
Abstract
In a previous study using male rats, a correlation was found between the development of “at-level” allodynia in T6-7 dermatomes following severe T8 spinal contusion injury and the sparing of some myelinated axons within the core of the lesion epicenter. To further test our hypothesis that this sparing is important for the expression of allodynia and the supraspinal plasticity that ensues, an injury that severs all axons (i.e., a complete spinal cord transection) was made in 15 male rats. Behavioral assessments were done at level throughout the 30-day recovery period followed by terminal electrophysiological recordings (urethane anesthesia) from single medullary reticular formation (MRF) neurons receiving convergent nociceptive inputs from receptive fields above, at, and below the lesion level. None of the rats developed signs of at-level allodynia (versus 18 of 26 male rats following severe contusion). However, the terminal recording (206 MRF neurons) data resembled those obtained previously post-contusion. That is, there was evidence of neuronal hyper-excitability (relative to previous data from intact controls) to high- and low-threshold mechanical stimulation for “at-level” (dorsal trunk) and “above-level” (eyelids and face) cutaneous territories. These results, when combined with prior data on intact controls and severe/moderate contusions, indicate that (1) an anatomically incomplete injury (some lesion epicenter axonal sparing) following severe contusion is likely important for the development of allodynia and (2) the neuronal hyper-excitability at the level of the medulla is likely involved in nociceptive processes that are not directly related to the conscious expression of pain-like avoidance behaviors that are being used as evidence of allodynia.
A number of studies have shown a positive correlation between the presence/absence of “at-level” allodynia post-contusion injury and neuronal hyper-excitability to innocuous and noxious mechanical stimulation in at-level dermatomes both within the dorsal horn immediately rostral to the injury (wide dynamic range/spinothalamic neurons [Drew et al., 2001; Crown et al., 2005]) and supraspinally in the medullary reticular formation (MRF) (Hubscher and Johnson, 1999a) and thalamus (Gerke et al., 2003; Hubscher and Johnson, 2006). In a recent study using male rats, we also found a positive correlation between both the behavioral and electrophysiological (thalamic recordings 30 days post-injury) evidence of allodynia and the sparing of some myelinated axons within the core of the lesion epicenter (Hubscher and Johnson, 2006).
In the present study, an injury that severs all axons (i.e., a complete spinal cord transection) was made to further test our hypothesis that this core epicenter sparing is important for the expression of allodynia and the supraspinal plasticity that ensues. Fifteen male Wistar rats, 90 days of age, were completely spinalized at the T8 spinal level 30 days before terminal electrophysiological experiments. Animals were anesthetized with a mixture of ketamine (80 mg/kg) and xylazine (10 mg/kg), injected intraperitonealy. Our standard SCI surgery procedures included administration of 0.5 mL of dual penicillin (Ambi Pen®; The Butler Company, Columbus, OH) prior to surgery, 5 mg/kg gentamicin (Abbott Laboratories, North Chicago, IL) once per day for 5 days post-surgery (to control for possible bladder infection) and 2.5 mg/kg of ketoprofen (Ketofen®; Fort Dodge Laboratories, Fort Dodge, IA) once per day for 2 days to alleviate post-surgical pain.
All surgeries were done under aseptic conditions, and the body temperature was controlled throughout the surgery and recovery period. A T7 laminectomy was performed to expose the underlying T8 spinal cord, and the dura was incised and reflected laterally. The spinal cord was cut using a pair of surgical microdissecting scissors. Gentle suction was used as needed to elevate the cut stump to verify the completion of the lesion (Kaddumi and Hubscher, 2007). Gelfoam soaked in thrombin was placed in the lesion cavity. The incision was closed using 4-0 nylon suture for the muscle layers and fascia, and surgical clips for the skin. The urinary bladder was expressed every 8 h.
