earlier, we evaluated the concentration of samples resulting in 50% cell death (TC50) for TiO2∼ODN as ∼1,800 μg/mL [16]. The used concentration of TiO2∼ODN1 (5 μg/mL) leads to 400-fold inhibition of the H7N9 virus replication in the cell system, which means that IC50 would be much lower. Consequently, the selectivity index SI = TC50/IC50 can be estimated as 1,800 /<< 5 = >> 360.
Toxicity of nanocomposites in vivo
The intraperitoneal administration of the nanocomposite in uninfected mice does not cause the death of animals. The comparison of the weight of the body and organs of intact and treated mice after intraperitoneal injection of the TiO2∼ODN1 nanocomposite showed no difference during the observation period (Supplementary Fig. S1 and Supplementary Table S1).
Visual examination of mice for 14 days also did not show any changes in the state of the coat, mucous membranes of the eyes, nose, mouth, and animal behavior. These results indicate that the studied nanocomposite is nontoxic to mice under the used conditions.
Inhibition of A/H7N9 in vivo
We examined inhibition of the A/H7N9 virus in infected ICR mice using the intraperitoneal administration of the most efficient TiO2∼ODN1 nanocomposite at different doses.
The antiviral effect of TiO2∼ODN1 was high enough even at the lowest dose we used (the survival rate of mice at a dose/mouse of 0.04 mg was 70%; Fig. 2, line f). As expected, this effect increased at the higher doses and reached 90% and 100% at a dose of 0.08 and 0.12 mg/mouse (lines j and i, respectively), which exceeded the effect of a common reference preparation oseltamivir (line g). The difference of these results from that for the infected untreated mice (line a) was statistically significant (P ≤ 0.05). The treatment of the infected mice with TiO2 nanoparticles and TiO2∼ODN10 did not lead to significant changes in the survival of mice as compared with the control group (lines b–e vs. line a). However, the difference of these results from the effect of TiO2∼ODN1 at the corresponding concentrations was pronounced (lines b and c vs. line f and lines d and e vs. line j). The high antiviral activity of TiO2∼ODN1 was confirmed by the data on its protective factors (60-90 depending on the dose) and life expectancy values, which were comparable or even higher than these parameters for oseltamivir (Supplementary Table S2).
Protective effect of the studied samples under intraperitoneal injection in ICR mice infected with the A/H7N9. Control without a sample (a); TiO2 and TiO2∼ODN10 at a dose/mouse of 0.04 mg (b, c); TiO2 and TiO2∼ODN10 at a dose/mouse of 0.08 mg (d, e); TiO2∼ODN1 at a dose/mouse of 0.04 mg (f), 0.08 mg (j) and 0.12 mg (i); oseltamivir (g). Asterisks indicate a significant difference from the control (without samples) at P ≤ 0.05 according to a Chi-square (χ2) analysis.
The virus titer was evaluated in the lungs of A/H7N9-infected mice treated with the TiO2∼ODN1 and TiO2∼ODN10 nanocomposites 3 and 6 days after the first injection (Fig. 3). On the third day, the virus titer significantly decreased in the mouse lungs in the presence of TiO2∼ODN1 (by 1.7 and 2.3 orders of magnitude at 0.04 and 0.08 mg/mouse, respectively, compared with the control group). Even a much greater effect was revealed on the sixth day after the treatment, that is, the virus titer decreased by 5.6 and 5.8 orders of magnitude at the above doses, compared with the control group. Almost no differences in the virus titers were observed between the TiO2∼ODN10-treated and untreated mice at both doses on either the third or sixth days. The results on the virus titer in the lungs (Fig. 3) were statistically significant when compared with the control group of infected mice (P ≤ 0.05) and correlate well with the data on the survival rate of mice (Fig. 2).

Virus titer in the lungs of A/H7N9-infected mice. Mice were treated with the studied samples at a dose/mouse of 0.04 mg (a) and 0.08 mg (b) on the third day (light-gray columns) and the sixth day (dark- gray columns) after infection. Lung titers were compared by Student's t-test. Asterisks indicate a statistically significant difference from the control at P ≤ 0.05 according to the Student's t-test.
Discussion
It is known that the genome of the IAV mutates constantly (especially genes encoding surface proteins, hemagglutinin and neuraminidase, which determine the subtype of the virus). Nevertheless, the viral genome contains conserved regions, the selection of which determines a successful action of NA-based drugs [23,26–31] against various, including newly emerging, IAV subtypes.
IAV contains eight single-stranded RNA segments of negative polarity as the genome. Segment 5 encodes highly conserved NP, which performs multiple essential functions throughout the virus life cycle (the organization of RNA packing, nuclear trafficking, viral RNA [vRNA] transcription, and replication, etc.). NP has no host cellular counterpart; thus, a high SI could be achieved for NP inhibitors. The multifunctional role of NP in the virus life cycle, coupled with the high conserved sequence, makes it an ideal drug target [31 and references therein]. As in our previous studies, we used, in this study, this segment as a target to affect viral replication.
