
Abstract
Electrical stimulation has been shown to enhance peripheral nerve regeneration after nerve injury. However, the impact of electrical stimulation on motor functional recovery after nerve injuries, especially over long nerve gap lesions, has not been investigated in a comprehensive manner. In the present study, we aimed to determine whether electrical stimulation (1 h, 20 Hz) is beneficial for motor functional recovery after a 10 mm femoral nerve gap lesion in rats. The proximal nerve stump was electrically stimulated for 1 h at 20 Hz frequency prior to nerve repair with an autologous graft. The rate of motor functional recovery was evaluated by single frame motion analysis and electrophysiological studies, and the nerve regeneration was investigated by double labeling and histological analysis. We found that brief electrical stimulation significantly accelerated motor functional recovery and nerve regeneration. Although the final outcome, both in functional terms and morphological terms, was not improved by electrical stimulation, the observed acceleration of functional recovery and axon regeneration may be of therapeutic importance in clinical setting.
Introduction
E
Studies concerning the effect of ES on nerve regeneration have been performed only in animal models of crush injury, transected injury, or a short nerve gap (Brushart et al., 2002; Angelov et al., 2007). The inherent regenerative capacity of the nerve in animals could be so efficient over short nerve gaps that the effects of the ES may not be fully addressed. Thus, a longer nerve gap is needed to better illustrate the impact of ES on nerve regeneration and motor functional recovery.
The present study was designed to investigate the possible beneficial effect of ES (1 h, 20 Hz) on motor functional recovery from a long femoral nerve defect (10 mm), which was repaired with an autologous nerve graft. Single-frame motion analysis and electrophysiological test were used to evaluate the rate of quadriceps muscle function recovery (Irintchev et al., 2005; Ahlborn et al., 2007). Double labeling and morphological analysis were performed to investigate the effect of brief ES on nerve regeneration. We found that brief ES significantly accelerated motor functional recovery and nerve regeneration. Although the final outcome of motor function and nerve regeneration is not considerably improved by ES in the present study, this finding highlights the possibility that the brief ES may improve final outcome of nerve gap lesions in human patients in whom delayed target reinnervation is a factor limiting recovery.
Methods
Surgical procedures and electrical stimulation
Young adult male Sprague-Dawley rats (provided by Laboratory Animal Center of the Fourth Military Medical University, Xi'an, China), weighing from 200 to 220 g, were anesthetized by intraperitoneal injection of 3% sodium pentobarbital solution (40 mg/kg body weight). The left femoral nerve was exposed by making a skin incision and splitting the underlying muscles in the left lateral thigh. A 10 mm long segment of femoral nerve was excised and removed. The removed nerve segment was reversed 180° and reimplanted across the nerve defect with a 10/0 epineural suture of monofilament polyamide under 40× magnification (Fig. 1A and B).
