
Abstract
Proliferation of Schwann cells in the injured peripheral nerve supports axonal regeneration, and physical training in experimental animals has been shown to promote nerve regeneration. Extracellular signal-regulated kinase 1/2 (ERK1/2) activity can mediate neuronal responses to lesion signals, but its role in non-neuronal cells in the injured area is largely unknown. Here we report that treadmill training (TMT) facilitates axonal regeneration via the upregulation of phospho-ERK1/2 protein levels in Schwann cells in the injured sciatic nerve. Low-intensity, but not high-intensity, TMT increased neurite outgrowth of dorsal root ganglion (DRG) sensory neurons and potentiated Schwann cell proliferation. TMT elevated levels of GAP-43 mRNA and protein, and phospho-ERK1/2 protein in the injured sciatic nerves. TMT also enhanced phospho-c-Jun protein levels in the injured nerve. In-vivo administration of the ERK1/2 inhibitor PD98059 eliminated phospho-c-Jun, suggesting ERK1/2 phosphorylation of the c-Jun protein. PD98059 treatment decreased levels of BrdU-labeled proliferating Schwann cells in the distal portion of the injured nerve, and delayed the axonal regrowth that was promoted by TMT. The present data suggest that increased ERK1/2 activity in Schwann cells may play an important role in TMT-mediated enhancement of axonal regeneration in the injured peripheral nerve.
Introduction
I
Physical exercise such as swimming, treadmill training (TMT), and weight-bearing training can increase neural activity in both the peripheral and central nervous systems (Cotman et al., 2007). Van Meeteren and colleagues (1997) have reported that exercise training not only enhanced signal conduction velocity in motor nerves, but also sustained the beneficial effect on functional recovery until the late phase of peripheral nerve regeneration. After spinal cord contusion, physical exercise improved sensory function, which was positively correlated with brain-derived neurotrophic factor (BDNF) levels in the injured spinal cord (Hutchinson et al., 2004). We have recently reported TMT-mediated enhancement in axonal regeneration of injured sciatic nerve and Schwann cell activation (Seo et al., 2006). Schwann cell activation in the injured nerve appeared to involve increased Cdc2 kinase activity known to be important for Schwann cell proliferation and migration (Han et al., 2007). However, the mechanisms of transducing external injury stimulation and physical activity into target molecules remain to be explored.
Here we investigated extracellular signal-regulated kinase 1/2 (ERK1/2) as a potential mediator for TMT-induced enhancement in axonal regeneration. As noted above, BDNF expression is increased in several neural tissues, including brain, spinal cord, and peripheral nerves by physical training or injury (Neeper et al., 1995; Hutchinson et al., 2004; Meyer et al., 1992; Molteni et al., 2004). Moreover, there is compelling evidence that BDNF administration may improve axonal regrowth after injury in the spinal cord and peripheral nerves (Vavrek et al., 2006; Song et al., 2008; Kobayashi et al., 1997; Hontanilla et al., 2007). ERK1/2 is known to be activated by diverse types of external stimulations, including BDNF; thus ERK1/2 may respond to an animal's physical activity and facilitate axonal regrowth. ERK1/2 functions in various cellular events ranging from proliferation and differentiation to regeneration, and is one of the key mediators for transmitting distal lesion signals into the nucleus for neuronal gene expression (Segal and Greenberg, 1996; Perlson et al., 2005). Increased ERK1/2 activation was found in regenerating peripheral nerves after injury (Sheu et al., 2000), particularly in the Schwann cells in the distal sciatic nerve, where proliferating Schwann cells form the band of Büngner and participate actively in regeneration processes (Agthong et al., 2006; Tapinos and Rambukkana, 2005). We have found nerve cell potentiation by TMT in the form of enhanced neurite outgrowth of DRG sensory neurons and Schwann cell proliferation. We further demonstrate that ERK1/2 activity was induced in Schwann cells by TMT, stimulated c-Jun phosphorylation, and facilitated axonal regeneration.
Methods
Animals and general procedures
Male Sprague-Dawley rats (n = 118, 7–8 weeks old, body weight 200–220 g; Samtago, Osan, Korea) were used in this experiment. They were placed in an animal room with controlled temperature (22°C) and 60% humidity, with a 12-h/12-h light-dark cycle. The animals were fed commercial chow (Samyang Co., Seoul, Korea) and water ad libitum. Rat care and all experimental procedures were in accordance with the Animal-Use Statement and Ethics Committee Approval Statement for Animal Experiments at Daejeon University, Daejeon, Korea. All the animals were acclimatized in the animal room for 1–2 weeks before use. For the experiments comparing the effects of training intensity, the rats were randomly assigned to intact control (CTL, n = 6), low-intensity TMT (LITMT, n = 6), and high-intensity TMT (HITMT, n = 6) groups. For the experiments examining the effects of TMT on nerve regeneration, the animals were divided into a sedentary group with sciatic nerve injury without exercise (n = 30), and a TMT group having sciatic nerve injury with TMT (n = 60); the TMT group included animals treated with PD98059 (TMT + PD, n = 18), and those treated with dimethyl sulfoxide (DMSO) (TMT + vehicle, n = 18). For the retrograde tracing experiments, 10 rats were used.
Sciatic nerve surgery
The nerve surgical procedure and injections were performed under aseptic conditions. The rats were anesthetized by IP injection of a mixture of ketamine (80 mg/kg) and xylazine (5 mg/kg). The sciatic nerve was exposed on the left side of the middle thigh and crushed with a pair of forceps (Dumont #5 forceps; Fine Science Tools, Foster City, CA) held tightly for 30 sec twice at a 1-min interval (Seo et al., 2006). The animals under anesthesia were then placed for 1 h on a water-circulating heating pad maintained at 37°C, then returned to their home cages. Running exercise for the TMT group was begun 12 h after surgery, and sedentary animals were left, in pairs, undisturbed in their standard plastic cages (42 × 28 ×18 cm3) for the same period of time.