Beginning 2 weeks post-injury (the period during which SCI pain is known to develop), the dorsolateral trunk was examined for sensitivity to mechanical stimulation in the at-level (T5-T8) dermatomes (Hubscher and Johnson, 2006) by stroking with a no. 5 paint brush (1.5 × 0.5 cm bristles, with an average pressure of 17 g). Avoidance behaviors, including efforts to escape or grab/push the stimulus probe away with the forelimbs (with or without vocalization), were taken as evidence of an allodynia-like response. An animal was considered sensitive to touch if it responded to at least 60% of the stimuli in a given session (minimum of five stimuli) for at least two sessions beyond the initial 2 weeks (Hubscher and Johnson, 2006). All avoidance responses to brush, if present, were assessed further for threshold values using a set of Semmes-Weinstein monofilaments (20 filament set, 15 of which are in the range of 0.008–15 g; obtained from Stoelting Co., Wood Dale, IL).
At 30 days post-transection, each animal was anesthetized with 50% urethane (1.2 g/kg), and the jugular vein, carotid artery, and trachea were exposed and intubated for anesthetic supplement (5% urethane, as needed), blood pressure monitoring, and respiratory rate/end expired pCO2 level monitoring, respectively. Body temperature was monitored throughout the experiment by an esophageal probe connected to a thermometer and maintained at around 37°C using a circulating water-heating pad. After mounting the animal onto a stereotaxic device, a dorsal incision was made to gain access to the brainstem. The dorsal surface of rostral medulla was exposed by removing part of the occipital bone and suctioning the caudal midline portion of the overlying cerebellum (Hubscher and Johnson, 1996, 1999b).
A tungsten microelectrode with ≈7 ± 1 MOhms impedance (Fredrich Haer and Co., Bowdoinham, ME) was lowered from the dorsal surface of the brainstem with a motorized drive (Fredrich Haer and Co.) into the MRF. Stereotaxic coordinates were 3400 μm rostral to obex, and 400 and 800 μm lateral to midline on both sides of the brainstem (two tracks per animal collectively). The search area for each dorso-ventral track was 2800–3000 μm in length, which covered the rostral part of the nucleus reticularis gigantocellularis (Gi), Gi pars alpha (GiA), and the medial part of the lateral paragigantocellular nucleus (LPGi) (Hubscher and Johnson, 1999a).
To insure, as in our previous studies, that the same population of somatovisceral convergent neurons were being sampled in our current experiments, pinching the ear was used as our search stimulus (many of these neurons respond to stimulation of the entire body surface, and 100% respond to ear pinch, as previously described [Hubscher and Johnson, 1999a, 2006]). Recording of a single neuron was established by monitoring the action potential on an oscilloscope with a spike-triggered analog delay module for discrimination of somato-dendritic neuron profiles from nerve fiber spikes as described previously (Hubscher and Johnson, 1996, 1999a, 2006). Once a single neuron responsive to ear pinching was found, responses to mechanical (stroke and pinch) stimuli of the dorsal trunk and to other above level somatic areas (eyelids and face) were tested. MRF neuronal responses were recorded to videotape and analyzed offline using Data Wave software (
At the end of the experiment each animal was perfused transcardially using 0.9% normal saline followed by 4.0% paraformaldehyde. Brainstems were removed and sectioned at 100 μm thickness on the vibratome and stained with cresyl violet. The electrophysiological tracks were identified in these sections under the light microscope and the location of each track was confirmed (Hubscher and Johnson, 1999a, 2006) and reconstructed (Paxinos and Watson, 1998). The spinal cord containing the lesion area was extracted from the transected animals and sectioned sagittally at 18 μm thickness in the cryostat and stained with both luxol fast blue and cresyl violet (Kluver-Barrera Method). The sections were viewed under the light microscope to confirm the completion of the spinal cord transection. Data was analyzed for significance using chi-square test with p < 0.05 level of significance. All animal procedures were reviewed and approved by the Institutional Animal Use and Care Committee, University of Louisville School of Medicine.
