
Abstract
This study sought to investigate the effects of injury-induced neuropeptide Y (NPY) on c-Fos expression in the cuneate neurons and neuropathic pain after median nerve injury. Four weeks after median nerve transection (MNT), the injured nerves stimulated at low intensity (0.1 mA) expressed significantly less NPY-like immunoreactive (NPY-LI) fibers in the cuneate nucleus (CN) than those stimulated at high intensities (1.0 mA and 10 mA). Conversely, a significantly higher number of c-Fos-LI cells were observed in the CN in rats stimulated with 0.1 mA compared to those stimulated with 1.0 mA or 10 mA. These results suggest that more NPY was released following low-intensity stimulation, and consequently fewer NPY-LI fibers and more c-Fos-LI cells were identified in the CN. Furthermore, the number of c-Fos-LI cells as well as the percentage of c-Fos-LI cuneothalamic projection neurons (CTNs) in the CN was markedly decreased after injection of NPY receptor antagonist along with retrograde tract-tracing method, indicating that NPY regulated c-Fos expression. In rats with median nerve chronic constriction injury (CCI), intracerebroventricular injection of NPY aggravated mechanical allodynia and low-intensity stimulus-evoked c-Fos expression, both of which were reversed by injection of NPY receptor antagonist. However, thermal hyperalgesia was not affected by injection of these two reagents. Taken together, these findings suggest that more NPY release, following low-intensity electrical stimulation of the injured nerve, significantly induces c-Fos expression in the CTNs, which possibly provide the ascending thalamic transmission of neuropathic pain signals.
Introduction
T
Based on the above observations, we hypothesized that electrical stimulation of the injured median nerve induced the release of NPY, which subsequently modulated c-Fos expression in the cuneate neurons and neuropathic pain development. To test this hypothesis, we examined the changes of NPY-LI fibers and c-Fos-LI cells in the injured side CN following various electrical stimulation intensities applied to the transected median nerve at 4 weeks after the operation. Meanwhile, we employed the stereotaxic injection of NPY receptor antagonist to clarify the role of NPY in the modulation of c-Fos expression in the CN. If the number of c-Fos-LI cells was adjusted in the CN by NPY receptor antagonist application, we also sought to determine whether they would be derived from CTNs. A neuropathic pain model, the chronic constriction injury (CCI) model, produced signs of mechanical allodynia and thermal hyperalgesia (Bennett and Xie, 1988; Obata et al., 2003). Therefore, to further address the role of NPY in neuropathic pain, we evaluated the effects of intracerebroventricular injection of NPY receptor antagonist or NPY on median nerve CCI-induced behavioral hypersensitivity and c-Fos expression.
Materials and Methods
Animal preparations
These experiments were reviewed and approved by the National Science Council Committee, as well as the Animal Center Committee, College of Medicine, National Taiwan University, Taiwan. Ethical guidelines from the Internal Association for the Study of Pain (Zimmermann, 1983) were followed in the use of animals. All efforts were made to minimize suffering and the number of experimental animals used was kept to a minimum. All studies used male Sprague-Dawley rats, weighing 180–250 g, that were housed under approved conditions with a 12/12 h light/dark cycle with food and water available ad libitum.
Surgery and electrical stimulation
Surgery
Rats were deeply anesthetized with 7% chloral hydrate (0.45 mL/100 g body weight, i.p.), and the median nerve was separated from the surrounding tissue at the level of the elbow immediately proximal to the entry between the two heads of the pronator teres muscle. A tight ligature (5.0 silk) was made around the nerve, and an approximately 2-mm segment of the distal end was removed (Lue et al., 2002; Tsai et al., 2004, 2007). The wound was subsequently sutured. For the sham surgeries, the median nerve was exposed at the same area on the forelimb, but not transected.
Electrical stimulation
Bi-polar silver hook electrodes were placed under the isolated median nerve, immediately proximal to the level of the elbow joint, at least 5 mm proximal to the nerve injury (transection) level, or at the same level in the sham-operated rats. Paraffin oil was applied around the exposed segment of the nerve, and a 10-min pulse train of electrical stimulation from a stimulator (Grass Technologies, West Warwick, RI) was performed through a constant-current unit (Grass CCU1A). All electrical stimuli applied to the nerves were administered with a pulse duration of 0.1 ms at 10 Hz. For sham stimulation, the median nerve was connected to stimulating electrodes, but no electrical stimulation was applied.
Experimental groups
The animals were divided into four groups (Fig. 1) as follows. Group 1 animals were the control. The rats underwent stimulation of the intact median nerve at low intensity (0.1 mA [n = 3]) and high intensities (1.0 mA [n = 3] and 10 mA [n = 3]) following sham operation.

Flow diagram of the MNT groups. MNT, median nerve transection; NPY, neuropeptide Y; CN, cuneate nucleus; FG, Fluorogold.
Group 2 animals underwent various electrical stimulation intensities that were applied to the transected median nerve; changes in the NPY-LI fibers and c-Fos-LI cells in the CN were subsequently observed. Four weeks after bilateral MNT, animals underwent unilateral stimulation of the injured median nerve at 0.1 mA (n = 7), 1.0 mA (n = 7), 10 mA (n = 7), or sham stimulation (n = 7).
Group 3 animals underwent unilateral MNT, followed by stereotaxic injections of NPY receptor antagonist (NPY(18–36), American Peptide Co., Sunnyvale, CA) (Ossipov et al., 2002) or vehicle (distilled water) into the rostral region of CN ipsilateral to the injured nerve. The role of NPY in the modulation of c-Fos expression in the CN was subsequently measured. The occipital bone over the caudal medulla was removed. Using a glass micropipette with a fine tip diameter connected to a Hamilton microsyringe, 1 μL of NPY receptor antagonist (0.5 nmol) or vehicle was injected into the rostral region of CN and allowed to diffuse into the middle region of CN. The injection site was defined using the stereotaxic coordinates provided by the rat brain atlas of Pellegrino and colleagues (1979) (coordinates AP, ML, H: −6.4, 1.4, −6.4; System A). At 4 weeks after MNT, injection of NPY receptor antagonist (n = 5) or vehicle (n = 3) was performed 30 min prior to low-intensity electrical stimulation (0.1 mA) of the nerve.