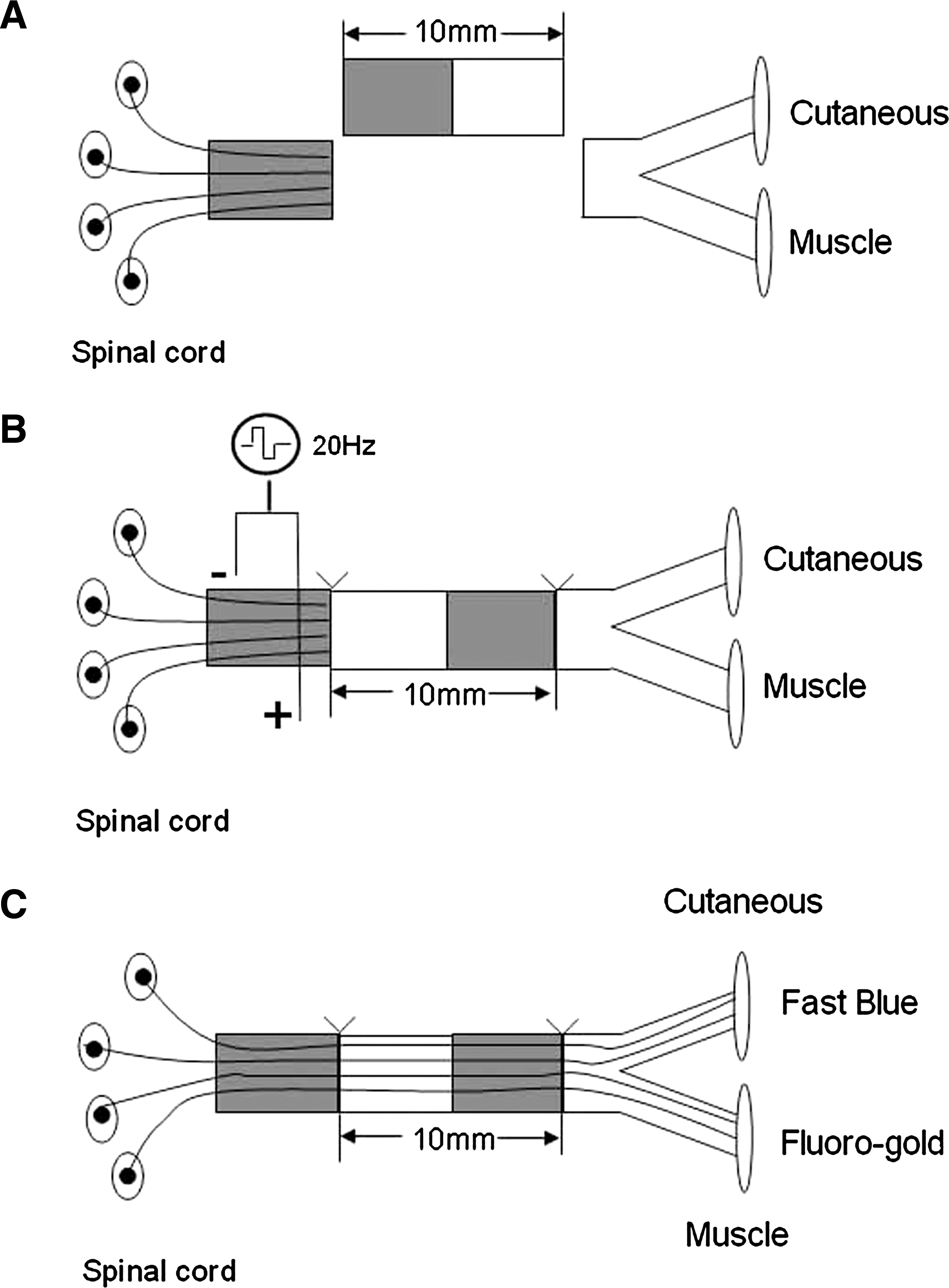
Diagrammatic of the regeneration model employed. (
In electrically stimulated rats, two insulated copper wires were bared of insulation for 2−3 mm, and the tip was twisted to form a loop to secure onto the proximal nerve stump to the lesion. A second electrode, used as an anode, was fixed to a muscle close to the nerve. The proximal nerve stump was then stimulated for 1 h by applying square 0.1 ms pulses at 20 Hz using an isolated pulse stimulator (Fig. 1). We chose a low stimulus frequency of 20 Hz because it is the physiological relevant frequency of hind limb motoneuron discharge. In sham-stimulated rats (control), the electrodes were positioned as in electrically stimulated rats, but no current was applied. During electrical or sham stimulation, the wound was covered by moistened paper to prevent drying of underlying tissues. Thereafter, the electrodes were removed, and the skin was closed with 6-0 stitches and liquid topical skin adhesive. Following surgery, the animals were retained in their cages and allowed to recovery for a period of 7 days before starting the functional testing.
