
Abstract
Known limitations of unfractionated heparin (UFH) have encouraged the evaluation of anticoagulant aptamers as alternatives to UFH in highly procoagulant settings such as cardiopulmonary bypass (CPB). Despite progress, these efforts have not been totally successful. We take a different approach and explore whether properties of an anticoagulant aptamer can complement UFH, rather than replace it, to address shortcomings with UFH use. Combining RNA aptamer 11F7t, which targets factor X/Xa, with UFH (or low molecular weight heparin) yields a significantly enhanced anticoagulant cocktail effective in normal and COVID-19 patient blood. This aptamer-UFH combination (1) supports continuous circulation of human blood through an ex vivo membrane oxygenation circuit, as is required for patients undergoing CPB and COVID-19 patients requiring extracorporeal membrane oxygenation, (2) allows for a reduced level of UFH to be employed, (3) more effectively limits thrombin generation compared to UFH alone, and (4) is rapidly reversed by the administration of protamine sulfate, the standard treatment for reversing UFH clinically following CPB. Thus, the combination of factor X/Xa aptamer and UFH has significantly improved anticoagulant properties compared to UFH alone and underscores the potential of RNA aptamers to improve medical management of acute care patients requiring potent yet rapidly reversible anticoagulation.
Introduction
Each year, cardiac surgeons perform cardiopulmonary bypass (CPB) on over a million patients worldwide to facilitate cardiac surgical operations [1,2]. Since 1953, when CPB was performed to facilitate the first successful open cardiac operation, unfractionated heparin (UFH) has been the standard CPB anticoagulant [2]. Intravenous (IV) UFH produces a potent, rapid-onset, and titratable anticoagulant effect that is usually readily reversible with IV protamine [3]. UFH binds and augments the inhibitory activity of the constitutive circulating serine protease inhibitor antithrombin (AT). UFH-AT complexes inhibit multiple procoagulant proteases, particularly thrombin, factor (F)Xa, and FIXa, through the formation of stable covalent bonds between AT and catalytic sites of those proteases [2,4].
Unfortunately, UFH-AT complexes cannot completely inhibit the continuous generation of thrombin during CPB [5,6]. Ongoing thrombin generation contributes to the concomitant generation of several circulating inflammatory proteins, thereby exacerbating the risk for CPB-induced systemic inflammatory response syndrome (SIRS), in addition to CPB-associated thrombosis [7]. An anticoagulation strategy that could reduce thrombin generation during CPB might reduce CPB-associated morbidity. For several years, investigators have identified and explored the use of RNA aptamer anticoagulants as alternatives to heparin for CPB [8–10]. The anticoagulant aptamers identified to date bind a specific exosite on a particular coagulation factor, rather than the catalytic site of their target protease.
They largely inhibit coagulation by inhibiting macromolecular interactions between their target protease and its substrate or cofactor [11–13]. Unfortunately, no anticoagulant aptamer alone has been shown to effect anticoagulation potent enough to facilitate safe CPB [1,14]. However, their mechanism of inhibition provides a unique opportunity for use as adjuvant therapeutics to UFH to potentially address its shortcomings.
The FX/FXa anticoagulant aptamer 11F7t binds a proexosite of FX and the corresponding exosite of FXa with low nanomolar affinity, and 11F7t strongly inhibits FXa from binding to its cofactor, FVa [15]. To lesser degrees, 11F7t also inhibits FXa-catalyzed cleavage of FVIII and FIXa-catalyzed cleavage of FX [15]. Thus, the aptamer uses a much different mechanism to achieve anticoagulation compared to UFH.
Based upon these observations, we hypothesized that aptamer 11F7t might be able to complement UFH and improve its anticoagulant effectiveness in CPB. In this study, we discuss the additive action of the aptamer 11F7t with subtherapeutic doses of UFH in in vitro assays and ex vivo models of CPB. This therapeutic strategy is able to achieve potent and reversible CPB-sufficient anticoagulation associated with minimal thrombin generation compared to UFH alone.
We also studied the possible application of this combinatorial therapy in patients severely ill from COVID-19. These patients can develop systemic thromboinflammatory complications due to SARS-CoV-2-induced tissue damage [16,17]. In addition, some COVID-19 patients require extracorporeal membrane oxygenation (ECMO) due to acute respiratory distress syndrome (ARDS). In these patients, COVID-19-associated coagulopathy (CoAC) is exacerbated further by the additional activation of coagulation through the contact activation system during ex vivo circulation of blood. Our results suggest that the ability of the UFH/aptamer 11F7t combination to limit thrombin generation in highly procoagulant settings may enable it to alleviate coagulopathic effects of COVID-19 and improve outcomes in patients subject to CPB [7,18–23].
Materials and Methods
Anticoagulants and antidotes
The FX/Xa aptamer 11F7t (5′-GAGAGCCCCAGCGAGAUAAUACUUGGCCCCGCUCUU-3′) and the control aptamer 11F7t-Mut4 (5′-GAGAGCCCCAGCGAGAUAAUACUUGUACCCGCUCUU3′) were purchased from Biosynthesis with C denoting 2′-fluorocytosine and U 2′ fluorouracil. Aptamers 11F7t and 11F7t-Mut4 were refolded at 65°C in 20 mM HEPES with 150 mM NaCl and 2 mM CaCl2 at pH 7.4. UFH, enoxaparin and protamine were purchased from the Duke University Hospital Pharmacy.