Group 4 animals underwent unilateral MNT, followed by a stereotaxic injection of 1 μL of 2% Fluorogold (FG) (Fluorochrome, Denver, CO) into the contralateral ventrobasal nucleus of the thalamus (coordinates AP, ML, H: 4, 3, −0.5; 4, 3, 0; 4, 2.5, −1; System A) (Pellegrino et al., 1979; Day et al., 2001; Lue et al., 2002). The effects of NPY receptor antagonist on c-Fos expression in the CTNs were observed. FG was administered 3 days prior to the stereotaxic injection of NPY receptor antagonist (n = 5) or vehicle (n = 3), along with low-intensity electrical stimulation (0.1 mA) of the nerve at 4 weeks after MNT.
c-Fos and NPY immunocytochemistry
The rats were reanesthetized and perfused 2 h after electrical stimulation (or nerve exposure without stimulation) with 4% paraformaldehyde/0.1 M phosphate buffer (PB) (pH 7.4). Medulla tissue blocks that contained the CN were resected and stored in PB/30% sucrose overnight, and 30-μm slices were transversely cut with a cryostat (Bright, Huntingdon, United Kingdom).
The floating medulla sections were collected and treated with 0.5% H2O2, blocked with 5% normal goat serum (NGS, Gibco BRL, Grand Island, NY) in PB for 1 h, and then alternatively incubated in rabbit polyclonal anti-NPY (1:2000; DiaSorin, Stillwater, MN) (Tsai et al., 2004, 2007) or c-Fos (1:4000; Calbiochem, San Diego, CA) (Day et al., 2001; Lue et al., 2002) antibodies for 48 h at 4°C. The primary antibodies were diluted in 0.01 M phosphate buffer saline (PBS, pH 7.4), containing 0.2% Triton X-100 and 5% NGS. After rinsing with PBS, the sections were incubated in 1:200 biotinylated anti-rabbit IgG (Vector Laboratories, Burlingame, CA) for 2 h, and processed with ABC (avidin-biotin-HRP complex, ABC kit, Vector) for 1 h at room temperature. Peroxidase activity was subsequently visualized by treating with Vector SG Substrate kit. Finally, the floating sections were mounted onto gelatinized slides and examined under a light microscope (Axiophot, Zeiss, Jena, Germany). Immunolabeling was absent in all sections incubated in normal rabbit serum in lieu of the above-mentioned primary antibodies, or with the omission of the same antibodies as control.
Enzyme-linked immunosorbent assay
Four weeks after bilateral MNT, animals underwent unilateral stimulation of the injured median nerve with 0.1 mA (n = 5), 1.0 mA (n = 5), 10 mA (n = 5), and sham stimulation (n = 5). Two hours after electrical stimulation (or nerve exposure without stimulation), all rats were decapitated. Following removal of the CN, they were frozen in liquid nitrogen and homogenized in ice-cold PBS (pH 7.4), containing protease inhibitors (aprotinin, leupeptin, pepstatin, PMSF) (Sigma, St. Louis, MO). After centrifugation at 10,000 g for 15 min at 4°C, supernatant was collected and NPY levels were quantified with a rat NPY ELISA kit (Phoenix Pharmaceuticals, Belmont, Canada). A monoclonal NPY-specific rat antibody was pre-coated onto a microplate. The supernatant and standards (Phoenix Pharmaceuticals) were pipetted into the wells, and any rat NPY was bound by the immobilized antibody. After washing away unbound substances, an enzyme-linked polyclonal antibody specific for rat NPY was added to the wells. Following a wash to remove any unbound antibody-enzyme reagent, a substrate solution was added to the wells. The enzyme reaction yielded a blue product that turned yellow after exposure to the “stop” solution. The intensity of the colored product was directly proportional to the concentration of rat NPY present in the original specimen. The microplates were read at 450 nm by a microplate reader (Molecular Devices, Sunnyvale, CA). The NPY concentration was obtained from a standard curve. NPY levels in the ipsilateral CN (MNT with electrical stimulation) and contralateral CN (MNT without electrical stimulation) from groups that received electrical stimulation at various intensities were statistically analyzed with one-way ANOVA or Student's t-test (Fig. 3B). The significance level was set to p < 0.05.
Chronic constriction injury studies
Drug delivery
Experimental rats were anesthetized (7% chloral hydrate, 0.45 mL/100 g body weight i.p.) and placed into a stereotaxic head holder to prepare for drug injection into the cerebral ventricle. The skull was exposed, and a brain infusion cannula was directed towards the cerebral ventricle. The injection site was determined according to stereotaxic coordinates provided by the rat brain atlas of Pellegrino and colleagues (1979). The brain infusion cannula was cemented in place and secured to the skull with instant adhesive gel. The animals were allowed to recover for 3 days after surgery. All rats were reanesthetized and implanted with an Alzet osmotic pump (model 2004, this model has a reservoir volume of 200 μL and a 28-day delivery duration), which was filled with 0.5 nmol of NPY receptor antagonist (n = 8), 1 nmol of NPY (American Peptide) (Ossipov et al., 2002) (n = 8), or vehicle (n = 8). Prior to implantation, osmotic pumps were incubated in a 0.9% NaCl solution for 24 h at 37°C and filled according to the supplier instructions. The catheter tube from the Alzet osmotic pump was attached to a brain infusion cannula, which was installed in the subcutaneous pocket of the back.
After implantation, unilateral median nerve chronic constriction injury (CCI) was performed on all experimental animals. The model of CCI on the median nerve was based on the methods previously described by Bennett and Xie (1988). Specifically, under a dissecting microscope, the right median nerve was separated from the surrounding tissue at the elbow level, immediately proximal to the entry between the two heads of the pronator teres muscle. Four loose ligatures (4.0 chromic gut) were made around the nerve. Subsequently, the incision was sutured. Six rats received a sham operation (median nerve exposure without ligation) at the same area on the right forelimb, and these rats did not receive intracerebroventricular injection. Upon removal of the pump at the end of the experiment, residual volume in the reservoir was measured to confirm solution delivery.
Behavioral testing and electrical stimulation
Behavioral testing was performed on nerve-ligated and sham-operated animals. A set of Von Frey filaments (Somedic Sales, Hörby, Sweden) and the plantar test (Ugo Basile, Comerio, Italy) (Hargreaves et al., 1988) were used to test withdrawal thresholds (g) and latency (s) of the rat forepaws.
Von Frey filaments of different bending forces, including 0.145, 0.32, 0.39, 1.1, 1.7, 3.3, 5.1, 8.3, 17, and 24 g, were used to test the mechanical threshold of the rat forepaws. The differences between successive filaments, which present the log10 of the bending forces, are irregular and the series is an ordinal scale. Testing was performed in the forepaw based on previously described practices by Tal and Bennett (1994). Each filament was applied five times to the medial plantar surface of each rat forepaw. Testing began with the smallest bending force and continued in increasing order. The first filament in the series that evoked at least two withdrawal responses was defined as the threshold. Tests were performed in the morning, one day prior to nerve ligation, and at several time points after nerve ligation. The parametric statistical procedure, Student's t-test, was used to measure the differences between groups. Significance was set at p < 0.05.