Single motion analysis
Motor functional recovery was first assessed by single-frame motion analysis (SFMA) (Irintchev et al., 2005; Ahlborn et al., 2007), which allows precise evaluation of quadriceps muscle function during ground locomotion. Prior to operation, rats were trained to perform a beam-walking test in which the animals walked unforced from one end of a horizontal beam (1500 mm length × 52 mm width) towards its home cage located at the other end of the beam. A rear view of one walking trial per animal prior to the operation was captured with a high-speed camera (Nikon, Japan). The recording was repeated each week after nerve transaction. Selected frames in which the animals were seen in defined phases of the step cycles were used for measurements of two parameters: the foot base angle (FBA) and the heels-tail angle (HTA), as described previously (Irintchev et al., 2005). Both parameters are directly related to the ability of the quadriceps muscle to keep the knee joint extended during contralateral swing phases. As a relative measure of functional recovery at different time points after nerve injury, the stance recovery index (Ahlborn et al., 2007), which is a mean of recovery index (RI) for the HTA and the FBA, was applied. The index for each angle is calculated in percent as:
where
Electrophysiological assessment
At the 4-, 8-, and 12-week end points, electrophysiological studies were performed prior to tissue harvesting. After anesthesia was induced, the femoral nerve was exposed, and the nerve repair site was identified under a surgical microscope. The nerve repair area was insulated from the surrounding muscle with a rubber dam. A bipolar stimulating electrode was placed under the femoral nerve at a location 10 mm proximal to the lesion. A recording electrode was placed in the quadriceps muscle. Nerve stimulation (0.2 ms, 1 mA rectangular pulse wave) and recordings were performed with a PowerLab 4SP digital data acquisition system (AD Instruments, Castle Hill, Australia). Digitized data were stored in a personal computer and the peak amplitude of the compound muscle action potential (CAMP), CAMP latency of onset, and nerve conduction velocity (NCV) values were calculated.
Retrograde double labeling of motoneurons
Retrograde labeling and counting of back-labeled cells (Simova et al., 2006) were performed 1, 2, 4, 6, 8, 10, and 12 weeks after nerve repair. After the left femoral nerve was exposed, a piece of parafilm was laid underneath the nerve trunks and the two nerve branches were transected approximately 6 mm distal to the bifurcation. Retrograde fluorescent tracers were applied to the nerve ends in powder form: Fluorogold (FG, Fluorochome, Denver, CO) to the muscle branch, and Fast Blue (FB, EMS-Chemie, Stuttgart, Germany) to the cutaneous branch (Fig. 1C). At 30 min after dye application, the nerve stumps were rinsed with PBS and the wound was closed. Animals were kept for 96 h after tracer application to allow the retrograde tracers to travel back to the neuronal cell bodies.
The lumber spinal cords were cut longitudinally at 50 μm on a freezing microtome. Sections were serially mounted on glass slide, dried, and coverslipped. Each spinal cord section was visualized under UV fluorescence by an observer who was unaware of which branch had received FG or FB. Motoneurons containing both FG and FB throughout the cell body were viewed by changing the fluorescent light. The counting of split cells twice was corrected for by the method of Abercrombie and colleagues (1946). In each group, motoneurons were scored as projecting axons (1) correctly to the muscle branch, (2) incorrectly to the cutaneous branch, or (3) simultaneously to both branches.
Histological analysis
Femoral nerves were dissected from animals fixed by perfusion with formaldehyde and post-fixed in 1% osmium tetroxide in 0.1 M sodium cacodylate buffer (pH 7.3) for 1 h at room temperature, and then dehydrated and embedded in resin according to standard protocols. Transverse semi-thin (1 μm) and ultra-thin (50 nm) sections were cut from motor and sensory nerve branches 3−4 mm distal to the bifurcation. The semi-thin sections were stained with a 1% toluidine blue/1% borax solution prepared in distilled water and examined using Leica microscope. Ultra-thin sections were stained with uranyl acetate and lead citrate, and subsequently subjected to transmission electron microscope (H-600, Hitachi, Tokyo, Japan). Morphometric evaluations were completed by examiners who were blinded to the experimental groupings. Axon density and axon diameter were measured in a random sampling. The degree of myelination was estimated by the ratio of axon to fiber diameter (g ratio).