Blood and plasma
For non-COVID-19 experiments, blood was obtained from healthy human volunteers. For all COVID-19-related experiments, citrated whole blood was obtained from severely ill COVID-19 patients in the intensive care unit (ICU) at Duke Hospital. All study activities were reviewed and approved by the Duke University Institutional Review Board. Citrated platelet-poor plasma (PPP) for the activated partial thromboplastin time (aPTT) and prothrombin time (PT) assays was purchased from George King Plasma Bio-Medical.
Enzyme-linked immunosorbent assay
Enzygnost prothrombin fragment F1 + 2 and thrombin-antithrombin (TAT) enzyme-linked immunosorbent assay kits were purchased from Siemens.
Plasma clotting assays (aPTT, PT)
aPTT and PT were performed on an ST4 mechanical coagulometer from Diagnostica Stago [13]. For aPTT, 50 μL of citrated normal pooled PPP was incubated at 37°C with 50 μL TriniCLOT aPTT S reagent for 5 min (Diagnostica Stago). Thereafter, either 5 μL of 11F7t or HEPES buffer was added, and incubated for 5 min. Subsequently, 50 μL of 0.2 M CaCl2 was added to initiate the assay. For the PT, 50 μL of citrated normal pooled PPP was incubated at 37°C for 5 min, followed by addition of 5 μL of 11F7t or HEPES buffer and a 5-min incubation at 37°C. Thereafter, 100 μL of TriniCLOT PT Excel reagent was added to initiate the assay (Diagnostica Stago).
Activated clotting time
Seventy-two microliters of citrated whole blood was mixed with 2 μL of 11F7t/HEPES buffer, 2 μL of UFH/HEPES buffer, and 2 μL of protamine/HEPES buffer as relevant in the test conditions. Around 2.1 μL of 245 mM CaCl2 was added, and immediately, 72 μL of this mixture was added to an activated clotting time (ACT)+ cuvette to measure the ACT using a Hemochron Jr Signature point-of-care device (Instrumentation Laboratory) [1]. In studies involving COVID-19 patient blood, 2.1 μL of 245 mM CaCl2 was added to 78 μL of blood pretreated with aptamer 11F7t to a final concentration of 2 μM. Seventy-eight microliters of this mixture was then used to determine the ACT as described.
Thromboelastography
Three hundred twenty microliters of citrated whole blood was mixed with anticoagulant (10 μL, in HEPES buffer) and 10 μL kaolin (Haemonetics). This mixture was added to 20 μL of 0.2 M CaCl2 and the assay was run at 37°C according to the manufacturer's instructions (Haemonetics). Clot formation was monitored using a Thromboelastograph Analyzer (Haemonetics) until a stable clot was formed or until 3 h had elapsed.
Rotational thromboelastometry
Citrated whole blood was pretreated with HEPES buffer or aptamer 11F7t (2 μM). EXTEM (extrinsic pathway), INTEM (intrinsic pathway), HEPTEM (INTEM performed in the presence of heparinase), and FIBTEM (fibrinogen only) tests were performed using a ROTEM delta analyzer (Instrumentation Laboratory) according to the manufacturer's instructions. Tests were run using 300 μL of blood and reagents from the manufacturer (Instrumentation Laboratory).
Calibrated automated thrombogram
Citrated whole blood from severely ill COVID-19 patients was centrifuged at 1,800 g for 20 min to prepare PPP. In an Immunlon 2HB clear U-bottom 96-well plate, 70 μL of PPP and 20 μL of the PPP Reagent-High (5 pm TF) were mixed with 10 μL of the HEPES-saline buffer described above or 11F7t (2 μM). The assay was initiated by the addition of 20 μL of the Flu-Ca reagent, which contains a thrombin-specific fluorogenic substrate as well as calcium. Thrombin generation was monitored for 60 min at 37°C by a Fluoroskan Ascent Plate Reader (Thermo Labsystems). The endogenous thrombin potential (ETP), lag time, peak thrombin concentration, and time to peak were automatically calculated by the Thrombinoscope software (Diagnostica Stago).
Ex vivo membrane oxygenator circuit
The miniature ex vivo membrane oxygenator circuit used in this study has been described previously [1,13]. In summary, the circuit consists of a venous reservoir, silicone tubing, and a mechanical roller pump (MasterFlex) along with an oxygenator. Oxygenation was maintained by a gas mixture of 95% O2 and 5% CO2. Approximately 33 mL of blood anticoagulated with 11F7t, UFH, or both was added to the circuit and circulated continuously for 120 min. During the study, samples were taken out for ACT assessment. At the completion of each experiment, the circuit blood was collected and stored for subsequent studies. All stored blood samples were anticoagulated with 3.2% sodium citrate and centrifuged at 3,000 rpm for 10 min to yield PPP.
Illustrations
Illustrations in Supplementary Figs. S1, S3a, and S5 were created using Biorender.com (https://biorender.com/)
Statistical analysis
All graphing and statistical analysis were done using GraphPad Prism 9 software. All data are presented as mean ± scanning electron microscopy unless otherwise stated.