TMT and drug administration
The treadmill device consisted of five moving platforms in parallel and was equipped with grids in order to motivate the rats to run. The intact rats were assigned into low-intensity and high-intensity TMT groups (LITMT and HITMT, n = 6 for each group), which was defined by their running speed on a moving horizontal platform (13 cm wide × 100 cm length) during the exercise period. The animals in the LITMT and HITMT groups were subjected to running at speeds of 8 m/min and 36 m/min, respectively, for 30 min twice a day (at 8
For inhibition experiments of ERK1/2 activity in vivo, the TMT animals were further divided into PD98059-treated (Cell Signaling, Danvers, MA) and DMSO vehicle-treated groups. PD98059 is known to be a potent, selective and cell-permeable inhibitor of MAP kinase kinase (MEK; Alessi et al., 1995). Immediately after crush injury, PD98059 (5 μL of 20-mM stock in 100% DMSO), or the equivalent volume of 100% DMSO, was injected focally into the crush site using a Hamilton syringe (Innovative Labor Systeme, Stuetzerbach, Germany). Expression levels of phosphorylated c-Jun, the rate of Schwann cell proliferation, and axonal regeneration were determined 1–7 days after a single dose of drug administration. The animals with drug administration did not show any behavioral or histological abnormalities.
The animals were sacrificed 1, 3, 7, and 14 days after nerve injury. They were deeply anesthetized with a mixture of ketamine and xylazine, and the sciatic nerve was dissected, immediately frozen, and kept at −70°C until use. For some experimental purposes, the sciatic nerve was prepared by dividing the nerve into the 5- or 10-mm-long segments proximal or distal to the injury site.
Immunohistochemistry
Nerve segments were embedded and frozen at −20°C. Longitudinal or transverse sections (20-μm thick) were cut on a cryostat and mounted on positively charged slides (Fisher Scientific, Pittsburgh, PA). For immunofluorescence staining, sections were fixed with 4% paraformaldehyde and 4% sucrose in PBS at room temperature for 40 min, permeablilized with 0.5% nonidet P-40 in PBS, and blocked with 2.5% horse serum and 2.5% bovine serum albumin for 4 h at room temperature. The sections were incubated with anti-GAP-43 antibody (rabbit polyclonal, 1:600; Santa Cruz Biotechnology, Inc., Santa Cruz, CA), anti-neurofilament-200 antibody (NF-200, rabbit polyclonal antibody, 1:700; Sigma, St. Louis, MO, or mouse monoclonal antibody, 1:600, Santa Cruz Biotechnology), anti-phospho-ERK1/2 antibody (mouse monoclonal antibody, 1:400; Cell Signaling, rabbit polyclonal antibody, 1:800, Cell Signaling), or anti-phospho-c-Jun antibody (rabbit polyclonal antibody, 1:400; Cell Signaling). Then they were incubated with rhodamine-goat anti-rabbit secondary antibody (1:10,000; Molecular Probes, Eugene, OR) or fluorescein-goat anti-mouse antibody (1:10,000; Molecular Probes) in 2.5% horse serum and 2.5% bovine serum albumin for 1 h at room temperature, and cover-slipped with gelatin mount medium. The sections were viewed with a fluorescence microscope (Nikon model E-600; Nikon, Kawasaki, Japan), and the images were captured with a digital camera, and analyzed using Adobe Photoshop Software (version 5.5; Adobe, San Jose, CA).
Proliferating Schwann cells in the injured sciatic nerve were analyzed by labeling with 5-bromo-2-deoxyuridine reagent (BrdU). In-vivo BrdU labeling of proliferating cells was performed as described by the manufacturer (Roche, Basel, Switzerland). Briefly, 36 h after crush injury, 5 μL of BrdU reagent (50 μg/mL) was injected into the injury site. The nerve sections (20-μm thick) were prepared after BrdU reaction for 48 h, fixed in 50-mM glycine/ethanol solution, and incubated for 15 min with trypsin solution (0.05% trypsin and 0.05% CaCl2 in PBS) at 37°C. The sections were treated with 2 N HCl for 20 min at 37°C, neutralized with PBS for 15 min, and stained with anti-BrdU-FLUOS antibody. Then microscopic images at 200 × magnification (290 μm × 365 μm) were chosen from the longitudinal sections at 500-μm intervals from the crush site, and the mean number of BrdU-positive cells per animal was counted in three or four nonconsecutive sections. The sections chosen for analysis were stereologically identified to be positioned at the central zone of the nerve by cryostating.
Primary culture of DRG sensory neurons and Schwann cells
Primary DRG sensory neurons were prepared from the lumbar L4–L6 levels of adult rats, and Schwann cells were prepared from the sciatic nerve as described previously (Han et al., 2007). For cell preparation for Western blot analysis, dissociated Schwann cells (5 × 106 cells) were plated in 60-mm culture dishes precoated with a mixture of poly-L-ornithine and laminin. After changing the medium to DMEM containing 5% FBS, 5% horse serum, 200 mM glutamine, and 1% penicillin/streptomycin 12 h later, the cells were cultured for 48 h. To observe neurite outgrowth of DRG neurons, the cells were cultured on the precoated cover-slips (1 × 105 cells per well on 24-well plates) for 48 h, and used for immunofluorescence staining with anti-βIII-antibody (TUJ1, rabbit polyclonal antibody, 1:500; Cavance, Berkeley, CA). Digital images of neuronal processes were captured and transferred to Adobe Photoshop. The number and length of neurite processes exhibiting clear outgrowth (longer than the cell body size) were analyzed using i-Solution software (Image and Microscope Technology, Goleta, CA). Mean neurite length was determined by analyzing at least 50 sensory neurons which were randomly selected from each experiment.
Schwann cells (1 × 105 cells per well on 24-well plates) were analyzed by immunofluorescence staining with anti-S100β antibody (1:300; Dako, Glostrup, Denmark), and anti-phospho-ERK1/2 antibody (1:800, mouse monoclonal antibody; Cell Signaling). Cellular nuclei were stained with 2.5 μg/mL of Hoechst 33258 (bis-benzimide; Sigma) for 10 min before final washing with 0.1% Triton X-100 in PBS.