Statistical analysis
All data were obtained blinded with respect to treatment of the animals and are presented here as mean values with standard errors of mean (S.E.M.). Student t test and one-way analysis of variance (ANOVA) for repeated measurements with Tukey post hoc test were used to compare mean values. One rat from the sham stimulated group was excluded from all analysis because of poor regeneration as indicated by a low recovery index and number of back-labeled motoneurons at 12 weeks after nerve repair. Statistical significance was accepted at p < 0.05 level.
Results
ES accelerates motor function recovery
The knee extensor function of the quadriceps muscle was evaluated in rats prior to and during a 12-week period after nerve lesion using three parameters: the heels-tail angle, the foot base angle, and stance recovery index (Irintchev et al., 2005; Ahlborn et al., 2007). As revealed by the postoperative changes in those parameters, functional performance was impaired to the same extent in both electrically stimulated and control rats one week after operation (Fig. 2A and B). The rate of reversal of the injury-induced disability after the second week was considerably faster in the electrically stimulated rat than that in the control rats (Fig. 2A and B). As shown in Figure 2C, the maximal degree of recovery reached in sham stimulated rats (65% of the preoperative values at 12 weeks) was achieved 4 weeks earlier after ES. Notably, a tendency for a better outcome in electrically stimulated rats was found when stance recovery indices in those two groups were compared at the 12-week end point (74% vs. 65%, Fig. 2C), but no statistically significance was found (t test, p = 0.096, Fig. 2C).