Results
Anticoagulation with aptamer 11F7t complements UFH
To determine if aptamer 11F7t and UFH provide additive anticoagulation, point-of-care coagulation assays, thromboelastography (TEG), and ACT were performed on human whole blood anticoagulated with aptamer 11F7t, UFH, or both anticoagulants (Fig. 1). Concentrations of 1 and 2 μM 11F7t raised TEG clotting time to 22.2 ± 3.5 and 25.1 ± 7.5 min, respectively, from a baseline of 6.7 ± 0.4 min. Concentrations of 0.25 U/mL UFH and 0.5 U/mL UFH raised the TEG clotting time to 38.3 ± 6.7 and 92 ± 27 min. In the concomitant presence of 2 μM 11F7t, both clotting times were elevated to ≥180 min, while 1 μM 11F7t plus 0.25 U/mL UFH prolonged the TEG clotting time to 165 ± 15 and ≥180 min when combined with 0.50 U/mL UFH (Fig. 1A and Supplementary Table S1 and Supplementary Fig. S1A).

Aptamer 11F7t addition enhances UFH anticoagulation of whole blood.
A similar elevation of aPTT and PT clot times (CTs) was observed in normal human plasma when 11F7t was added to very low doses of UFH (Supplementary Fig. S2). These results indicate that aptamer 11F7t and UFH augment each other's anticoagulant activity in human whole blood.
The mutual augmentation of UFH and 11F7t was also observed in whole blood ACT assays performed in the presence of 2 μM 11F7t plus UFH concentrations ranging from 1 to 5 U/mL (Fig. 1B). In CPB, the standard concentration of UFH is 4–5 U/mL (approximately equivalent to 0.5–0.7 μM based on an average molecular weight of 15 kDa) [24,25]. At this concentration, UFH raises the ACT to between 400 and 500 s from a baseline of 90–120 s. An ACT of 455 ± 38 s was observed with the combination 2 U/mL UFH and 2 μM 11F7t from a baseline of 110 ± 6 s (Fig. 1B). In comparison to 5 U/mL UFH, which produced an ACT of 439 ± 20 s, there was no significant difference (P > 0.9999).
These data indicate that the presence of 2 μM 11F7t allows the concentration of UFH needed to achieve an ACT of >400 s to reduce from 5 to 2 U/mL. UFH concentrations higher than 2 U/mL increasingly amplified the increase in ACT produced by the aptamer (Fig. 1B; P ≤ 0.005). The control aptamer 11F7t-Mut4 did not augment the activity of UFH, further confirming that the aptamer 11F7t was solely responsible for the increased anticoagulant activity (Fig. 1C). This observation provided further evidence that this combinatorial anticoagulation strategy might achieve an anticoagulant effect potent enough to facilitate safe CPB, while using less than half the normal amount of heparin required.
Protamine can reverse the anticoagulant effects of aptamer 11F7t and UFH when used in combination
Following discontinuation of CPB, it is important for patient outcomes that normal hemostasis be restored in a timely manner to minimize the risk for postoperative bleeding. As such, it is important that this combinatorial anticoagulation strategy be quickly reversible for optimal use in CPB. Due to the substantial negative charge on both UFH and the backbone of aptamer 11F7t, each forms inactive complexes through electrostatic interactions with positively charged macromolecules [26]. Protamine, a highly basic protein, has been the standard reversal agent for UFH since 1953 [27]. We hypothesized that protamine could simultaneously reverse the activities of both UFH and 11F7t when both agents are present together.
As seen in Fig. 1D, the administration of protamine at the concentration of 0.05 mg/mL to blood anticoagulated by combinations of UFH and 11F7t restored blood clotting to the normal range of non-anticoagulated blood (3–8 min) within 15 min following addition. Similarly, 0.25 mg/mL protamine restored clotting within 12–18 min following addition. These results indicate that protamine treatment can rapidly reverse the anticoagulant activity of UFH and aptamer 11F7t when both are present in blood together. Because most RNA aptamers are negatively charged, we anticipate that protamine would have the same effect as other anticoagulant aptamers, an observation we previously reported [26].
Aptamer 11F7t allows subtherapeutic doses of UFH to maintain ex vivo oxygenator circuit patency
To further investigate whether the concomitant presence of aptamer 11F7t might reduce the concentration of UFH required for CPB anticoagulation, fresh human blood was treated with different 11F7t and UFH combinations and continuously circulated for 120 min in a miniature ex vivo oxygenator circuit (see Materials and Methods section and Supplementary Fig. S3) [1]. During this procedure, flow rate and blood pressure in the circuit were measured continuously to monitor patency of the circuit (Fig. 2A, B).

Aptamer 11F7t enhances UFH efficacy and limits thrombin generation in extracorporeal oxygenation and circulation of human blood.
As expected, in circuits carrying blood anticoagulated with 5 U/mL UFH alone, no significant change from the initial flow rate of 50 mL/min occurred throughout the 120 min of circulation. Mean circuit pressure also remained constant at ∼100 mmHg (Fig. 2A, B). The circuit remained free of visible clot, and electron micrographs (EMs) of the postcirculation oxygenator membrane showed no evidence of fibrin deposition (Fig. 2D). In circuits carrying blood anticoagulated with 2 U/mL UFH, flow rate and pressure also remained stable for 120 min.
However, EMs of post-recirculation oxygenator membranes from those circuits revealed the presence of thrombus deposition (Fig. 2D). As predicted from the TEG and ACT assays, 2 μM 11F7t alone also did not prevent clot formation during circulation. Clots became visible <30 min after circulation was started, the flow rate fell from 50 to <10 mL/min, and mean circuit pressure simultaneously increased from 97.5 ± 5.5 mmHg to over 400 mmHg (Fig. 2A, B).
Postcirculation EMs revealed obvious membrane thrombus deposition (Fig. 2D). In contrast, in circuits containing blood anticoagulated with 2 μM 11F7t plus either 2 U/mL UFH or 5 U/mL UFH, no clot formation occurred throughout the 120 min of continuous circulation period. Flow and pressure measurements mirrored those seen in circuits carrying blood anticoagulated with 5 U/mL UFH alone.