In-situ hybridization
The rats were deeply anesthetized and perfused transcardially with phosphate-buffered 4% paraformaldehyde. Cryoprotected spinal cord tissues in 15% sucrose in PBS were sectioned at 20 μm, and then exposed to the following solutions: 4% paraformaldehyde (10 min), DEPC-treated water (rinse), 0.1 M triethanolamine (TEA, pH 8.0; rinse), 0.1 M TEA plus acetic anhydride (0.25% for 10 min; Sigma), 2X SSC (twice, rinse), and an ethanol series (50%, 70%, 95%, and 100%, 3 min each; dehydration). Both sense and antisense 35S-labeled GAP-43 cRNA probes were prepared using an in-vitro transcription protocol (Promega, Madison, WI) as described elsewhere (Namgung and Routtenberg, 2000). Then the probe (1 × 104 cpm/μL of in-situ hybridization buffer) was applied to the slides and incubated at 60°C for 16 h. The slides were washed in 2X SSC (twice for 10 min each), RNase A solution (20 μg/mL in 500 mM NaCl and 10 mM Tris-HCl [pH 7.4] for 30 min at room temperature), 2X SSC (twice for 10 min each), 0.1X SSC (60°C for 2 h), 0.5X SSC (twice for 10 min each), and ethanol (50%, 70%, 95%, and 100%, each containing 0.3 M ammonium acetate). All wash solutions contained 10 mM 2-mercaptoethanol and 1 mM EDTA, except for the RNase A solution, 0.5X SSC washes, and ethanol solutions. The slides were exposed to X-ray film for 1–2 days for autoradiography, and GAP-43 mRNA signals on the film were scanned as negatives using a film scanner (Nikon Super Coolscan 4000ED), and further analyzed by i-Solution software.
Western blot analysis
The nerve segments were washed with ice-cold PBS, and sonicated under 400 μL of Triton lysis buffer (20 mM Tris [pH 7.4], 137 mM NaCl, 25 mM β-glycerophosphate [pH 7.14], 2 mM sodium pyrophosphate, 2 mM EDTA, 1 mM Na3VO4, 1% Triton X-100, 10% glycerol, 5 μg/mL leupeptin, 5 μg/mL aprotinin, 3 μM benzamidine, 0.5 mM DTT, and 1 mM PMSF). The cell lysate from cultured Schwann cells was similarly prepared as described elsewhere (Han et al., 2007). Protein (15 μg) was used for Western blot analysis using anti-GAP-43 antibody (1:300, rabbit polyclonal antibody; Santa Cruz Biotechnology), anti-total ERK1/2 antibody (1:3000, rabbit polyclonal antibody; Cell Signaling), anti-phosphorylated ERK1/2 antibody (1:3000, rabbit polyclonal antibody; Cell Signaling), anti-phosphorylated c-Jun antibody (1:1000, rabbit polyclonal antibody; Cell Signaling), anti-actin antibody (1:3500, mouse monoclonal antibody; ICN Biomedicals, Cleveland, OH) and horseradish peroxidase-conjugated secondary antibodies (1:1000, goat anti-rabbit antibody; Santa Cruz Biotechnology, or sheep anti-mouse antibody; Amersham Biosciences, Piscataway, NJ). The Western blotting procedure has been described elsewhere (Namgung and Xia, 2000), and quantitative analysis of protein bands in the autoradiographic images was determined using i-Solution software.
Retrograde tracing technique
For retrograde tracing of DRG sensory neurons and motor neurons, adult rats (n = 10) were assigned to intact control (n = 2) and sedentary and TMT groups (n = 4 each). The sedentary and TMT animals were subjected to sciatic nerve crush injury, and fluorescent lipophilic carbocyanine dye l,l′-dioctodecyl-3,3,3′,3′tetramethylindocarbocyanine perchlo-rate (DiI; 5 μL of 3% in DMSO) was applied to the region 10 mm distal to the injury site with a micropipette as described previously (Seo et al., 2006). At 12 hours later, TMT group animals were subjected to running exercise for 3 days. The training protocol and treatments were essentially the same as those described above. The animals were sacrificed, and the DRG at L4 and L5, and the spinal cord between the lower thoracic and upper lumbar levels were dissected. The total number of DiI-labeled cells from three nonconsecutive sections (20 μm thick) per animal was counted, and the mean percentage of labeled cells in the sedentary and the TMT groups was compared to those of intact animals. Cell counting analysis was conducted by an examiner blinded to the experimental treatment conditions.
Statistical analysis
All data were presented as mean ± standard error of the mean (SEM). Statistical analysis for protein band intensity and other results was done using Student's t-test or one-way and two-way ANOVA using SPSS software (version 12.0; SPSS, Inc., Chicago, IL). Statistically significance was set at p < 0.05, < 0.01, or < 0.001.
Results
Low-intensity, but not high-intensity, TMT facilitates both neurite outgrowth of DRG sensory neurons and Schwann cell proliferation, and increases GAP-43 expression in injured sciatic nerves
To examine the effect of low- and high-intensity TMT on peripheral nerve cell activation, cultured DRG sensory neurons and Schwann cells in the sciatic nerve were prepared from rats after 14 days of TMT performance. Neurite length of DRG neurons labeled with neuron-specific-βIII-tubulin was significantly increased after 2 days of culture in the low-intensity TMT group compared to the high-intensity TMT group or intact controls (Fig. 1A). Similarly after 2 days of culture, the number of S100β-stained Schwann cells in the low-intensity TMT group was significantly higher than that in the high-intensity TMT group or control group (Fig. 1B). These data suggest that low-intensity TMT was effective for increasing neurite outgrowth and Schwann cell proliferation. Thus we used the low-intensity TMT paradigm (“TMT” hereafter) for the rest of this study.

Low-intensity TMT potentiates increased neural activity in peripheral nerves. After 14 days of TMT, DRG sensory neurons and Schwann cells in the sciatic nerve were cultured for 2 days and identified by immunofluorescence staining with anti-βIII-tubulin and anti-S100β antibodies, respectively. (
We then examined the effects of TMT on regenerative responses by measuring expression levels of GAP-43. GAP-43 protein was induced in the sciatic nerves at 3, 7, and 14 days post-crush (dpc) and further elevated by TMT (Fig. 2A). In-situ hybridization analysis showed increased GAP-43 mRNA signals in the ventral horn in the spinal cord between T12 and L1, where the sciatic motor neuron cell bodies are located (Fig. 2B). Quantitative comparison of GAP-43 mRNA signals in the ipsilateral ventral area showed the largest increase (116%) in TMT compared to sedentary controls at 14 days after injury. Images of the immunohistochemistry sections in the ventral horn area showed elevated levels of GAP-43 protein in motor neurons in the TMT group that were higher than those seen in sedentary control (Fig. 2C).