None of the 15 rats met our avoidance criteria that would indicate sensitivity to touch (allodynia) “at level,” as tested up to 30 days following transection. One of the 15 rats, however, vocalized to an 8- and 60-g monofilament (left and right side at level, respectively), but did not show any avoidance responses. Four of the 15 rats showed evidence of excessive grooming below level beginning 2–3 weeks post-injury. Three had very small spotty patches of sores, one rat on just the abdomen (bilateral), one rat on the abdomen and inner thigh (left side only), and the other rat on just the lower abdomen and inner thigh (bilateral). The fourth rat had two large sores angled from midline outward on the ventral side of the body on the left side only; one close to but below the lesion level transitional zone (T8-9 region), which measured 3.0 × 0.5 cm, and the other caudal to that (T11-12 region; 2.5 × 0.5 cm).
A total of 206 MRF neurons were found responsive to the search stimulus, pinching the ear. The neuronal characteristics of these neurons did not differ significantly (χ2, p > 0.05) from intact controls (Kaddumi and Hubscher, 2006). Most neurons were excitatory (73.2%) and many had spontaneous discharges at rest (36%), with a mean activity rate of 17.5 ± 1.3 spikes/sec. There was a significant (χ2, p < 0.05) increase in the percentage of MRF neurons responding to mechanical stimulation of different somatic areas (face, eyelid, and dorsal trunk above the level of spinal cord transection) compared to our previous findings using intact rats. In addition, there was a significant increase in the percentage of MRF neurons responding to just low-threshold levels of stimulation (touch/gentle pressure) for all above level areas tested. A summary of the MRF neuronal responses to stimulation of the different somatic regions in normal versus spinalized animals is presented in Table 1. Examples of responses recorded in two MRF neurons to high- and low-threshold stimuli are shown in Figure 1.
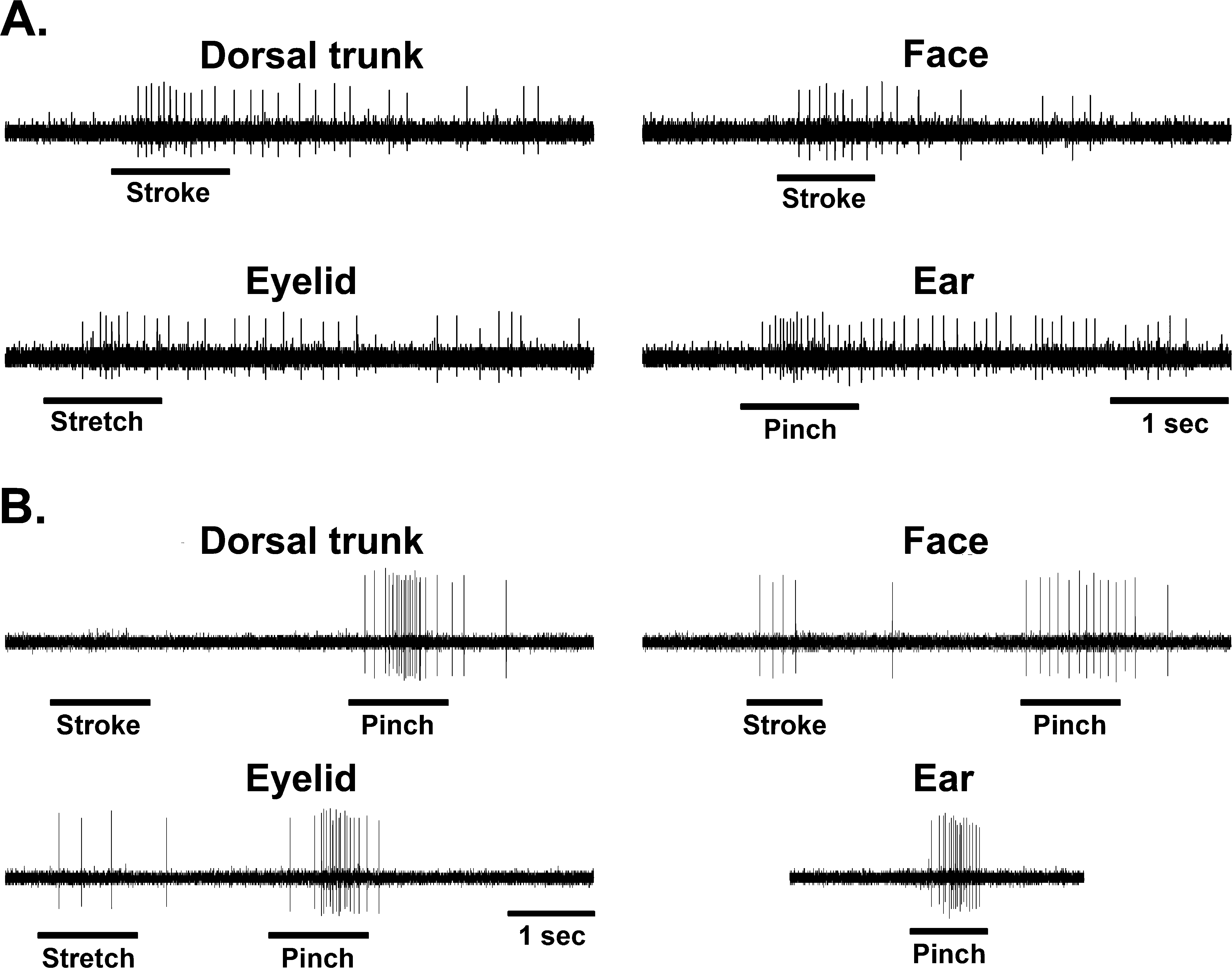
Typical examples of stimulus-response characteristics in medullary reticular formation (MRF) neurons from spinally transected animals. (