In addition, EMs of postcirculation membranes from circuits conducted with 2 U/mL UFH plus 2 μM aptamer 11F7t showed no evidence of fibrin deposition, unlike EMs of postcirculation membranes from circuits conducted with 2 U/mL UFH alone. EMs of the former membranes were indistinguishable from EMs of membranes from circuits conducted with 5 U/mL UFH (Fig. 2D). These results demonstrate that low-dose UFH can be combined with 11F7t to provide CBP-sufficient anticoagulation as demonstrated by its ability to keep the circuit patent just as 5 U/mL UFH does.
In addition to pressure and flow rate, the ACT of the circulated blood was monitored throughout the procedure to determine if the combinatorial anticoagulation strategy was maintaining CPB sufficient levels of anticoagulation (ACT >400 s). The ACT was monitored throughout circulation, except in circuits where early clot formation occurred. Despite maintaining circuit patency (as shown by steady pressure and flow rate), 2 U/mL UFH alone only increased the ACT from baseline (90–120 s) to 259 ± 17 s, consistent with fibrin depositing on the membrane (Fig. 2C, D). In contrast, when 2 U/mL UFH was combined with 2 μM 11F7t, an average high of 536 ± 22 s was achieved, and the ACT generally remained above 400 s for the duration of the procedure (Fig. 2C).
This result was encouraging, as the treatment with 5 U/mL UFH raised the ACT to 518 ± 69 s and subsequent ACT levels also remained close to 400 s. Therefore, 2 μM 11F7t plus 2 U/mL UFH produced an ACT similar to that produced by 5 U/mL UFH. Finally, addition of 2 μM 11F7t plus 5 U/mL UFH further prolonged the ACT compared to 5 U/mL of UFH alone or 2 μM 11F7t plus 2 U/mL UFH. In addition, administration of 0.05 mg/mL protamine to the blood following circulation brought the ACT down to baseline for circuits treated with 5 U/mL of UFH alone, 2 U/mL of UFH plus 2 μM 11F7t, and 5 U/mL of UFH plus 2 μM, supporting the hypothesis that the combinatorial strategy is readily reversible.
The addition of aptamer 11F7t helps control thrombin generation
One of the main drivers of UFH-associated adverse effects that occur during CPB is UFH's limited ability to effectively inhibit thrombin generation [28]. Serving as an adjuvant therapeutic to UFH, we evaluated if aptamer 11F7t may attenuate that shortcoming [28]. To examine this possibility, we measured the levels of prothrombin cleavage fragment F1 + 2 and TAT complex levels, both of which are markers of thrombin generation, in plasma prepared from blood utilized in the ex vivo circuits. Figure 2E and F show that, as expected, ex vivo continuous circulation of blood stimulates thrombin generation.
Following 120 min of circulation, the concentration of F1 + 2 had risen to 9,909 ± 587 pM in blood anticoagulated with 2 U/mL UFH alone, but to only 1,954 ± 255 pM in blood anticoagulated with 2 U/mL UFH plus 2 μM 11F7t (P < 0.0001) (Fig. 2E). The corresponding postcirculation F1 + 2 values in blood anticoagulated with either 5 U/mL UFH alone or 5 U/mL UFH plus 2 μM 11F7t were 3,382 ± 496 and 1,412 ± 119 pM, respectively (P < 0.05). Thus, addition of the aptamer limits thrombin generation ∼80% and 60% more effectively than UFH 2 and 5 U/mL alone, respectively. Finally, addition of 11F7t to 2 U/mL of UFH gave an F1 + 2 value of 1,953 ± 254 pM versus 5 U/mL of UFH alone, which gave a value of 3,382 ± 496 pM.
Although this difference did not reach statistical significance, this trend suggests that the combination of 2 U/mL UFH plus 2 μM 11F7t may be more effective at limiting thrombin generation that 5 U/mL UFH alone. Nevertheless, these data indicate that the combination of 2 μM 11F7t and a subtherapeutic UFH dose (2 U/mL) is at least as effective as the standard dose of UFH (5 U/mL) at controlling thrombin generation, while addition of aptamer to therapeutic levels of UFH (5 U/mL) is more effective than either anticoagulation strategy at controlling thrombin generation during circulation.
Similarly, the addition of 2 μM 11F7t to varying doses of UFH also resulted in corresponding and significant reductions in postcirculation plasma TAT levels in postcirculation blood (Fig. 2F). These studies reveal that the addition of aptamer 11F7t to UFH inhibits thrombin generation to a significantly greater degree than UFH alone in blood circulated continuously for 120 min within an ex vivo oxygenator circuit.
Aptamer addition limits thrombin generation in COVID-19 ICU patient samples
Subcutaneous (SC) low molecular weight heparin (LMWH) is recommended for thromboprophylaxis in most hospitalized COVID-19 patients, whose hypercoagulable state is marked by elevation of circulating D-dimer and fibrinogen [29]. Reducing thrombin generation in those patients may improve their clinical outcomes as higher thrombin generation has been linked to worse patient prognosis [21]. Those observations and the fact that aptamer 11F7t enhances the anticoagulant potency of UFH led us to test the hypothesis that 11F7t would also enhance the anticoagulant potency of LMWH. In TEG assays performed on healthy human blood anticoagulated with 2.5, 5.0, and 10.0 mg/L of enoxaparin (LMWH), clotting times were 11.6 ± 1.9, 16.5 ± 3.6, and 40.2 ± 6.4 min, respectively, compared to a baseline of 6.1 ± 0.6 min (Fig. 3A).
