
Abstract
The severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) has caused the current worldwide pandemic and the associated coronavirus disease 2019 with potentially lethal outcome. Although effective vaccines strongly contributed to reduce disease severity, establishing a toolbox to control current and newly emerging coronaviruses of epidemic concern requires the development of novel therapeutic compounds, to treat severely infected individuals and to prevent virus transmission. Here we present a therapeutic strategy targeting the SARS-CoV-2 RNA genome using antisense oligonucleotides (ASOs). We demonstrate that selected locked nucleic acid gapmers have the potency to reduce the in vitro intracellular viral load by up to 96%. Our promising results strongly support the case for further development of our preselected ASOs as therapeutic or prophylactic antiviral agents.
Introduction
Mankind is currently dealing with the third outbreak in 20 years of an emerging human-infecting coronavirus, the severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) [1]. The pandemic was partially contained due to the successful development of vaccines, especially the newly emerged messenger RNA (mRNA)-based vaccines [2]. However, antiviral drugs are and will be needed to complement vaccination in the clinical arena, for example, for patients with severe chronic infections, to serve as prophylactics for high-risk groups, or to control future outbreaks of novel coronaviruses.
Multiple attempts to repurpose existing drugs resulted in few candidates that are currently used in clinics (eg, Paxlovid, Molnupiravir [3]). Other therapeutic strategies have been developed particularly aiming to reduce viral transmission, including “miniproteins” [4], lipopeptides [5], antibody cocktails [6], or small-molecule drugs that target viral components [7].
We took a parallel approach to develop antisense oligonucleotides (ASOs) that specifically target SARS-CoV-2 viral RNA, aiming to block viral replication. SARS-CoV-2 is a positive-sense single-stranded RNA virus with a genome of ∼30 kb [8] and well-described structural and functional RNA elements [9–11]. Given the structural architecture of the SARS-CoV-2 RNA genome, we specifically selected (un)structured elements that can serve as ASO binding site and that were known to have functional roles in viral replication (Fig. 1A) [12–14].

In vitro screening of ASOs targeting the SARS-CoV-2 genome in a cell-based assay.
We designed multiple viral RNA-targeting ASOs of different chemistries (locked nucleic acid gapmers [LNA gapmers] [DNA ASOs with RNA-like segments on both sides], LNA-2’Ome gapmers, 2’Ome gapmers, 2’Omes, 2′-O-methoxyethyl [MOEs], MOE gapmers). Gapmers [15] are powerful tools for mRNA loss-of-function studies as they catalyze the RNase H-dependent degradation of complementary RNA targets, whereas RNA ASOs are expected to interfere through sterical hindrance [16]. The LNA gapmers (containing the LNA flanking regions) are presumed to be superior to other gapmers, as the LNA region increases target binding affinity and confers nuclease resistance [15].
Here we present the selection and in vitro evaluation of new antiviral ASOs against SARS-CoV-2, with the potential of developing a new therapeutic appproach and prevention options for future emerging coronaviruses of pandemic concern. Our findings are in line with much further developed LNA ASO therapeutics [17] and with numerous siRNA therapeutics developed against SARS-CoV-2 [18–22].
Materials and Methods
Molecular cloning
Firefly luciferase reporter constructs were generated by inserting appropriate viral genome segments into the pCI-neo-FLuc (exact nucleotide positions indicated in Figs. 1B and 3A).
Antisense oligonucleotides
All ASOs (containing fully modified phosphorothioate backbones) were produced by Eurogentec. ASO sequences and target locations in the SARS-CoV-2 genome are specified in Supplementary Table S1 and depicted schematically in Fig. 1A.