Kaddumi and Hubscher, 2006.
Significantly different (χ2, p < 0.001) than normal.
Significantly different (χ2, p < 0.025) than normal.
The 24.8% increase in the proportion of dorsal trunk touch-responsive MRF neurons (indicating hyper-excitability) after spinal cord transection is similar to the 29% increase seen previously in the MRF of spinally contused rats showing signs of allodynia (Hubscher and Johnson, 1999a). Our spinally transected rats, however, did not show any behavioral signs of allodynia, while 69% of the contused animals developed allodynia after injury (Hubscher and Johnson, 1999a, 2006). These results support the idea that at least some intact spinal axons traversing the site of injury are a requisite for the development and maintenance of at-level allodynia (Johnson and Hubscher, 2002; Hubscher and Johnson, 2006). Although chronic transitional zone pain is more frequent in patients with “clinically complete” injuries (Beric, 1999), as is the case for rats with “severe” contusions (Hubscher and Johnson, 1999a; Lindsey et al., 2000), studies show (Finnerup et al., 2004) that many of these patients labeled as “clinically complete” are really anatomically incomplete (i.e., discomplete) (Sherwood et al., 1992), raising the potential of sub-clinical functions for residual ascending and descending spinal tracts. This notion is consistent with our demonstration of residual myelinated axons at the lesion epicenter (Hubscher and Johnson, 2006). Those rats all had “at-level” allodynia, but no electrophysiological evidence (based on thalamic recordings) of any functional ascending sensory pathways from “below-level” territories. An alternate explanation is that “at-level” allodynia in rats with severe contusion may result from local ischemia. Although we did not directly measure the degree of ischemia, this possibility is not likely since ischemia will occur in all rats with severe contusion injuries (anatomically discomplete), and we have shown in several studies that allodynia develops in only about 70% (Hubscher and Johnson, 1999a; 2006). Other groups have also found that not all rats develop allodynia, and likewise, not all SCI humans experience neuropathic pain.