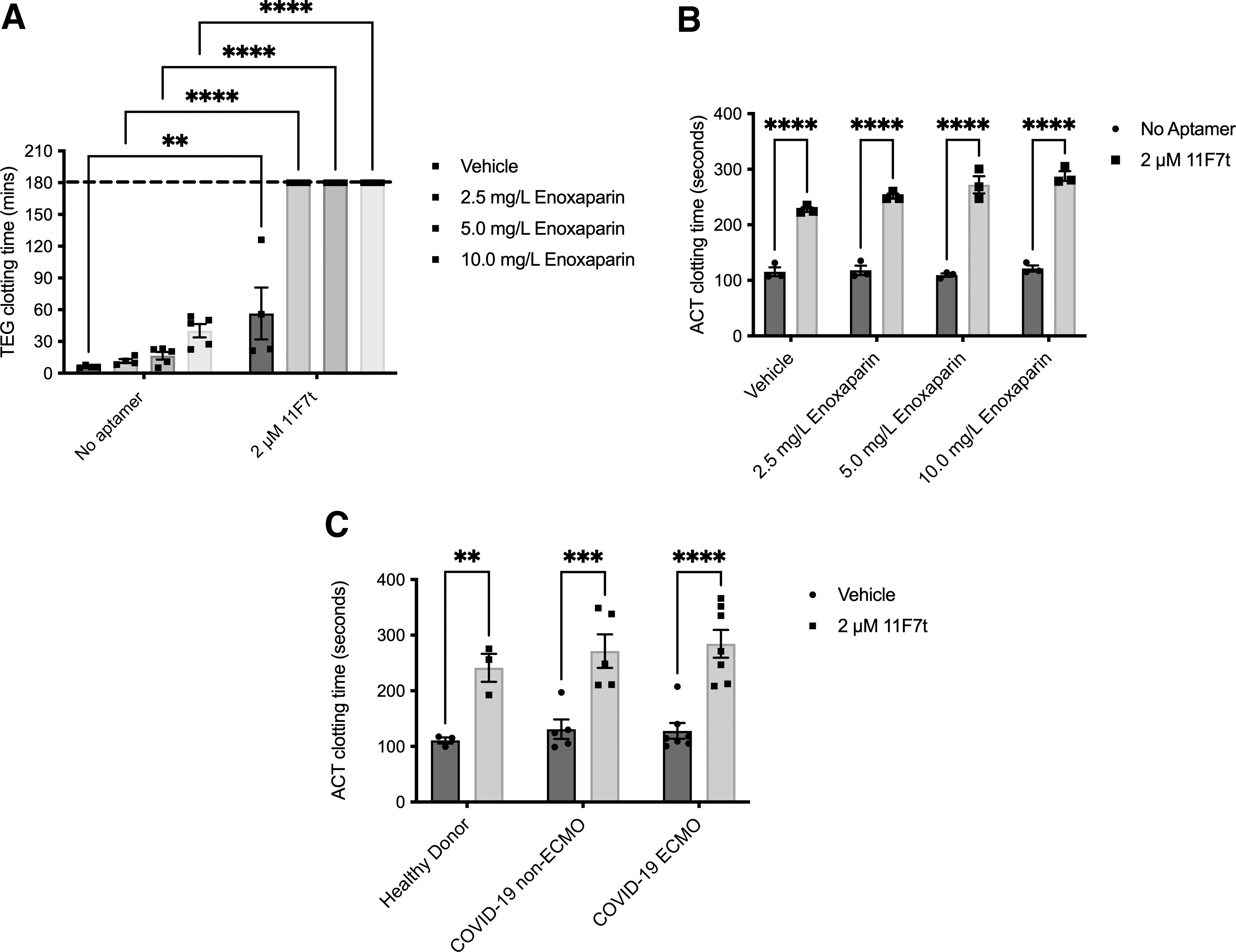
Aptamer 11F7t enhances anticoagulation of enoxaparin (LMWH) and UFH in healthy donor and COVID-19 ICU patient blood.
These concentrations of enoxaparin are approximately equivalent to 0.6, 1.1, and 2.2 μM, respectively, using an average molecular weight of 4.5 kDa [30]. In the concomitant presence of 2 μM aptamer 11F7t, each of those clotting times increased significantly to >180 min. Similar combinatorial anticoagulation was observed in ACTs run using the same concentrations of enoxaparin and 11F7t (Fig. 3B). Thus, the addition of aptamer 11F7t increases the anticoagulant potency of LMWH, as was observed with UFH.
This combinatorial anticoagulation by LMWH and 11F7t in healthy human donor blood shows potential for application in COVID-19 patients. The therapeutic dose of SC enoxaparin in severe COVID-19 is 1 mg/kg (∼15 mg/L or 3.4 μM) every 12 h [31]. Hence, in these healthy donor blood samples, low doses of enoxaparin were seen to be potentiated by 11F7t. This augmentation of enoxaparin activity shows potential to better control thrombin generation in the setting of CoAC. While combinatorial anticoagulation is milder than in combination with low-dose UFH, it may allow the potentiation of low-dose LMWH to the ECMO target ACT of 180–200 s [32–34]. That combination, through its ability to increase the ACT to >200 s, may also comprise a useful alternative to UFH for ECMO anticoagulation in both COVID and non-COVID patients [35].