Cell culture and transfections
Vero E6 cells were cultured as described [23]. For luciferase assays, cells were Lipofectamine transfected with the reporter, control plasmid, and ASO, readings were performed 30 h later (SpectraMax i3x; Molecular Devices). To assess antiviral activity, cells were infected with SARS-CoV-2 for 1 h at 37°C with 12,000 Plaque forming unit (PFU)/well, followed by a 16-h ASO treatment. Multiplex real-time reverse transcription PCR (qRT-PCR) analysis was performed using Taqman probes against the SARS-CoV-2 RdRp gene, whereas the mRNA of the phosphoglycerate kinase 1 (PGK1) housekeeping gene was used for normalization.
Results and Discussion
To facilitate ASO selection, we conducted the initial functional evaluation in Vero E6 cells transfected with reporter gene constructs expressing Firefly luciferase in combination with specific SARS-CoV-2 genome segments. The viral sequences included in the reporter mRNAs are depicted in Fig. 1B. Each reporter construct tested resulted in the selection of ASOs reducing the level of luciferase in comparison with no ASO/ctrlASO (scrambled sequence) controls (Fig. 1C). At this time point, different ASO chemistries were tested and the LNA gapmers seemed to be more potent than the LNA-2’Ome or 2’Ome gapmers.
To exclude potential effects resulting from ASO toxicity, cell viability assays were performed in parallel with this experiment (Supplementary Fig. S5). Testing the most potent ASOs targeting the SARS-CoV-2 5′ untranslated region (UTR) in the same assay set-up in Calu-3 cells (a human lung cancer cell line) resulted in a comparable reduction of luciferase activity (Supplementary Fig. S1). SARS-CoV-2 5′ UTR-targeting lead candidate ASOs were tested to select the most potent gapmer sequence. The arrangement of 10–13 nt central DNA strand flanked by the four LNA nucleotides from the 5′- and the 3′-ends was the most optimal (Supplementary Fig. S2).
To evaluate the level of interference with virus replication of the ASOs selected in the reporter assays, we performed experiments using SARS-CoV-2-infected Vero E6 cells. Initially we used the commonly used experimental set-up in which cells are first pretreated with ASOs before being infected with SARS-CoV-2 (Supplementary Fig. S3). Next we switched to experimental set-up where Vero E6 cells are first infected with SARS-CoV-2 for 1 h, followed by virus removal from the medium and the ASO treatment of the infected cells for 16 h. Intracellular RNA was assayed for the reduction of viral RNA loads in qRT-PCR using primers targeting the viral RdRp and E protein genes.
Using both approaches, the 5′ UTR-targeting ASOs 38 and 44 as well as the N gene-targeting ASOs 41 and 42 were identified as the most potent, leading to a 53%–87% reduction of the intracellular viral RNA load when used alone (depicted in Fig. 2A; data using primers targeting the viral E gene are depicted in Supplementary Fig. S4A). Using any combination of these ASOs, their potency was retained, and the effect was even strengthened, leading up to a 96% reduction of the intracellular viral RNA load (Fig. 2B; Supplementary Fig. S4B). The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay demonstrated no effect of the ASOs used on cell viability.