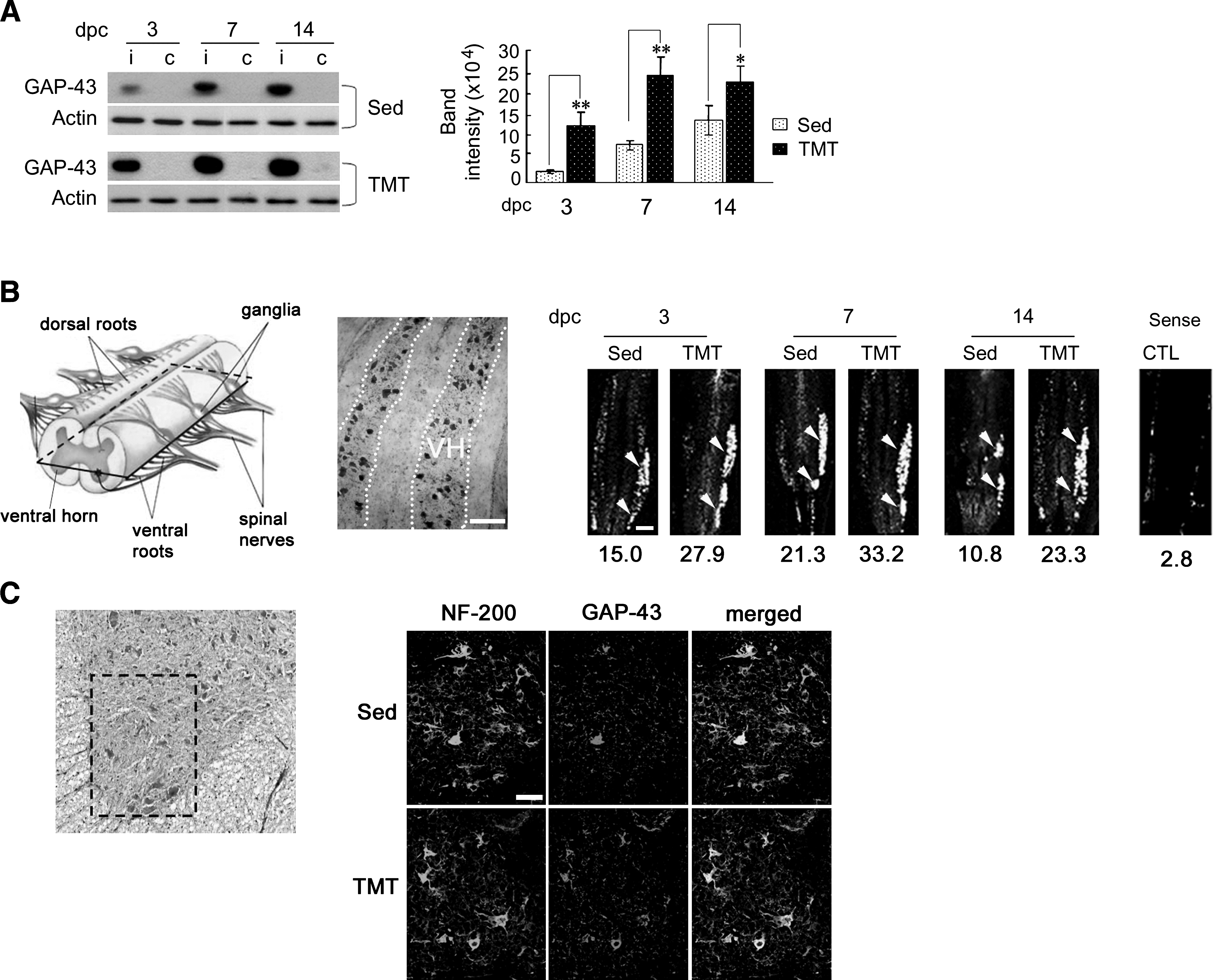
TMT upregulates expression levels of GAP-43 mRNA and protein induced by sciatic nerve injury. (
TMT upregulates ERK1/2 activation in Schwann cells after injury
Phospho-ERK1/2 protein levels were increased in the sciatic nerves at 1–7 dpc, and decreased to basal level 14 days later. In the TMT group, phospho-ERK1/2 protein levels were significantly higher than those in sedentary controls at 3 dpc, and decreased to basal levels at 14 dpc (Fig. 3A). Analysis of phospho-ERK1/2 protein levels revealed a characteristic difference between the proximal and distal distributions seen in the TMT and sedentary groups. In the TMT group, phospho-ERK1/2 protein levels in the distal stump, which were already higher than those in the proximal stump at 1 dpc were further increased at 3 dpc, whereas in the sedentary group phospho-ERK1/2 levels in the distal stump were lower than those in the proximal stump at 1 dpc, and increased to levels similar to those in the proximal stump at 3 dpc (Fig. 3B). The ratio of distal to proximal band intensity was significantly higher in the TMT group than in the sedentary group at 1 and 3 dpc, suggesting a facilitated distal shift of phospho-ERK1/2 by TMT.