To measure the thermal threshold of the forepaws, the plantar test was utilized. Briefly, the rats were placed on an elevated glass floor, where they could move about freely. A plastic barrier was used to prevent them from escaping. After an adaptation period during which they could explore the cage, the rats remained calm and quiet. A radiant heat source was positioned under the glass floor directly beneath the forepaws. The withdrawal latencies of the forepaws to the radiant heat source were automatically measured to the nearest 0.1 s by the apparatus. A maximal cut-off latency of 40 s was used to prevent tissue damage. A more complete description of the apparatus has been previously provided (Hargreaves et al., 1988). The forepaws were tested in a random fashion. Four measurements were taken for each forepaw in a session. The interval between two consecutive trials was at least 10 min. Four latency values per side were averaged. Data were summarized and expressed as mean ±standard deviation. The Student's t-test was performed to test for differences between groups. Significance was set at p < 0.05. Animals were tested in the afternoon, at least 5 h after Von Frey filament testing, one day prior to nerve ligation, and at several time points after nerve ligation. One day after the end of behavioral testing, all animals underwent stimulation of the injured median nerve at 0.1 mA (n = 5 in each reagent-treated group), or sham stimulation (n = 3 in each reagent-treated group). In sham-operated group, animals underwent stimulation of the intact median nerve at 0.1 mA (n = 3) or sham stimulation (n = 3). The rats were reanesthetized and perfused 2 h after electrical stimulation (or nerve exposure without stimulation) with 4% paraformaldehyde/0.1 M PB (pH 7.4). Medulla tissue blocks that contained the CN were resected and stored in PB/30% sucrose overnight, and 30-μm slices were transversely cut with a cryostat (Bright). Subsequently, the floating medulla sections were processed for the c-Fos and NPY immunocytochemistry as described above.
Data presentation and statistical analysis
For quantitative analysis, tissue sections were collected from the entire rostrocaudal extent of the CN. The middle CN was defined as the area 0.3–0.7 mm caudal to the obex (Maslany et al., 1991; Day et al., 2001; Lue et al., 2002; Tsai et al., 2004). Seven sections were collected from the middle region of each group. To determine the changes in NPY and c-Fos immunoreactivity in the CN, sections were examined with a Zeiss light microscope and images were captured with a digital camera (Xillix MicroImager 1400, Nikon, Tokyo, Japan) at a magnification of 100x. Pictures were processed and evaluated with a computer-based image analysis system (MGDS) and Image Pro-Plus software (Media Cybernetics, Bethesda, MD). The area occupied by NPY-LI fibers and the area of outlined CN were measured (Tsai et al., 2004). The former divided by the latter was defined as the percentage of area occupied by NPY-LI fibers in the ipsilateral CN (Figs. 3A and 12A). The density of c-Fos-LI cells in the CN was defined as the number of c-Fos-LI cells divided by the number of tissue sections (Figs. 5, 7, and 12B). The mean percentage of area occupied by NPY-LI fibers and the density of c-Fos-LI cells in each experimental group were calculated and statistically compared with one-way ANOVA or Student's t-test (Figs. 3A, 5, 7, and 12). In experimental group 4, the c-Fos-LI cells, FG-labeled CTNs, and c-Fos-LI cells containing FG (c-Fos-LI/FG double-labeled neurons) were enumerated using a combination of bright field and incident FG fluorescence illumination with a wide-band ultraviolet filter. The densities of c-Fos-LI cells, FG-labeled CTNs, and double-labeled neurons in the middle CN were defined by the number of each cell type divided by the total number of respective tissue sections (Fig. 9). The percentage of double-labeled neurons was calculated as the number of double-labeled neurons divided by the number of total c-Fos-LI cells (Fig. 9). The densities of c-Fos-LI cells, FG-labeled CTNs, double-labeled neurons, and the percentage of double-labeled neurons among different experimental groups were statistically compared with Student's t-test (Fig. 9).
Results
After nerve injury, all experimental rats appeared to be physically healthy, maintained normal body weight, and displayed normal activities. Although the rats often raised the affected forelimbs and held them in guarded positions while standing, there was no evidence of self-mutilation to the forelimbs.
Effects of varied electrical intensities on NPY and c-Fos expression in the cuneate nucleus
Few, if any, NPY-LI fibers or c-Fos-LI cells were detected in the CN of sham-operated rats, with or without electrical stimulation (Figs. 2A and 3A). However, numerous NPY-LI fibers were detected after sham stimulation at 4 weeks after bilateral MNT, particularly in the ipsilateral middle CN (Fig. 2B). Following unilateral electrical stimulation (0.1 mA) to the injured nerve, a few NPY-LI fibers were observed in the ipsilateral CN (Fig. 2C). However, there were large amounts of NPY-LI fibers in the CN of animals stimulated with 1.0 mA or 10 mA after nerve injury (Fig. 2D and E). Quantitative analysis of NPY expression using morphometric measurement and ELISA demonstrated that NPY levels in the ipsilateral CN were significantly less in animals stimulated with 0.1 mA compared to animals stimulated with 1.0 mA and 10 mA (Fig. 4). Conversely, animals unilaterally stimulated with 0.1 mA, after bilateral MNT (Fig. 3B), presented with more c-Fos-LI cells in the ipsilateral middle CN compared to that of animals stimulated with 1.0 mA and 10 mA (Fig. 3C and D). Quantitative analysis also demonstrated that the average number of c-Fos-LI cells in the ipsilateral CN was significantly higher in animals stimulated with 0.1 mA compared to those stimulated with 1.0 mA and 10 mA (Fig. 5).