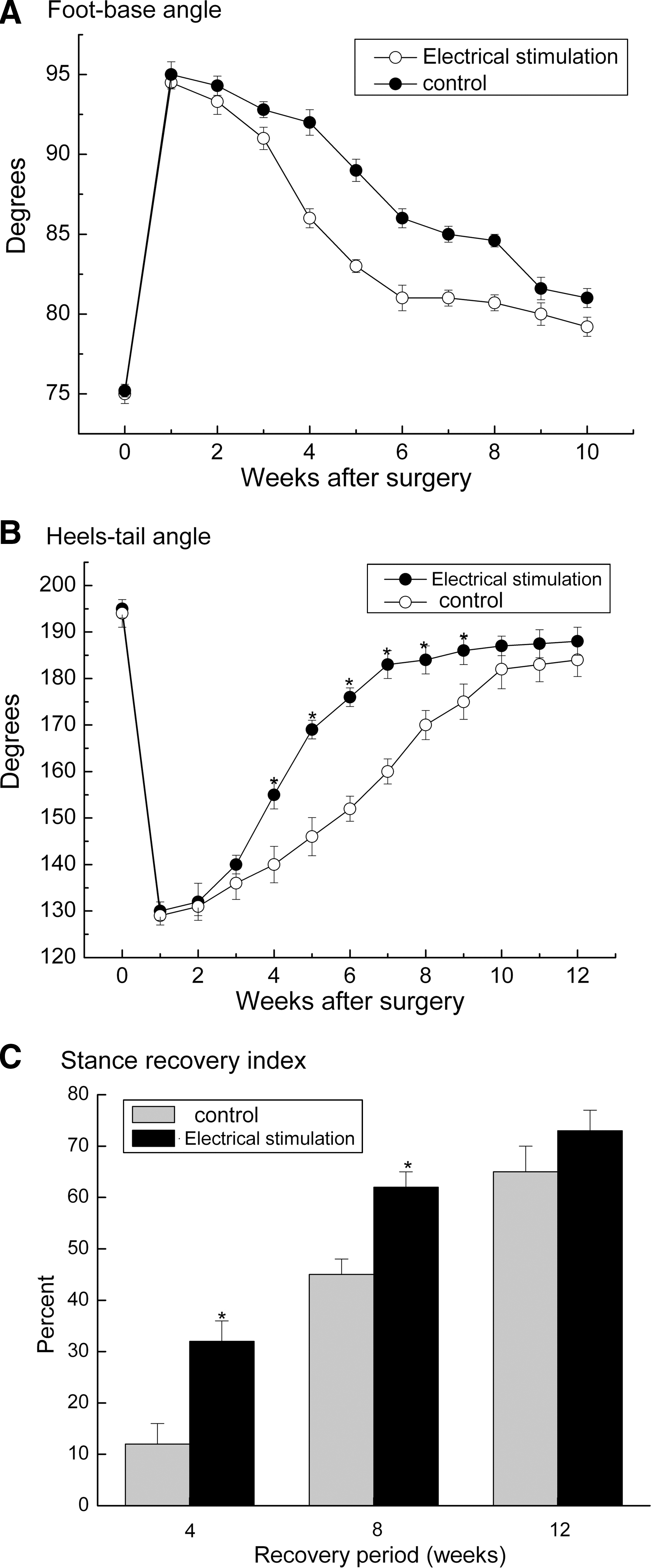
Time course and degree of motor functional recovery after femoral nerve lesion. Data are expressed as values ± S.E.M. of foot-base angle (
Comparison of electrophysiological parameters
Electrophysiological studies were performed to further investigate the beneficial effect of ES on motor functional recovery. The peak amplitude of the CAMP, CAMP latency of onset, and NCV were calculated from every rat (Table 1). Brief ES significantly lowered the CAMP latency of onset 4 weeks after surgery (Table 1, p < 0.05). Further, brief ES significantly enhanced the peak amplitude of the CAMP and NCV 4 and 8 weeks after ES (Table 1, p < 0.05). However, the three parameters in both stimulated and control rats were not significantly different 12 weeks after surgery and ES (Table 1, t test, p > 0.05).
Values are expressed as mean ± S.E.M.
p < 0.05 compared to sham stimulated group.
ES accelerates the speed of nerve regeneration
The impact of ES on autograft-repaired 10 mm femoral nerve gap was dramatic. We found that 1 h of electrical stimulation at 20 Hz accelerated the speed of nerve regeneration. All motoneurons regenerated their axons across the nerve gap within 6 weeks in the electrically stimulated rats, in contrast to the 10 − 12 weeks required in the control group (Fig. 3A).
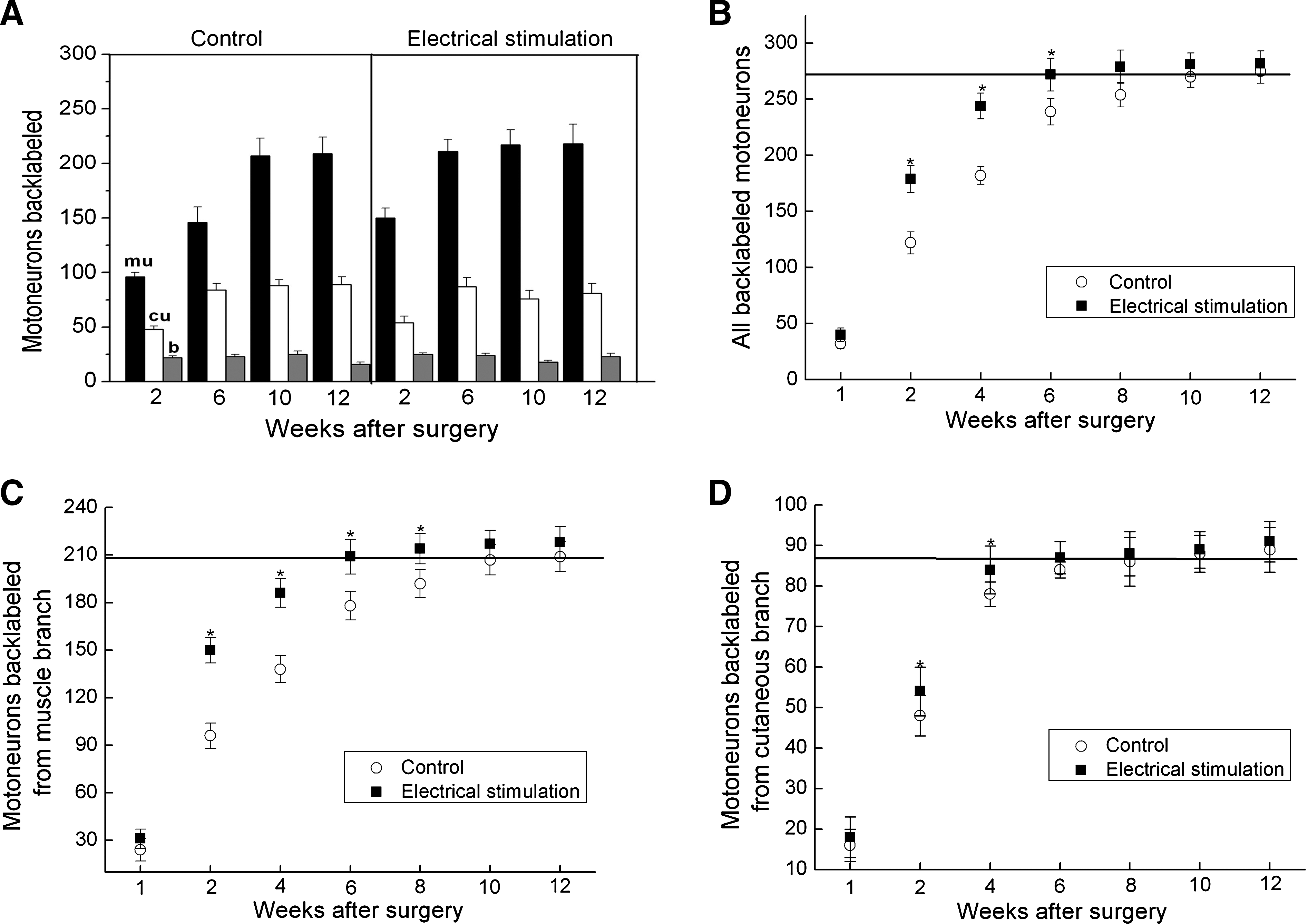
Effects of electrical stimulation on motor axonal regeneration and preferential motor reinnervation. (