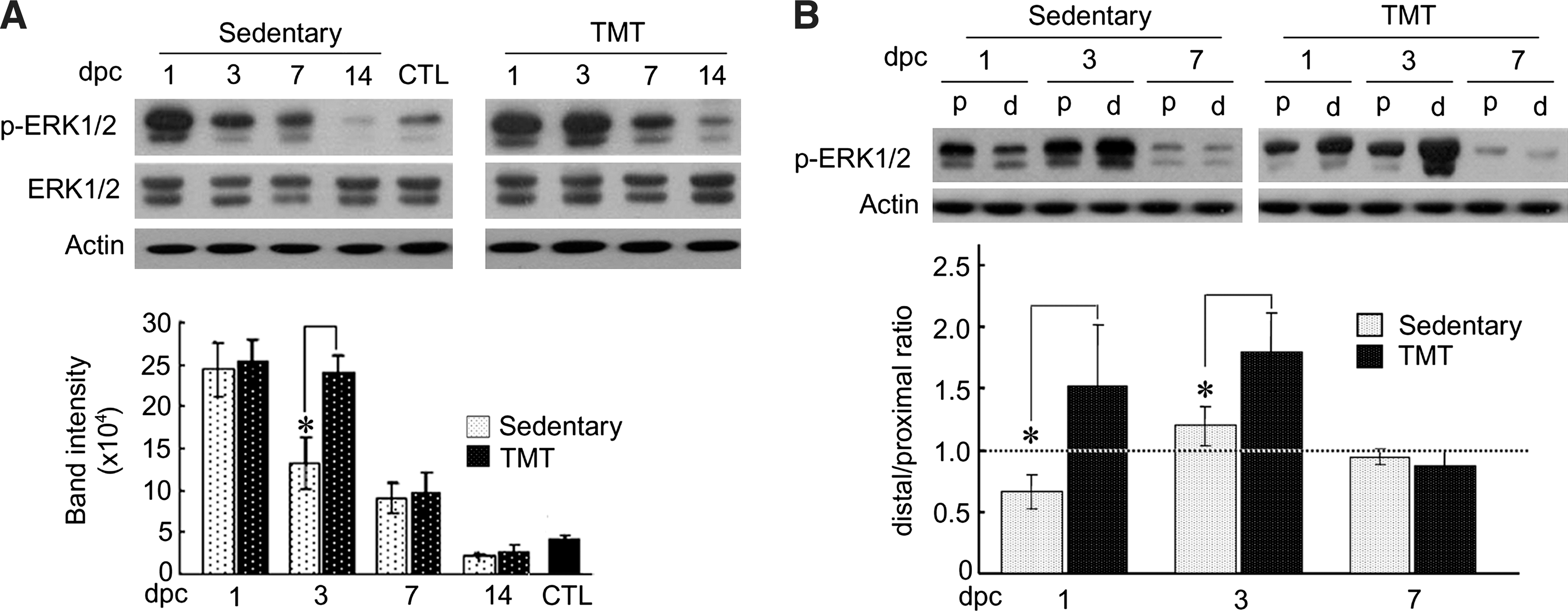
Regulation of ERK1/2 activity by TMT. (
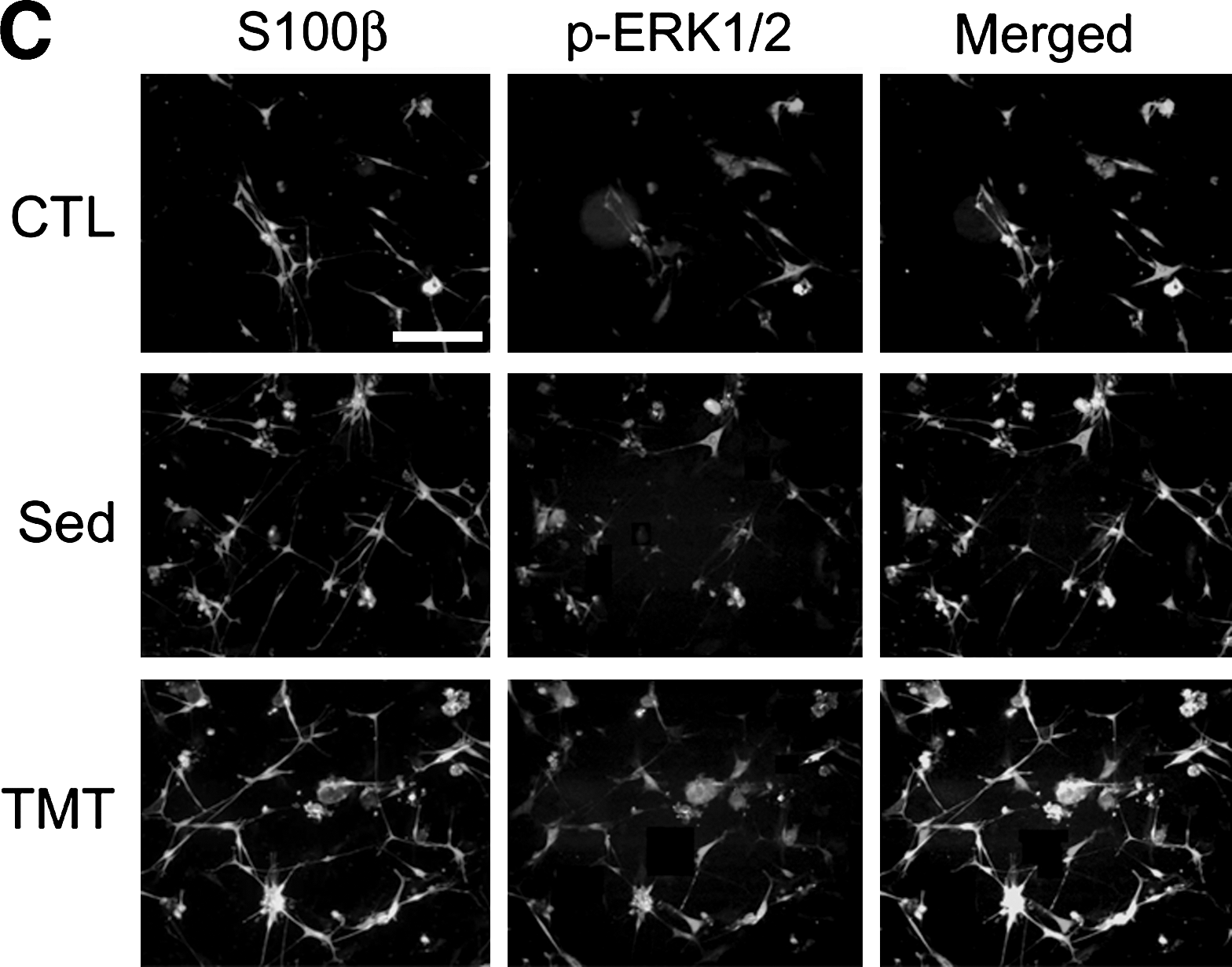
To examine the possibility of ERK1/2 activation in Schwann cells, we analyzed ERK1/2 protein in the sciatic nerve and cultured Schwann cells. Immunofluorescence staining of transverse sciatic nerve sections revealed that phospho-ERK1/2 protein was found in both the sedentary and TMT groups, with more intense signals seen in the TMT animals (Fig. 4A). Most of the phospho-ERK1/2 signals were not found in NF-200-labeled axons. To further demonstrate ERK1/2 activation in Schwann cells, we prepared primary culture cells from the sciatic nerves of rats that had undergone crush injury and had TMT for 3 days. Western blot analysis of the cultured Schwann cell lysate showed significant increases in phospho-ERK1/2 protein in the TMT group compared to the sedentary controls (Fig. 4B). Immunofluorescence staining showed that phospho-ERK1/2 protein signals largely overlapped with those of Schwann cell marker S100β, and were increased after sciatic nerve injury. Phospho-ERK1/2 protein levels were further elevated by TMT (Fig. 4C). In the sedentary and TMT groups, 74% and 92% of phospho-ERK1/2–expressing cells were S100β-positive, respectively. These data suggest that in the sedentary state, ERK1/2 activity may be produced by cell types such as macrophages and fibroblasts in addition to Schwann cells, but was increased more exclusively in activated Schwann cells by TMT.