Thus, we aimed to determine whether the combination of LMWH/UFH plus 11F7t could abrogate aberrant clotting in COVID-19 patient blood. We determined the ACT of whole blood from severely ill COVID-19 patients, each of whom had received UFH or LMWH, in the absence or presence of aptamer 11F7t (2 μM) (Fig. 3C). For comparison, ACT assays in the presence or absence of 11F7t were also performed on blood from healthy donors. Interestingly, for blood from COVID-19-positive patients without any aptamer, CTs were in the normal range, despite those patients having received UFH or LMWH. Dosing for LMWH ranged from 40 to 60 mg daily, dosed once or twice daily. This dosing regimen has been shown to provide and maintain thromboprophylaxis against venous thrombosis [36].
In addition, patients who were on ECMO and treated with UFH were on a continuous infusion. When used in patients not on ECMO, UFH was administered at the prophylactic dose or higher. As such, the inability to increase the ACT above baseline anticoagulation signals a hypercoagulative state. This finding reinforces the fact that COVID-19 patients are hypercoagulable. In the COVID-19 patient group, addition of 2 μM 11F7t to UFH in the blood from ECMO patients led to an increase in clotting time from 134 ± 25 to 292 ± 33 s (P < 0.0001). Similar results were observed in the non-ECMO patient group (primarily on LMWH). That increase was consistent with 11F7t's previously established effect in normal human blood when combined with LMWH or UFH.
To characterize further if aptamer 11F7t addition may help counteract COVID-19-induced hypercoagulability, we performed rotational thromboelastometry (ROTEM) assays on blood from heparinized COVID-19 patients. The ROTEM is a viscoelastic assay that helps to determine the contributions of different procoagulant stimuli to clot formation (Supplementary Fig. S1B). To this end, several tests are run focused on different parts of the coagulation cascade. In the INTEM test, coagulation is triggered by ellagic acid to focus on thrombosis and fibrinolysis through the intrinsic pathway of coagulation.
It is sensitive therefore to anticoagulation affecting the intrinsic pathway and can detect the presence of UFH in patient blood. In the HEPTEM test, in addition to ellagic acid, heparinase is also added in the assay to eliminate any perturbation due to UFH presence. In combination, these tests help us assess the effect of each anticoagulant (11F7t or UFH) alone or in combination. Different parameters in these tests detect how different parts of the coagulation cascade are affected, giving insight on the overall thrombotic state of a patient (Supplementary Fig. S1).
As shown in Fig. 4A and in Supplementary Table S2, ROTEM assays performed on COVID-19 patient blood displayed shorter CT, shorter clot formation time, greater maximum clot firmness, and greater alpha angle measurements than assays performed on blood from healthy controls, as is typical with patients in a hypercoagulable state [37]. Assays performed on blood from both ECMO and non-ECMO COVID-19 patients showed changes in parameters reflecting this hypercoagulable state. Baseline CT in the INTEM tests in COVID-19 ECMO patients was higher (244 ± 39 s) compared to the baseline CT in the HEPTEM (223 ± 16 s), signaling the presence of either LMWH or UFH in the patient blood (Fig. 4B).
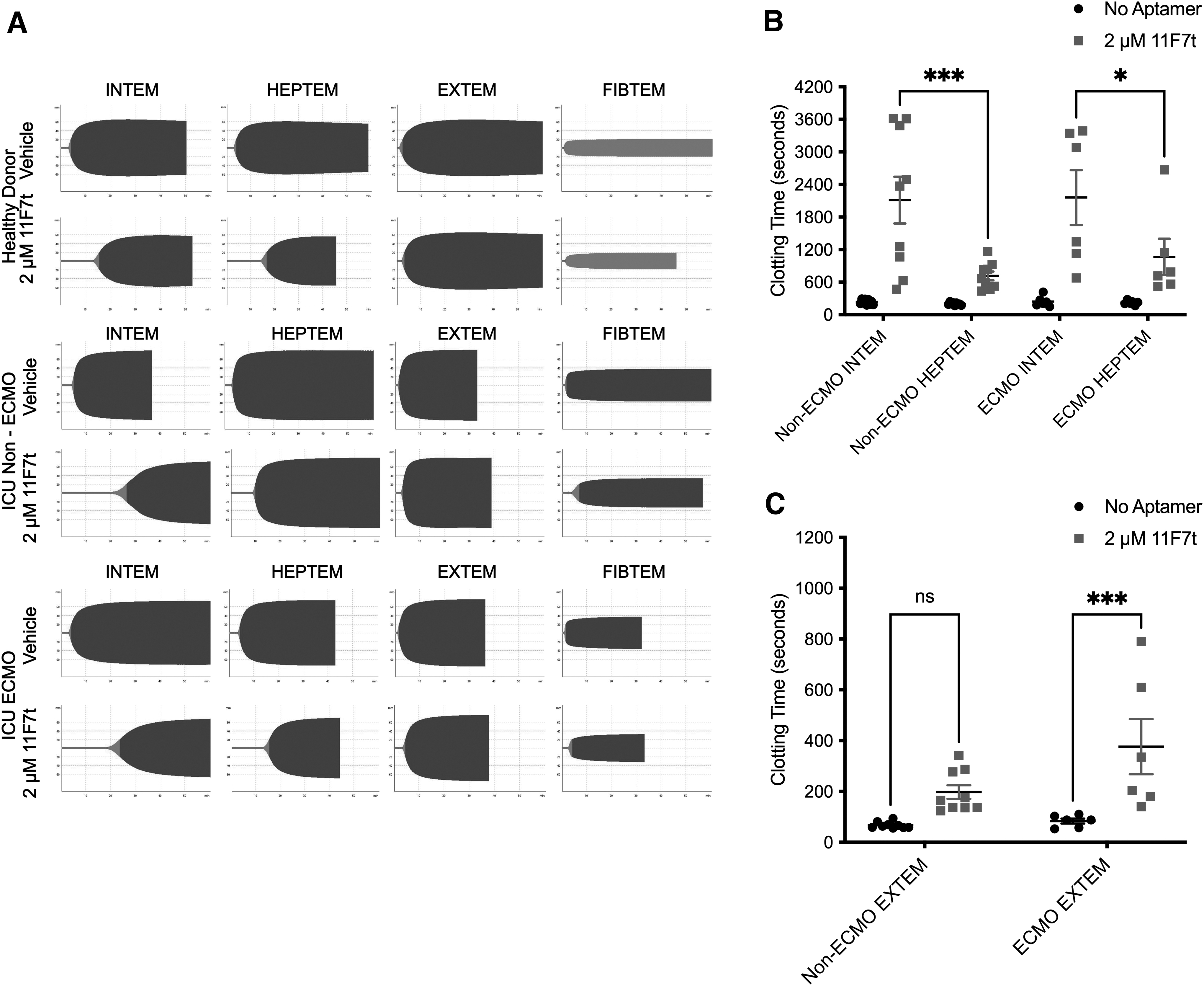
Aptamer 11F7t enhances anticoagulation by UFH and LMWH in COVID-19 ICU patient blood as measured by ROTEM.