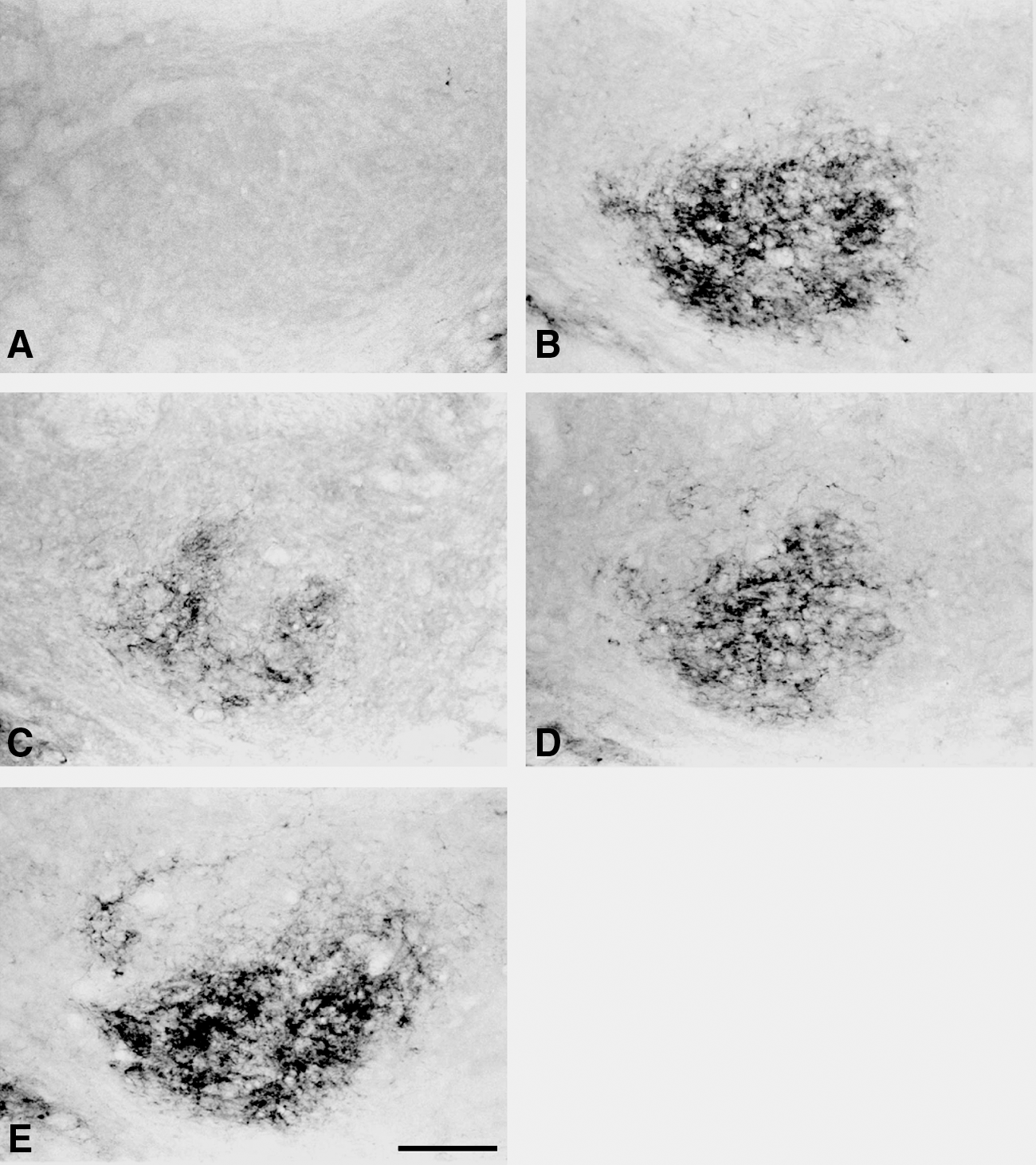
Photomicrographs displaying NPY-LI fibers in the middle region of the CN, ipsilateral to the sham-operated median nerve (

Photomicrographs showing c-Fos-LI cells in the middle region of the CN, ipsilateral to the injured nerve 4 weeks after bilateral MNT with unilateral sham stimulation (
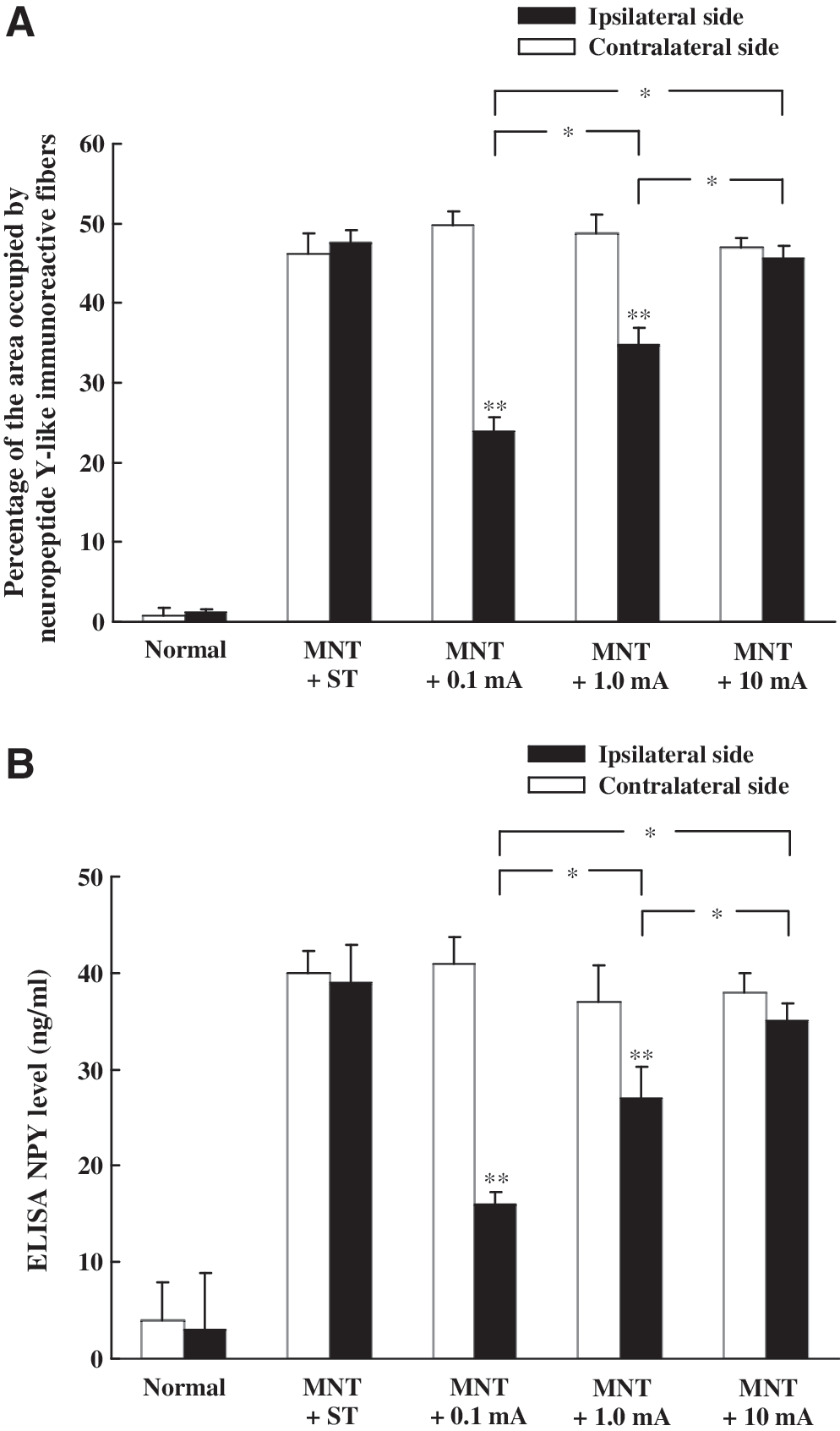
Histograms showing morphometric measurements (

Histograms showing c-Fos-LI cells in the middle region of the CN following unilateral electrical stimulation with various intensities applied to the injured nerve 4 weeks after bilateral MNT. Note that the animals stimulated with 0.1 mA exhibited significantly more c-Fos-LI cells in the CN compared to that of the animals stimulated with 1.0 mA and 10 mA (*p < 0.05). ST, sham stimulation.