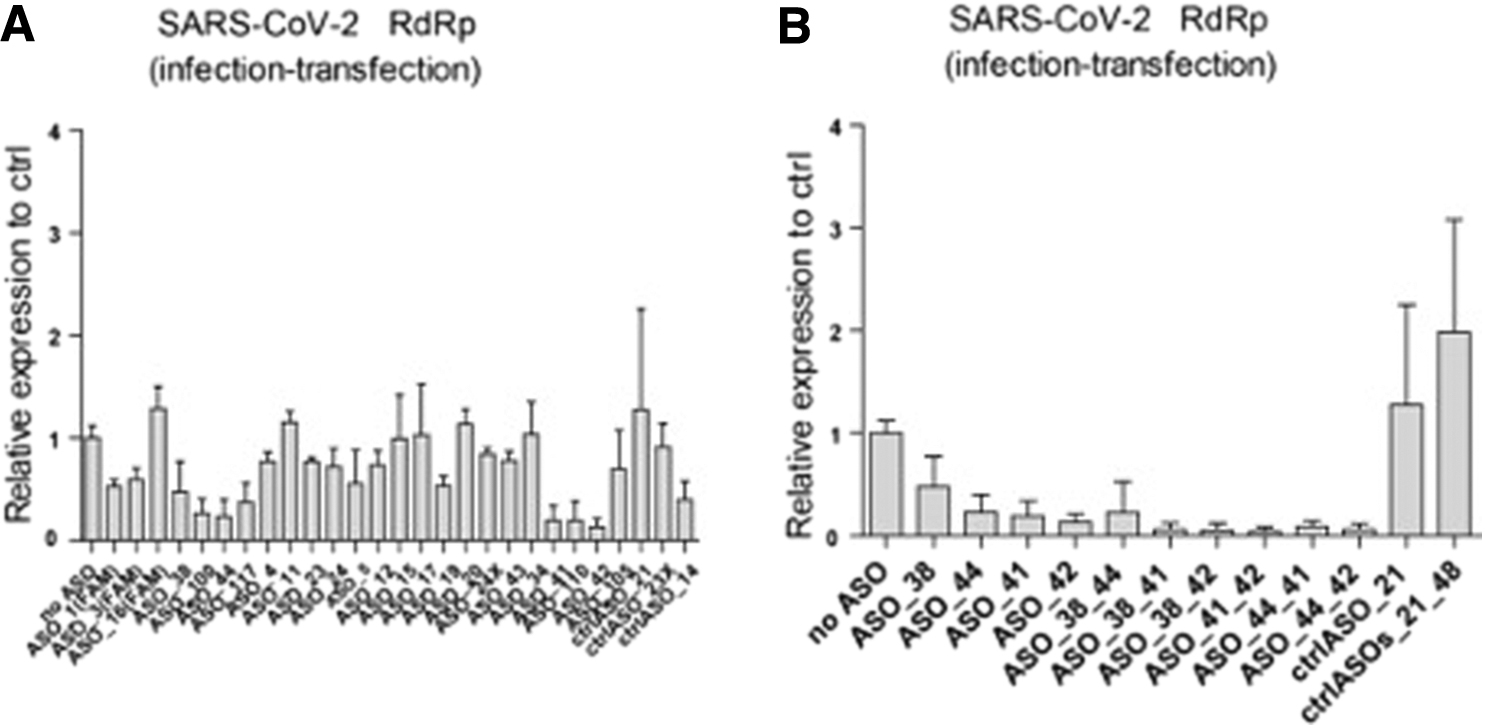
Selected ASOs are potent in reducing intracellular viral RNA levels.
Additional reporter constructs have been generated to assay the potency of lead candidate ASOs against highly conserved sequences from other betacoronaviruses (Fig. 3B). To evaluate the potential for broad-spectrum activity, the lead candidate ASOs targeting the SARS-CoV-2 5′ UTR region were tested using constructs carrying 5′ UTR sequences from SARS-CoV, Middle East respiratory syndrome coronavirus (MERS-CoV), and mouse hepatitis virus (MHV). Figure 3C shows that lead candidate ASOs 38 and 44 are most active against SARS-CoV sequences and in addition ASO 44 showed potency against MHV sequence, but no significant effect was obtained against the MERS-CoV sequence.