This difference was amplified with the addition of aptamer 11F7t (2 μM) to blood from COVID-19 ECMO patients where the CT in the HEPTEM was 1,067 ± 333 s (effect of 11F7t alone) compared to CT in the INTEM 2,160 ± 506 s (effect of 11F7t plus LMWH or UFH in patient blood) (P < 0.05). Aptamer 11F7t produced similar, although less marked changes in CT in the EXTEM test (Fig. 4C). In this test, coagulation is triggered by tissue factor, thereby allowing assessment of thrombosis and fibrinolysis through the extrinsic pathway. In all these studies, the combinatorial anticoagulation produced by UFH and 11F7t is greater than between LMWH and 11F7t, as shown in Supplementary Fig. S4.
Next, we aimed to determine if aptamer addition translated to an improved inhibition of thrombin generation in blood samples from severely ill COVID-19 patients. We prepared PPP from five COVID-19 ICU patients who were receiving prophylactic LMWH or therapeutic UFH (one patient on ECMO). Using the calibrated automated thrombogram (CAT) assay, we compared thrombin generation potential in those samples in the presence and absence of aptamer 11F7t (Fig. 5 and Supplementary Fig. S5) [1,38]. We observed extensive thrombin generation in these patient samples, similar to that in non-anticoagulated blood, despite the presence of UFH or LMWH, as has been observed in other studies [20]. The CAT assay revealed that the addition of aptamer 11F7t significantly reduced thrombin generation in PPP from COVID-19 patients.

Addition of aptamer 11F7t significantly reduces thrombin generation potential in heparin-treated COVID-19 ICU patient blood. Calibrated automated thrombogram assays were performed to measure thrombin generation in platelet-poor plasma obtained from severely ill COVID-19 patients. All patients were treated with enoxaparin (n = 4) or UFH (n = 1) the day before blood isolation.
The ETP dropped from an average of 2,547 ± 192 nM*min down to 0 nM*min (below detection) for four patients and to 1,901 nM*min for one patient (Fig. 5 and Supplementary Table S3). Corresponding changes in other parameters from the assay such as a longer lag time, longer time to peak, and a lower peak of the curve were observed (Fig. 5A and Supplementary Table S3). Those findings support the hypothesis that addition of aptamer 11F7t to either UFH or LMWH can significantly limit thrombin generation in COVID-19 patient samples, which if utilized clinically should help reduce the severity of COVID-19-associated coagulopathy and associated comorbidities [21].
Discussion
Several coagulation assays (Fig. 1 and Supplementary Fig. 2) reveal that the addition of aptamer 11F7t to UFH or LMWH augments each's anticoagulant effects in human blood samples. In addition, the combination of 2 U/mL UFH plus 2 μM 11F7t prevented both macroscopic and microscopic clot formation in human blood circulated continuously for 2 h within an ex vivo oxygenator circuit. Neither anticoagulant alone, at those concentrations, prevented thrombus formation in that setting.
Protamine reversed the anticoagulant effects of both agents. Levels of prothrombin fragment F1 + 2 and TAT in postcirculation circuit blood were significantly lower if aptamer 11F7t was added to UFH and used for anticoagulation compared to UFH alone. Thus, aptamer 11F7t's ability to increase the anticoagulant potency of UFH in human blood appears to result from its ability to reduce the level of thrombin generation that occurs despite the presence of UFH [3,6,39–41].
This combinatorial anticoagulation strategy may provide clinical benefits in other procoagulant settings in which UFH and LMWH comprise the standard-of-care anticoagulation strategy. The hypercoagulability of COVID-19 patients leaves many vulnerable to life-threatening complications, despite anticoagulation [42,43]. This threat is even more serious in patients with COVID-19-induced ARDS, who require ECMO [44,45]. Treatment with UFH or LMWH has become standard-of-care for most hospitalized COVID-19 patients, in whom morbidity and mortality frequently result from thromboembolic complications [46].