TMT upregulates ERK1/2 activation in Schwann cells of the injured sciatic nerve. (
TMT upregulates levels of ERK1/2-mediated c-Jun phosphorylation and Schwann cell proliferation
To examine the possible link between phospho-ERK1/2 activity and c-Jun, we investigated time-dependent changes in c-Jun phosphorylation in the injured sciatic nerves. Phospho-c-Jun was not detected at 1 dpc in any of the groups, but was transiently increased at 3 dpc, followed by a decrease to basal levels at 7 dpc (Fig. 5A). At 3 dpc, the levels of phospho-c-Jun, similarly to those of phospho-ERK1/2, were significantly higher in the TMT than in the sedentary group. However, treatment with the MEK inhibitor PD98059 to block ERK1/2 activity decreased phospho-c-Jun to a basal level, suggesting ERK1/2 regulation of c-Jun phosphorylation in Schwann cells. Immunofluorescence images of the sciatic nerve sections from the TMT group prepared at 3 dpc showed that phospho-c-Jun was highly co-localized with phospho-ERK1/2, suggesting their activation in Schwann cells (Fig. 5B). It was further shown that most of the ERK1/2 and phospho-c-Jun proteins found in longitudinal nerve sections were not found in NF-200-stained axons (Fig. 5C).

TMT mediates ERK1/2 phosphorylation of c-Jun in the injured sciatic nerve. (
We further investigated the role of ERK1/2 on Schwann cell proliferation in vivo in the injured sciatic nerve. Proliferating Schwann cells were detected by fluorescein-conjugated 5-bromo-2-deoxyuridine reagent (BrdU) reagent. The Schwann cell counts along the axonal length up to 2.5 mm distal to the injury site were much higher in the TMT group than in sedentary controls (Fig. 6). PD98059 treatment of the TMT group significantly decreased BrdU-labeled cells in the nerve, to levels below those of the sedentary group. Taken together, these data suggest that TMT-mediated ERK1/2 activation may be associated with Schwann cell proliferation via c-Jun phosphorylation.

ERK1/2 inhibition decreases the number of BrdU-labeled cells in the distal sciatic nerve. Proliferating Schwann cells were labeled in vivo by BrdU treatment of the sciatic nerve at 3 dpc. (
Axonal regrowth facilitated by TMT is inhibited by the blockade of ERK1/2 activity
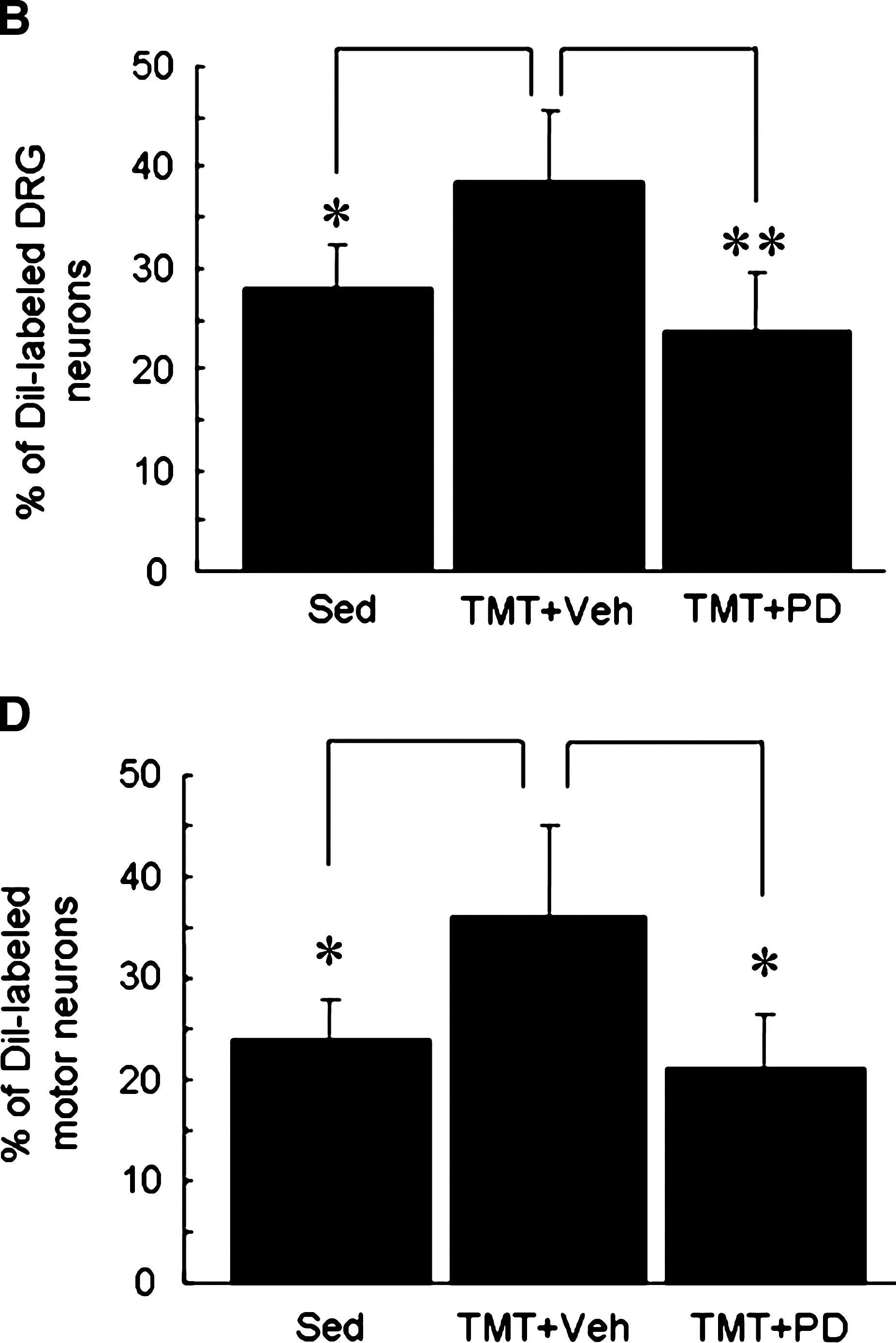
We have recently demonstrated, using anterograde and retrograde tracing techniques, that TMT facilitates axonal regeneration in the injured sciatic nerve (Seo et al., 2006). In the current article we examined the effect of ERK1/2 activity on axonal regeneration. Immunofluorescence images of NF-200–labeled axons showed that TMT for 3 days increased the number of fibers in the nerve segments 3 mm and 5 mm distal to the injury site (Fig. 7). PD98059 administration into the injured sciatic nerve significantly delayed axonal extension to the distal area compared to vehicle-treated TMT group. We further assessed axonal regeneration by retrograde DiI-labeling of DRG sensory neurons and spinal cord motor neurons after sciatic nerve injury (Fig. 8). DiI-labeled sensory neurons and motor neurons were significantly increased in the TMT group compared to the sedentary group, but the cell counts were significantly decreased by PD98059-inhibitor treatment.