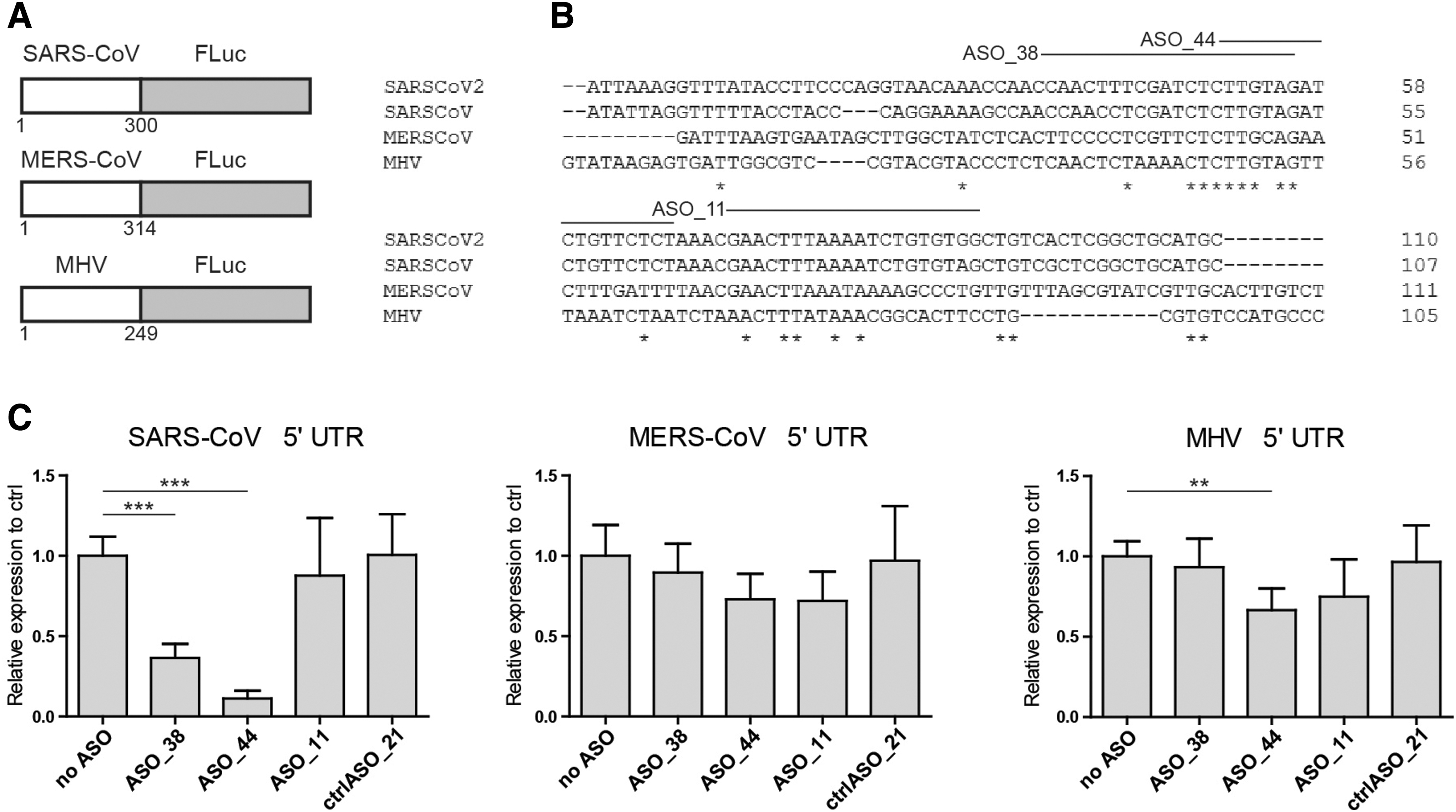
In vitro screening of SARS-CoV-2 genome-targeting ASOs against the other SARS-related betacoronaviruses.
Based on the in vitro results presented here, we expect that ASOs may be effective in reducing virus replication in vivo. Likewise, our in vitro data suggest that ASOs might be effective against other emerging coronaviruses of pandemic concern. In addition, testing of already identified highly conserved weakly structured regions within viral RNA genomes might aid the development of antiviral ASO therapeutics [12].
Work still needs to be done to define the best administration method (intranasal or systemic) and the uptake in the upper and lower respiratory tract, in the systemic circulation and organs. Selected ASOs need to be evaluated for the efficacy in vivo, stability and toxicity. Operating by a different mechanism, ASOs could be envisioned as additives to the existing/in development antivirals for intranasal prophylactic approach, preventing viral transmission.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.