The TEG and ACT clotting times of whole blood from healthy donors indicate that aptamer 11F7t augments the anticoagulant effects of LMWH (Fig. 4). Consistent with those observations, ACT and ROTEM assays performed on blood from COVID-19 patients treated with either UFH or LMWH further demonstrated the ability of aptamer 11F7t to augment the anticoagulant effects of those agents (Fig. 5).
In addition, CAT assays performed on PPP from COVID-19 ICU patients, who were receiving either UFH or LMWH, showed that the addition of aptamer 11F7t significantly reduced thrombin generation (Fig. 5). Thus, aptamer 11F7t augments the anticoagulant potency of both UFH and LMWH and reduces thrombin generation in blood from severely ill, hospitalized COVID-19 patients. Whether aptamer 11F7t's ability to reduce thrombin generation can reduce the severity of systemic thromboinflammation in COVID-19 patients receiving UFH or enoxaparin (LMWH) will require further study in the clinic.
The addition of 11F7t to UFH yields an anticoagulation strategy that is potent and reversible and reduces thrombin generation, and therefore has potential to address several CPB-associated adverse effects. This strategy addresses several limitations of using UFH alone and other anticoagulants that have been considered alternatives to UFH in CPB. Examples of these include bivalirudin, a synthetic peptide inhibitor of thrombin, which carries a short half-life of 25 min. In a clinical trial comparing UFH and bivalirudin for pediatric CPB, achieving a steady target ACT was more problematic with bivalirudin [47]. More importantly, bivalirudin limits thrombin generation significantly less effectively than UFH [48].
Thus, bivalirudin has no advantage over UFH for CPB anticoagulation in patients who have no contraindication to UFH. Another alternative that has been explored is the combination of direct oral anticoagulants (DOACs) targeting FXa in combination with aptamer 11F7t [1]. Unfortunately, no FXa DOAC is approved for IV use, and the high cost of the only approved FXa DOAC antidote, andexanet alfa, makes it unsuitable for standard use in the setting of CPB [49–51]. As a result, despite its limitations, UFH remains the safest, most well controlled, and potent anticoagulant for use in CPB.
With this new therapeutic strategy, we allow UFH therapy to be more effective. By further limiting thrombin generation during UFH administration, there is potential to limit the development of SIRS in patients during CPB [7]. Due to UFH and other heparinoids' widespread use in anticoagulation, this aptamer-based therapeutic approach holds promise across several applications.
With the ongoing COVID-19 pandemic, some patients develop severe ARDS in which mechanical ventilation becomes insufficient, requiring them to receive ECMO treatment. Both CoAC and ECMO lead to a systemic hypercoagulative and inflammatory state, leaving patients at a high risk of thrombotic events as seen in several studies [32,34]. In this study, we observed that the addition of aptamer 11F7t to UFH or LMWH in blood from severely ill patients (on ECMO and not) improved anticoagulation and showed potential for reduced thrombin generation.
In addition, the use of 11F7t as an adjuvant therapeutic would also work to address the dose–response variability that is observed with UFH use. Due to the heterogenous nature of UFH, there is high variability in patients' response to the therapeutic [6]. Also, as we suggested earlier, 11F7t could potentiate LMWH such that it can be used to replace UFH in highly procoagulant settings that it is currently not applicable due to limited anticoagulant activity. Moreover, concerns about UFH shortages remain; an outbreak of African Swine Flu in 2018–2019 affecting 25%–35% of the pig herds in China during the years 2019 and 2020 caused a shortage of UFH in the United States [52,53].
As such, by reducing the dose of UFH needed to achieve CPB-sufficient anticoagulation as observed in our studies, we provide an aptamer-based contingency approach or potential universal anticoagulation strategy that would allow preservation of UFH supplies in the face of shortages. This approach also holds promise in alleviating potential shock on UFH/LMWH supplies during the ongoing COVID-19 pandemic.
Another complication of UFH therapy that the 11F7t aptamer has the potential to alleviate is heparin-induced thrombocytopenia (HIT). In this illness, heparin molecules complex with platelet factor 4 forming an immunogenic complex leading to aberrant activation and consumption of platelets, leaving the patient in a hypercoagulable state. The incidence of HIT correlates with the dose of UFH used, hence limiting how much UFH patients are exposed to by complementing with the aptamer will help lower the risk of developing HIT [54].
The findings of this study suggest that during human CPB, the addition of aptamer 11F7t to UFH at normal or reduced doses might limit thrombin generation more effectively than UFH alone and that this strategy has potential utility across other indications. Finally, they suggest that anticoagulant aptamers may much more quickly find a home in the clinic by improving an existing anticoagulant strategy, based upon the use of heparin, rather than immediately fully replacing this 68-year old, widely utilized approach. We anticipate that given their mechanism of inhibition, other aptamers will find clinical value augmenting other classes of therapeutic agents as well.
Footnotes
Acknowledgments
The authors would like to acknowledge J. Levy, G. Arepally, T. Ortel, J. Layzer, and G. Pitoc for useful discussion and guidance during this study.
Author Disclosure Statement
Duke has submitted patent applications on the factor X/Xa anticoagulant aptamer(s) and combinations of the aptamer with other anticoagulants. Dr. Sullenger and Dr. Frederiksen are inventors on such Duke Intellectual Property.
Funding Information
This work was supported by a US National Institutes of Health grant (Grant No. P01-HL139420).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.