Some publications described the use of NA fragments against different IAV subtypes [26,32,33]. The small interfering RNA (siRNA)/chitosan nanoparticle and siRNA/cell-penetrating peptide complexes protected mice from A/H1N1infection by 50%–60% [30,34]. Oligonucleotides complementary to different regions of vRNA and messenger RNA (mRNA) of the H5N1 virus reduced the viral reproduction in MDCK cells by factors of up to 8 [28,29]. There is currently no data on the use of NA-based agents for the A/H7N9 subtype. We used for the first time NA-based drugs against the new highly pathogenic A/H7N9 subtype.
After infection of cells with the influenza virus, vRNA segments (negative strands) are transcribed into mRNAs and replicated into complementary RNAs (positive strands) [35]. We studied the antiviral effect of the proposed nanocomposites bearing oligonucleotides complementary to chosen regions of both negative and positive strands of segment 5. The sequences of these regions of RNA segment 5 of different IAV subtypes, including A/H7N9 are conserved (Table 2).
The efficiency of the inhibition of A/H7N9 replication with oligonucleotides addressed to viral (−)RNA is higher than that of targeted to (+)RNA (Fig. 1). The most vulnerable sites in segment 5 of A/H7N9 were the terminal 3′-noncoding region of (−)RNA and the neighboring region, which is complementary to the AUG-containing region in (+)RNA (the targets for ODN1 and ODN2, which inhibited A/H7N9 by factors of 400 and 350, respectively, Fig. 1). The 3′-noncoding region contains the conserved sequence of 12 nucleotides common for all 8 RNA segments of various IAV strains [27]. Most likely, the observed strong effect of ODN1 is partially explained by its simultaneous action to the other segments of the viral genome. The region that contained the initiating AUG codon in (+)RNA was also the “Achilles' heel” of the virus genome (∼200-fold inhibition). Thus, it can be assumed that the in vitro pattern, which has been identified for the previously existing H3N2, H5N1, and H1N1 subtypes [23], is also inherent in newly emerging pathogenic IAV subtypes with pandemic potential. Our results indicate a possibility of using the TiO2∼ODN nanocomposites to affect not only existing but also newly emerged IAV subtypes, provided that ODN is targeted to the conserved regions of vRNA.
We studied the inhibition of pathogenic A/H7N9 in infected mice with the proposed TiO2∼ODN1 nanocomposite that contained the most active ODN1 targeted to the 3′-terminal region of (−)RNA of IAV segment 5 (Fig. 1). First, it was shown that the proposed TiO2∼ODN nanocomposites are not toxic to mice under the conditions used (Supplementary Fig. S1 and Supplementary Table S1). The TiO2∼ODN1 nanocomposite showed a high protective effect against the A/H7N9 virus in the infected mice (100% survival), which exceeded the action of oseltamivir (Fig. 2 and Supplementary Table S2). The treatment of the infected mice with the control samples (TiO2 nanoparticles and the TiO2∼ODN10 nanocomposite) did not lead to significant changes in the viability of mice. It should be noted that the proposed approach using the TiO2∼ODN nanocomposites provides the more effective inhibition of the IAV subtypes compared with the other used NA-based agents in vitro (eg, Kumar et al. [36], Zhang et al. [37], Zhiqiang et al. [38], and Khantasup et al. [39]) and in vivo (eg, Kumar et al. [36], Gabriel et al. [40], Rajput et al. [41]). TiO2 nanoparticles like other particles can protect oligonucleotides against nucleases. Oligonucleotides in our nanocomposites are noncovalently attached to TiO2 nanoparticles, thus allowing them to be released from the nanocomposites after delivering into cells, enter the nuclei, and affect the target vRNA without interference from nanoparticles [17]. The high penetrating ability of nanocomposites and the long-term stability of oligonucleotides provide their highly efficient action on vRNA in vitro and in vivo.
The virus titers in the lungs of A/H7N9-infected mice treated with the TiO2∼ODN1 was reduced by about two and six orders of magnitude on the third and sixth days, respectively, after the treatment of mice with the TiO2∼ODN1 nanocomposite, whereas the use of the TiO2∼ODN10 nanocomposite resulted in almost no effect as compared with the control group (Fig. 3). These results indicate the possibility of the ODN delivery into the lungs of mice. The survival rate of mice is due to a significant decrease in the virus titer in the lungs (comp. Figs. 2 and 3).
Our results demonstrated for the first time the efficient, sequence-specific, and dose-dependent effect of phosphodiester oligonucleotides on the new pathogenic A/H7N9 subtype with pandemic potential in vitro and in vivo. We anticipate that the proposed approach to virus inhibition can be tested for other viruses, for example, new emerging influenza viruses and coronaviruses with pandemic potential. The developed method may find a wide application not only for inhibition of viral genes but also for other genes responsible for various (hereditary, oncological, etc.) diseases associated with NA.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.