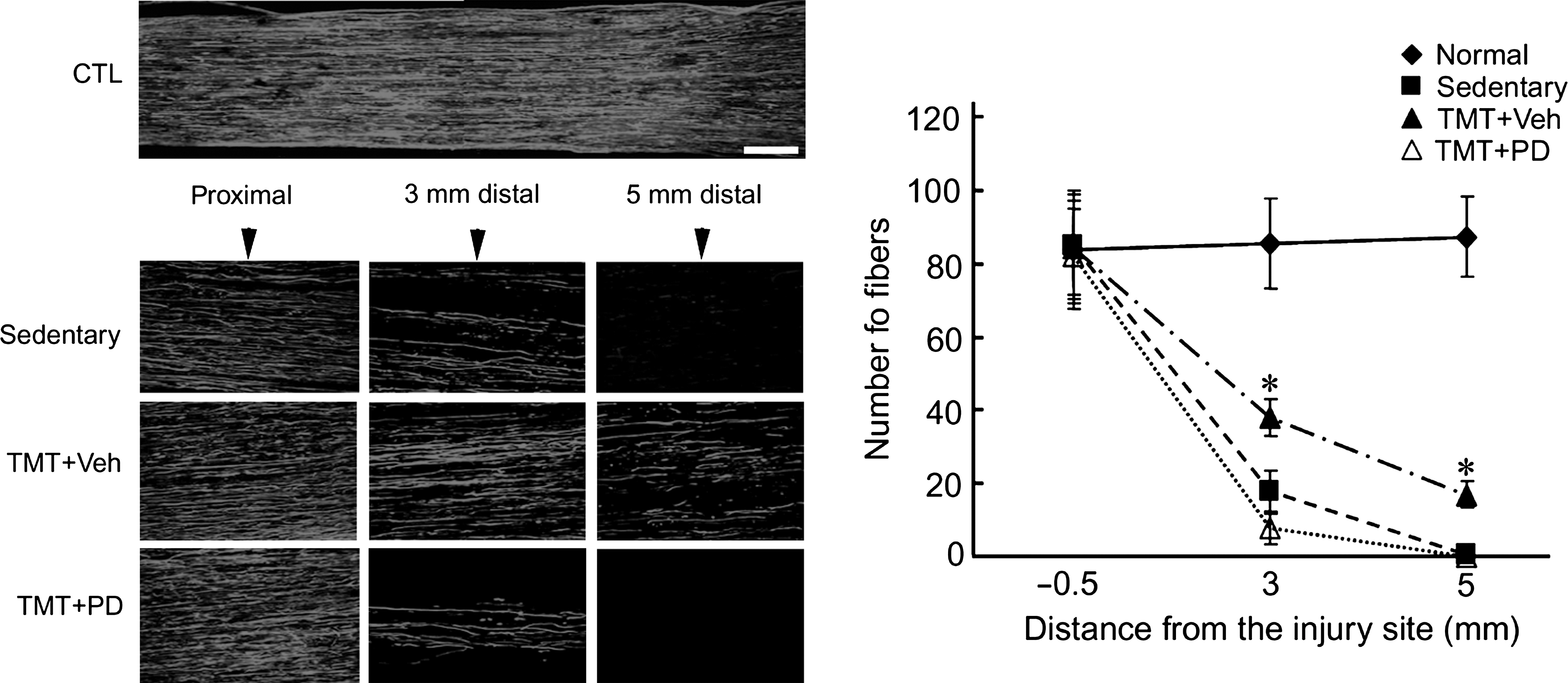
Axonal regeneration facilitated by TMT is retarded by ERK1/2 inhibition caused by PD98059 treatment. Sciatic nerves were prepared at 3 dpc and regenerating axons were visualized by immunofluorescence staining of NF-200 protein. Elongated axons were observed in the DMSO-treated TMT group (TMT + Veh) 3 and 5 mm distal to the injury site, but they were greatly decreased by PD98059 treatment (TMT + PD). In each experiment, the mean number of fibers was counted 0.5 mm proximal and 3 and 5 mm distal to the injury site from three or four nonconsecutive sections (marked with vertical arrows). There was a significant difference between the vehicle-treated and PD98059-treated TMT groups (n = 6, F = 31.6, df = 1,10; p < 0.001 by two-way ANOVA), and there was also a significant difference between the same two groups at distances of 3 and 5 mm (*p < 0.001 by one-way ANOVA; scale bar = 20 μm).