Effects of pretreatment with an NPY receptor antagonist in addition to low-intensity electrical stimulation
Using a stereotaxic injection of an NPY receptor antagonist in combination with low-intensity electrical stimulation (0.1 mA) to the injured nerve, the number of NPY-LI fibers and c-Fos-LI cells in the ipsilateral middle CN were dramatically reduced (Fig. 6). Statistical analysis demonstrated that the number of c-Fos-LI cells in the NPY receptor antagonist group was significantly less than in the vehicle group (Fig. 7). Group 4 rats subsequently received a FG injection to the thalamus; FG-labeled CTNs and immunoperoxidase-labeled c-Fos-LI cells were detected under fluorescence and bright field illumination throughout the CN, respectively. In the NPY receptor antagonist and vehicle groups, double-labeling demonstrated the presence of abundant FG-labeled CTNs intermingled with immunoperoxidase-labeled c-Fos-LI cells as well as some FG-labeled c-Fos-LI (double-labeled) neurons (Fig. 8). The density of c-Fos-LI cells, FG-labeled CTNs, and double-labeled neurons, as well as the percentage of double-labeled neurons in the middle region of CN, are presented in Figure 9. After comparing the NPY receptor antagonist and vehicle groups, there was no significant difference in the density of FG-labeled CTNs in the CN. However, the density of c-Fos-LI cells and double-labeled neurons, as well as the proportion of double-labeled neurons among c-Fos-LI cells, was markedly less in the NPY receptor antagonist group compared to that of the vehicle group (Fig. 9).
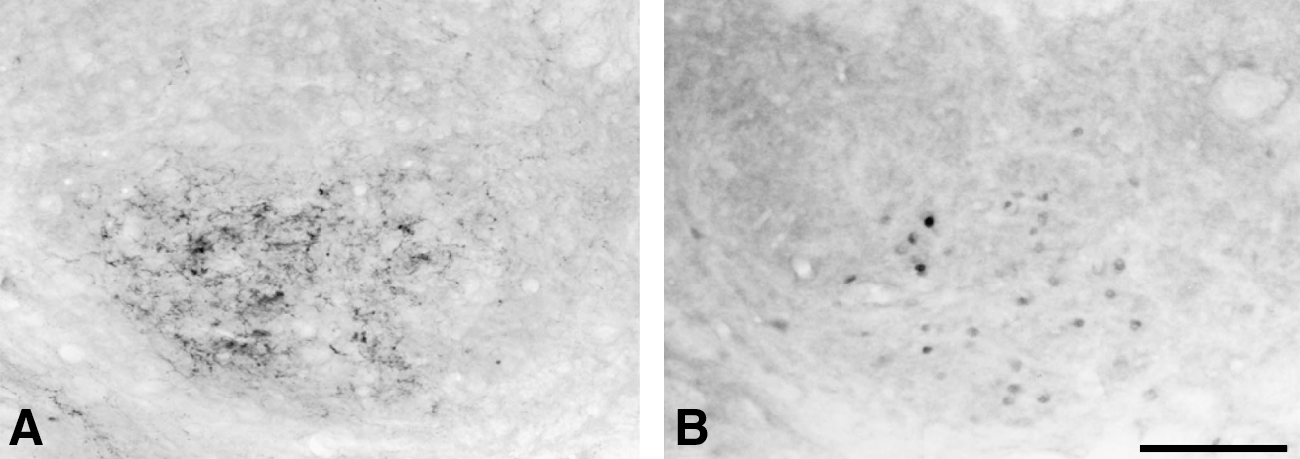
Photomicrographs showing NPY-LI fibers (

Histograms showing c-Fos-LI cells in the middle region of the CN after stereotaxic injection of NPY receptor antagonist or vehicle into the rostral region of CN, followed by low-intensity electrical stimulation to the injured nerve 4 weeks after MNT. Note that the number of c-Fos-LI cells in the NPY receptor antagonist group is significantly less than that in the vehicle group (*p < 0.05).

Photomicrographs showing FG-labeled CTNs (asterisks) and c-Fos-LI cells (arrows) in the middle region of the CN after stereotaxic injection of NPY receptor antagonist (

Histograms showing the density of c-Fos-LI cells, FG-labeled CTNs, double-labeled neurons (
Amelioration of CCI-induced mechanical allodynia upon application of NPY receptor antagonist
Von Frey filament testing showed that the intracerebroventricular injection of vehicle in CCI rats decreased mechanical withdrawal thresholds from a basal value of 15.9 ± 2.3 g to 4.8 ± 1.0 g; rats developed mechanical allodynia one day after CCI, which persisted for 28 days (Fig. 10A). However, intracerebroventricular injection of NPY receptor antagonist to CCI rats increased mechanical withdrawal thresholds and ameliorated the tactile hypersensitivity (Fig. 10A). On the other hand, intracerebroventricular injection of NPY aggravated CCI-induced tactile hypersensitivity (Fig. 10A). CCI rats received the intracerebroventricular injection of vehicle and plantar testing was performed; thermal withdrawal latency decreased from a basal value of 6.4 ± 0.2 s to 3.7 ± 0.3 s, and the rats developed thermal hyperalgesia (Fig. 10B). However, thermal withdrawal latencies of CCI rats that received either NPY receptor antagonist or NPY were not significantly different compared to CCI rats that received vehicle treatment (Fig. 10B).