The disconnection between the behavioral and electro-physiological data suggests that neuronal changes in MRF (i.e., Gi, GiA, and LPGi) may not be directly associated with the circuitries that are important for the expression of avoidance to gentle mechanical touch. A similar disconnection was obtained previously for neurons in the intralaminar nuclei of the thalamus, which receives input from the MRF (Peschanski and Besson, 1984; Jones and Yang, 1985; Peschanski and Ralston, 1985). As with MRF neurons, those in the intralaminar regions had a 23.3% increase in the number of neurons responding to touch “at-level” post-contusion regardless of the presence/absence of behavioral signs of allodynia (Hubscher and Johnson, 2006). Similar increases in sensitivity found in neurons residing in other thalamic nuclei (within ventral and posterior subregions) were, however, positively correlated with the presence of allodynia. Since the thalamic intralaminar region (like the MRF) has been implicated in contributing to arousal and vigilance (Kinomura et al., 1996; Bester et al., 1999), it is possible that the plasticity within these and other CNS areas are related to other aspects of sensation that we are not testing for. Alternatively, since these two nuclear regions project into limbic regions including the anterior cingulate cortex (ACC), a region associated with the motor avoidance response to a painful stimulus (Johansen et al., 2001), enhanced neuronal firing patterns may provide input into the motivational-affective dimension of pain but the ACC requires input from the sensory-discriminitive component pathways or some other input from the spared spinal axons in order to give rise to a stimulus-oriented motor response.
In terms of surviving circuitries contributing to neuropathic pain “at level” following contusion injury, behavioral evidence of allodynia was correlated to novel responses to touch/gentle pressure of “at-level” dermatomes for ventral and posterior thalamic neurons which normally receive widespread nociceptive-specific somatovisceral convergent inputs (Hubscher and Johnson, 2006) and are integral in pain perception pathways. The partial loss of many below-level inputs to individual groups of neurons that normally receive widespread (often whole body) inputs involved in nociceptive processing may produce an imbalance that precipitates a cascade of cellular events leading to altered responsiveness to spared inputs (i.e., dorsal trunk) and subsequent to that the eventual behavioral output (avoidance to touch). In a recent study (Hubscher et al., 2008), the degree of allodynia was found to be significantly correlated to the amount of ventrolateral funiculus damage/sparing at or immediately above the lesion epicenter. Thus, some of the surviving axons traversing the anatomically incomplete contusion site as shown in our previous study (Hubscher and Johnson, 2006) may be some remaining spinothalamic projection neurons that contribute to altered thalamic neuronal responsiveness. In addition, since incoming segmental inputs ascend and descend several segments before terminating in the spinal dorsal horn, partial loss of these inputs following SCI at any given level may likewise be responsible for some of the cellular changes noted for spared spinothalamic dorsal horn neurons “at level” (Mills and Hulsebosch, 2002; Crown et al., 2005), which likely also contributes to the altered neuronal responsiveness within the thalamus. Note, however, that like the MRF, the hyper-excitability of “at-level” dorsal horn neurons may also not be directly associated with the circuitries that are important for the expression of avoidance to gentle mechanical touch.
In a study using a combined contusion/transection lesion (Hoheisel et al., 2003), allodynia was found in 13 of 16 female rats. The differences between our results may relate to the procedures used to define the presence of allodynia. The response criteria demonstrating the presence of allodynia in the Hoheisel et al. (2003) study not only included defense reactions but also withdrawal movements or vocalization, two behaviors that are not adequate for assessing cortically mediated pain perception, as they can, for example, be evoked in decerebrate rats (Woolf, 1984). Other labs also use reflex withdrawals as all or part of their response criteria for at-level and below-level allodynia (Eaton et al., 1997; Hutchinson et al., 2004; Oatway et al., 2004; Zhang et al., 2005; Hains and Waxman, 2006). Simple reflexes do not distinguish motor versus sensory effects. Moreover, unconscious nociceptive reactions and consciously perceived pain, however, may be confounded (Vierck et al., 1989, 2005), so care must be taken when a SCI-induced change is related to specific (or a group of) behaviors.
Footnotes
Acknowledgments
We wish to thank James Armstrong for excellent technical assistance. This study was supported by NS40919 from the NIH and grant number RR015576 from the National Center for Research Resources (NCRR), a component of the NIH.
Author Disclosure Statement
No competing financial interests exist.