DiI retrograde labeling showing the effects of PD98059 treatment on axonal regeneration after sciatic nerve injury. After 3 days of TMT or sedentary treatment, DiI-labeled cells were analyzed in the DRG (
Discussion
Although recent studies have shown that exercise training can promote sensory and motor activity after nerve injury, the precise molecular mechanisms of the stimulation elicited by physical activity on an intracellular level are not known. Our data suggest that ERK1/2 activation induced by TMT in proliferating Schwann cells may contribute to facilitating axonal outgrowth after sciatic nerve injury. TMT increased ERK1/2 activity in the injured nerve tissues both in in-vivo and in cultured Schwann cells.
It has been reported that preconditioning of animals by sciatic nerve injury or by voluntary exercise induced neurite outgrowth in cultured DRG sensory neurons (Smith and Skene, 1997; Molteni et al., 2001). Similarly, increased proliferation and migratory activity in cultured Schwann cells derived from injured sciatic nerves in rats was also reported (Han et al., 2007). These studies support the notion that axonal regeneration–associated in-vivo stimulation of experimental animals may be retained to a certain extent in cultured cells. In the present study, TMT increased neurite outgrowth of DRG sensory neurons and Schwann cell proliferation, and upregulated ERK1/2 activation in cultured Schwann cells from injured sciatic nerve.
One intriguing question is how TMT in the rats could lead to ERK1/2 activation in Schwann cells and nerve regeneration. Retrograde signaling of skeletal muscle activity into Schwann cells is one pathway to consider. Glial cell line–derived neurotrophic factor (GDNF) expression is upregulated in skeletal muscle cells and Schwann cells by physical activity, and GDNF is secreted extracellularly to activate paracrine/autocrine receptors on Schwann cells (Henderson et al., 1994; Dupont-Versteegden et al., 2004; Bohn, 2004). It has been shown that GDNF interacts with neural cell adhesion molecule associated with GPI-linked GDNF family receptor α1 in Schwann cells, and also activates Fyn/FAK and ERK1/2 pathways (Paratcha et al., 2003; Iwase et al., 2005). Furthermore, GDNF stimulates Schwann cell migration (Paratcha et al., 2003), which may support axonal regeneration (Han et al., 2007). Another possible explanation may be descending inputs associated with exercise-induced brain plasticity. Enhanced brain activity by training may strengthen peripheral axonal activity through neurotrophic signaling, including BDNF (Cotman and Berchtold, 2002), yet how the descending neuronal inputs to injured peripheral nerves activate ERK1/2 in Schwann cells remains to be explored. Since BDNF in cerebellar granule neurons can increase ERK1/2 activity, leading to an enhancement of cAMP-dependent stimulation of neurite outgrowth (Gao et al., 2003), the autocrine and paracrine actions of BDNF via p75 or trkB receptors in Schwann cells in the injured sciatic nerves may be the molecular events connecting TMT and ERK1/2 activation. Finally, increased ERK1/2 activity in Schwann cells may cause nerve regeneration by downregulating CNTF expression (Abe et al., 2001), or by activating Cdk pathways (see below).
Given that TMT alone stimulated Schwann cell proliferation (see Fig. 1), phospho-ERK1/2 induced in Schwann cells after TMT may be involved in their proliferation, which may also be augmented by nerve injury. In the injured nerve area, Schwann cells undergo proliferation and migration at an early stage, and are subsequently differentiated to myelinate axons (Bunge et al., 1986). We have recently found that TMT upregulated Cdc2 kinase activity in Schwann cells (Seo et al., 2006), which was identified as a key player in cell migration (Han et al., 2007). Although there is no direct evidence of molecular cross-talk between Cdc2 and ERK1/2 kinase, ERK1/2 phosphorylation of cyclin B1, a principal Cdc2 activator, was implicated in epistatic regulation of the signaling pathway (Walsh et al., 2003; Yuan et al., 2002). Based on the functional similarity of ERK1/2 and Cdc2 in axonal regeneration after TMT, we speculate that both of these kinase pathways contribute to Schwann cell proliferation and migration.
Our data further showed that phospho-c-Jun protein at first was strongly induced in injured sciatic nerves after TMT, but then was inhibited by the blockade of ERK1/2 activity. c-Jun, which is primarily phosphorylated by c-Jun N-terminal kinase, can also be phosphorylated by ERK1/2 during PC 12 cell differentiation (Leppa et al., 1998). It was reported that expression and phosphorylation of c-Jun by the ERK pathway within the neurons were required for neurite outgrowth of cultured DRG neurons and axonal regeneration in injured peripheral nerves (Raivich et al., 2004; Lindwall et al., 2004). Our histochemical examination revealed weak phospho-ERK1/2 signals within the sciatic nerve axons and DRG sensory neurons at L4–L6. Moreover, inhibition of Schwann cell ERK1/2 activity blocked phospho-c-Jun expression, and also resulted in inhibition of Schwann cell proliferation and delayed sciatic nerve regeneration. While this observation suggests that Schwann cells are the major site of phospho-ERK1/2 activation by injury, the possibility of transient induction of ERK1/2 activity along the regenerating axonal length, particularly at an earlier time point, cannot be excluded. The current study showed that some phospho-ERK1/2 and phospho-c-Jun ERK1/2 signals were found in NF-200-stained axons at 3 dpc (Fig. 5C), and it was reported that phospho-ERK1/2 in cultured neurons exposed to injury signals were transported in a retrograde fashion, and may reside either in the axon or the cell body within 24 h (Perlson et al., 2005). ERK1/2 activity, which is induced 3–7 days after injury, may contribute to initiating Schwann cell activation in a different portion of the distal regenerating nerve, while activity induced earlier in the neuron may promote axonal elongation. Since ERK1/2 activity is involved in physiological activation of muscle cells as well as neural cells, proper ERK1/2 regulation by different cell types would be important in developing therapeutic strategies for enhanced peripheral nerve regeneration after injury.
Footnotes
Acknowledgments
This work was supported by grants from the Korea Research Foundation (C00158 to U. Namgung and G00035 to T.B. Seo). We also wish to thank Aryeh Routtenberg (Northwestern University) for providing GAP-43 cDNA, and David Elzi (University of Texas San Antonio).
Author Disclosure Statement
No competing financial interests exist.