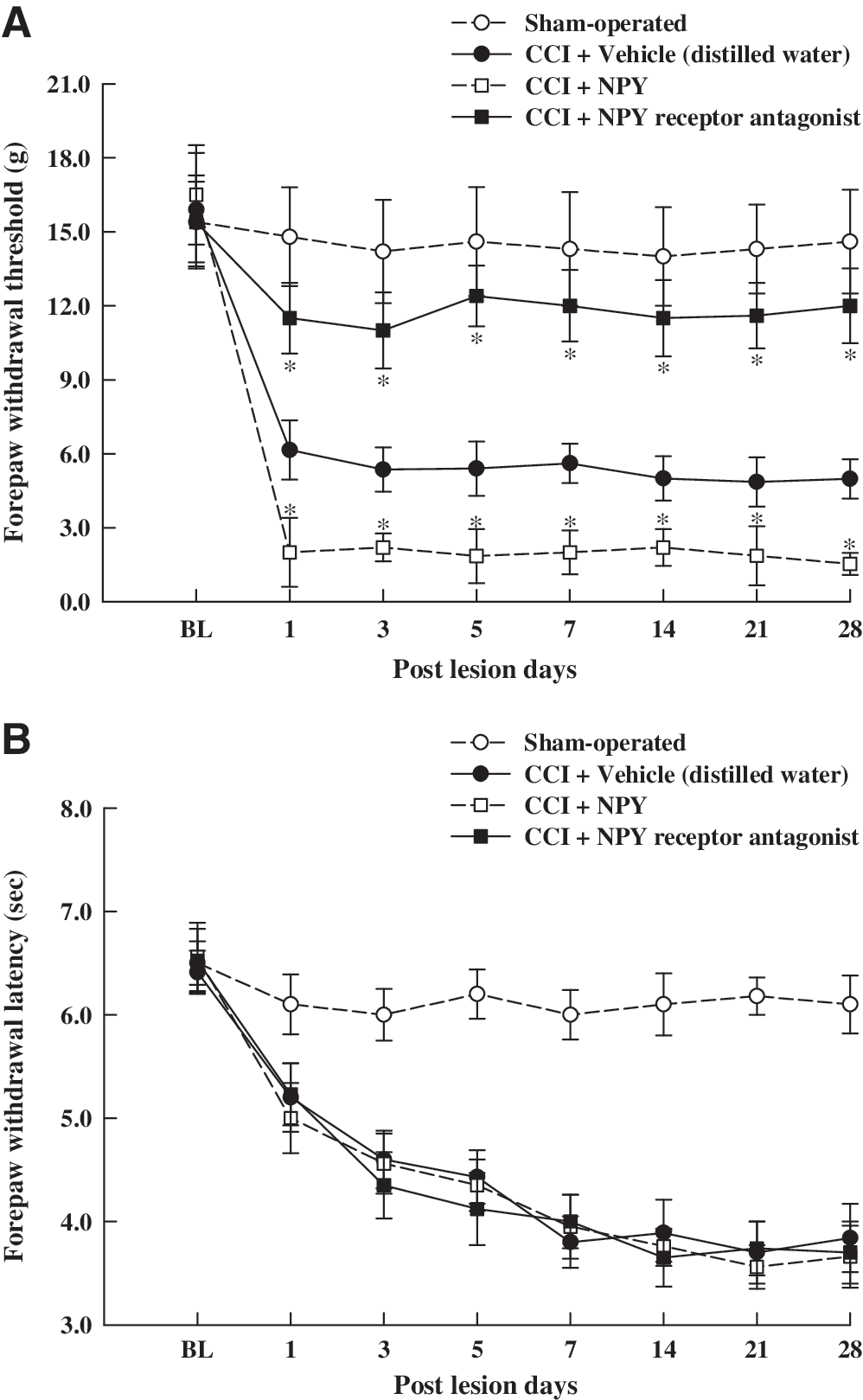
Effects of intracerebroventricular injection of the NPY receptor antagonist or NPY on tactile hypersensitivity (
Reduced c-Fos expression after application of NPY receptor antagonist in CCI rats with low-intensity electrical stimulation
After intracerebroventricular injection of vehicle in CCI rats with sham stimulation, numerous NPY-LI fibers were detected in the ipsilateral middle CN, but almost no c-Fos-LI cells were observed (Fig. 11A and B). However, upon intracerebroventricular injection of vehicle (Fig. 11C and D) or NPY (Fig. 11E and F) to CCI rats following low-intensity electrical stimulation (0.1 mA) of the injured nerve, fewer NPY-LI fibers and more c-Fos-LI cells were identified in the CN. In addition, upon intracerebroventricular injection of NPY receptor antagonist in combination with low-intensity electrical stimulation to the injured nerve, the number of NPY-LI fibers and c-Fos-LI cells in the CN were dramatically reduced (Fig. 11G and H). Quantitative analysis demonstrated that the vehicle-treated CCI rats with low-intensity electrical stimulation showed a significant decrease in the number of NPY-LI fibers in the CN compared to that of the vehicle-treated CCI rats with sham stimulation (Fig. 12A). However, following low-intensity electrical stimulation, there was no significant difference in the number of NPY-LI fibers in the CN between the vehicle-, NPY-, and NPY receptor antagonist-treated CCI groups (Fig. 12A). Quantitative analysis of c-Fos expression showed that the average number of c-Fos-LI cells in the ipsilateral CN was significantly greater in the vehicle-treated CCI rats with low-intensity electrical stimulation compared to that of the vehicle-treated CCI rats with sham stimulation (Fig. 12B). In NPY-treated CCI rats, the number of low-intensity electrical stimulation-induced c-Fos-LI cells was significantly greater than that in the vehicle-treated CCI rats (Fig. 12B). Conversely, in NPY receptor antagonist-treated CCI rats, the number of low-intensity electrical stimulation-induced c-Fos-LI cells was significantly less than that in the vehicle-treated CCI rats (Fig. 12B).
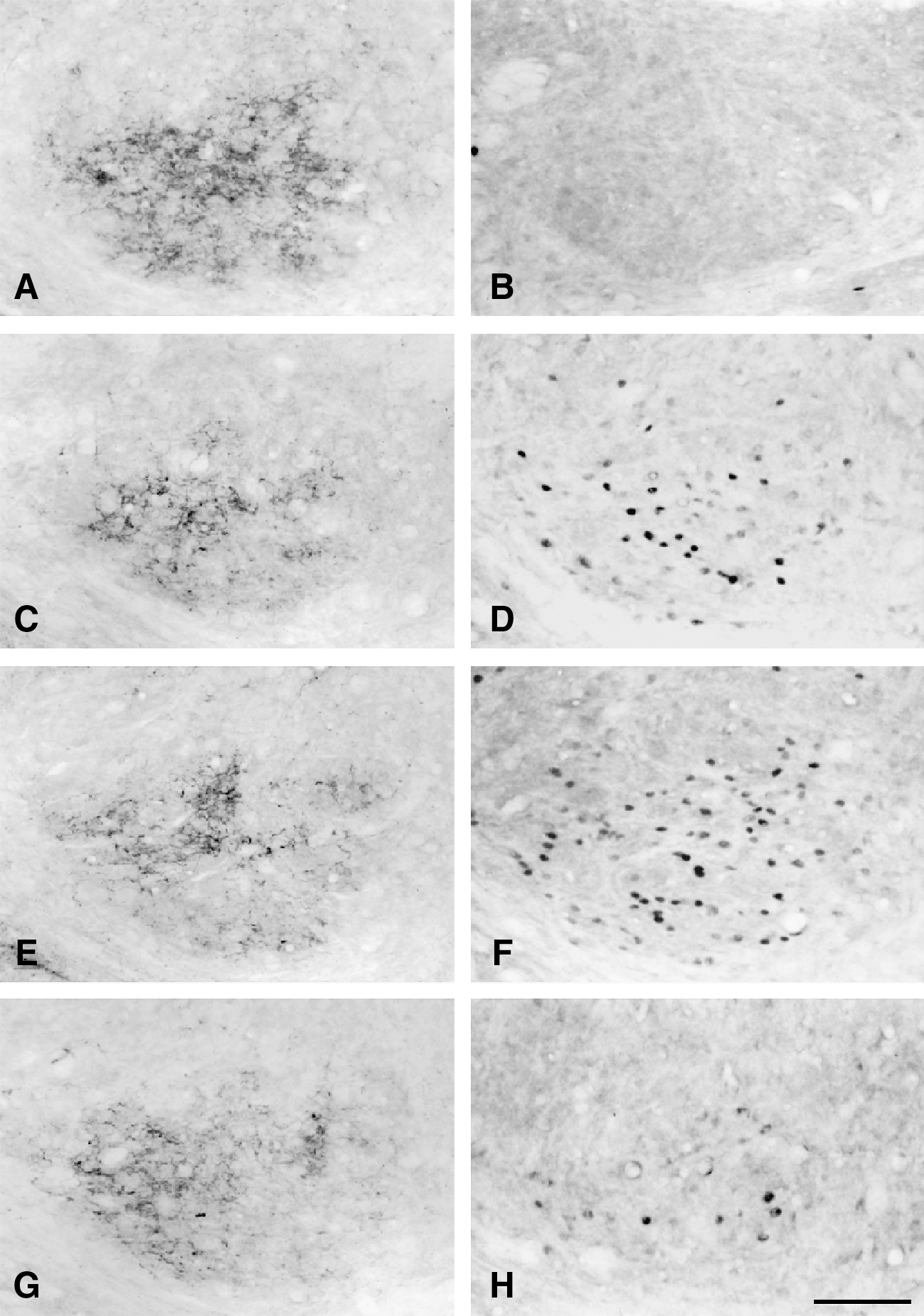
Photomicrographs showing NPY-LI fibers (left panel) and c-Fos-LI cells (right panel) in the middle region of the CN in CCI rats after injection of vehicle with sham (
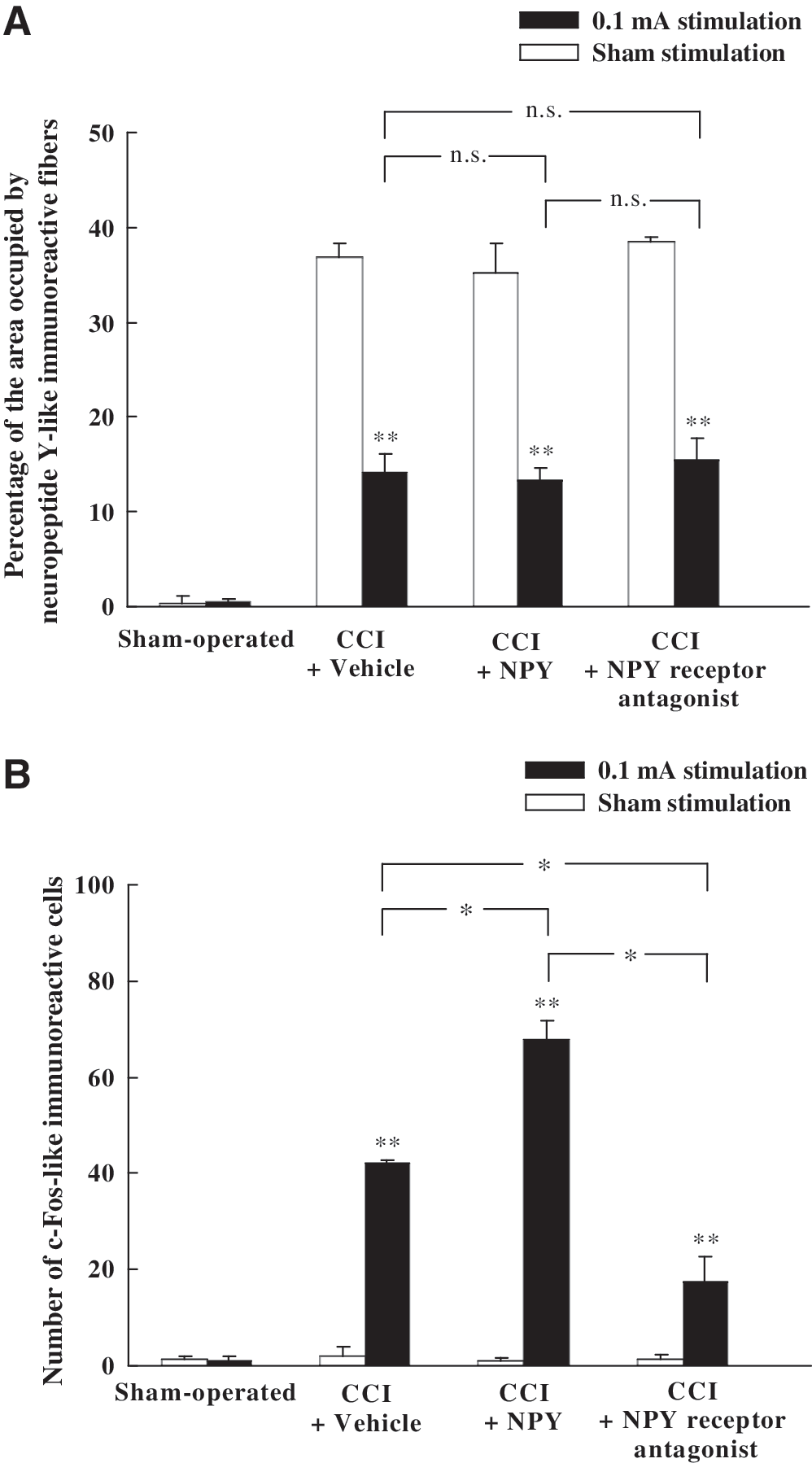
Histograms showing NPY-LI fibers (
Discussion
This study demonstrates that low-intensity electrical stimulation to the injured median nerve results in few NPY-LI fibers, but many c-Fos-LI cells in the CN. Upon NPY receptor antagonist injection into the rostral CN, the number of c-Fos-LI cells and percentage of c-Fos-LI CTNs in the CN were remarkably reduced. In addition, intracerebroventricular administration of NPY receptor antagonist significantly ameliorated CCI-induced mechanical allodynia, but aggravated allodynia was observed after intracerebroventricular application of NPY. In the CN of CCI rats with electrical stimulation of the injured nerve, intracerebroventricular injection of NPY and NPY receptor antagonist promoted and inhibited c-Fos expression, respectively. The results suggest that injury-induced NPY expression modulates c-Fos expression and neuropathic pain development.
After application of electrical stimulation of varied intensities to the injured nerve at 4 weeks after MNT, the number of NPY-LI fibers in the ipsilateral CN was significantly decreased, but the number of c-Fos-LI cells was significantly increased in rats stimulated at low-intensity (0.1 mA) compared to rats that received high-intensity (1.0 mA and 10 mA) or sham stimulations. The results suggested that more NPY was released following low-intensity stimulation, and consequently fewer NPY-LI fibers and more c-Fos-LI cells were identified in the CN. A recent study demonstrated that NPY-LI fibers in the CN are exclusively derived from injured dorsal root ganglion (DRG) neurons, particularly medium- and large-diameter neurons, via primary afferent fibers (Tsai et al., 2007). In the present study, low-intensity electrical stimulation to the injured nerve, which could only excite large myelinated Aα/β fibers (Woolf and Wall, 1982; Nishimori et al., 1990), resulted in few NPY-LI fibers but numerous c-Fos-LI cells in the CN. We speculate that NPY release from primary afferent terminals was promoted by low-intensity electrical stimulation and, subsequently, the peptide influenced the excitability of CN neurons, as well as c-Fos expression. The mechanisms of reduced NPY release after nerve injury upon high stimulation intensities remain uncertain; however, several possibilities are discussed below. Previous studies have shown that primary afferent terminals contain the excitatory glutamate neurotransmitter and make axodendritic synaptic contacts with GABA-LI dendrites in the CN (Lue et al., 1996, 1997). Pharmacological and morphological studies have demonstrated that GABA plays an important role in presynaptic inhibition of primary afferent terminals (Davidson and Southwick, 1971; Lue et al., 1996). Furthermore, high-intensity stimulation to the dorsal root produced a significant increase in glutamate release in spinal cord slices (Kawagoe et al., 1986; Kangrga and Randic, 1990). Electrophysiological studies (Chung et al., 1984a, 1984b) also revealed that high-intensity conditioning stimulation to the sural nerve caused a powerful inhibition of spinothalamic tract neurons and was associated with long-lasting analgesia. Taken together, these findings suggest that high-intensity electrical stimulation to the injured median nerve might enhance glutamate release and elicit GABA-LI neuronal activity. These in turn could exert an inhibitory influence on primary afferent terminals to lessen NPY release.
Yang et al. (1995) reported that NPY application in the solitary tract nucleus induced an increase of c-Fos-LI cells in the same nucleus. Moreover, the time course of changes in the number of NPY-LI fibers and c-Fos-LI cells were similar in the ipsilateral CN following median nerve injury (Lue et al., 2002; Tsai et al., 2004). After application of NPY receptor antagonist to the CN, along with low-intensity electrical stimulation to the injured nerve, the number of NPY-LI fibers and c-Fos-LI cells were remarkably decreased in the ipsilateral CN. These results suggested that NPY release was a result of low-intensity electrical stimulation to the injured nerve; moreover, NPY receptor antagonist blocked the induction of c-Fos expression. Consequently, less NPY-LI fibers and c-Fos-LI cells were identified in the CN. Prior studies revealed that after electrical stimulation to the injured median nerve, a greater proportion (about 78%) of c-Fos-LI cells in the middle region of the CN were CTNs (Day et al., 2001; Lue et al., 2002), and the injury-induced NPY-LI terminals made direct synaptic contacts with the CTN dendrites (Yeh et al., 2008). The present study demonstrated that administration of NPY receptor antagonist decreased the number and percentage of c-Fos-LI CTNs in the CN. In light of the above findings, it is probable that injury-induced NPY expression affected CTN excitability and modulated the c-Fos protein expression.
Although injury-induced NPY might directly mediate c-Fos expression in the CN after electrical stimulation to the injured median nerve, the role of this neuropeptide in the modulation of nociception has remained inconclusive. Ohara and colleagues (1994) reported that following sciatic axotomy the excitability of sensory neurons might be mediated by NPY upregulation, which increased the excitability of DRG neurons through the enrichment of Y2-mediated excitation and repression of Y1-mediated inhibition (Abdulla and Smith, 1999). Following peripheral nerve injury, subcutaneous injection (Tracey et al., 1995) or intrathecal administration (White, 1997) of NPY might aggravate injury-induced mechanical hyperalgesia. Furthermore, microinjection of an NPY receptor antagonist or anti-NPY antiserum into the gracile nucleus ipsilateral to the nerve injury side caused a reversal of tactile, but not thermal, hypersensitivity in the affected hindpaw (Ossipov et al., 2002). In this study intracerebroventricular administration of NPY receptor antagonist significantly ameliorated CCI-induced mechanical allodynia and decreased the number of low-intensity electrical stimulation-induced c-Fos-LI cells in the CN, whereas application of NPY brought about the opposite results. The expression of c-Fos immunoreactivity has been accepted as a useful marker for elucidating nociceptive pathways (Hunt et al., 1987; Harris, 1998), and the antinociceptive effects of many treatments have been assessed by examining their ability to reduce the expression of c-Fos (Zhang et al., 1994; Mitsikostas et al., 1998). This study demonstrated that the number of c-Fos-LI cells was positively correlated with the degree of mechanical allodynia. Taken together, the results suggested that injury-induced NPY might upregulate the c-Fos expression in the CN, which was associated with the magnitude of tactile hypersensitivity following median nerve injury.
In summary, the present study demonstrated that injury-induced NPY expression might directly mediate c-Fos expression in CTNs. In addition, the number and percentage of CTNs expressing c-Fos (double-labeled neurons) in the NPY receptor antagonist group were significantly less than that in the vehicle group. Moreover, CCI-induced mechanical allodynia was ameliorated and exacerbated by intracerebroventricular injection of NPY receptor antagonist and NPY, respectively. Thus, following low-intensity electrical stimulation to the injured nerve, it is possible that NPY released from the injured median primary afferent terminals upregulated c-Fos expression in the CTNs. This pathway might be involved in the transmission of tactile hypersensitivity to the thalamus.
Footnotes
Acknowledgments
We are grateful to Ms. Yi-Tien Li for technical assistance. This study was supported in part by a research grant from the National Science Council of Taiwan (NSC 94-2320-B002-003) and a grant from the Fu Jen Catholic University (109531030990-2), Taiwan.
Author Disclosure Statement
No conflicting financial interests exist.