Acceleration of nerve regeneration by ES was accompanied by accelerated preferential growth of these regenerating axons into appropriate muscle pathways but not into cutaneous pathways (Fig. 3B–D). The accelerated axonal regeneration in the electrically stimulated rats was clearly seen when the total number of motoneurons that regenerated and those that separately regenerated into the muscle branch and cutaneous branch were plotted as a function of time after femoral nerve lesion and surgical repair (Fig. 3B–D). As shown by the solid squares, brief ES accelerated axonal regeneration of all injured motoneurons by at least 4 weeks compared with the 10–12 week periods required for all motoneurons to cross the 10 mm nerve gap in the control group (Fig. 3B and C). The increase in the total number of regenerated motoneurons (Fig. 3B) was accounted for by a corresponding increase in the number of axons that regenerated into the appropriate muscle branch (Fig. 3C). The number of motoneurons that regenerated their axons into the cutaneous branch did not increase significantly between 4 and 12 weeks after surgery (Fig. 3D).
Morphological evaluation of nerve regeneration
Analysis of ultra-thin sections of the femoral nerve branches distal to the lesion revealed that regenerated axons had significantly larger diameters in the electrically stimulated rats compared with those of the control rats 4 and 8 weeks after surgery, but no significant difference in axon diameter was observed 12 weeks after surgery (Fig. 4A and B; Fig. 5).
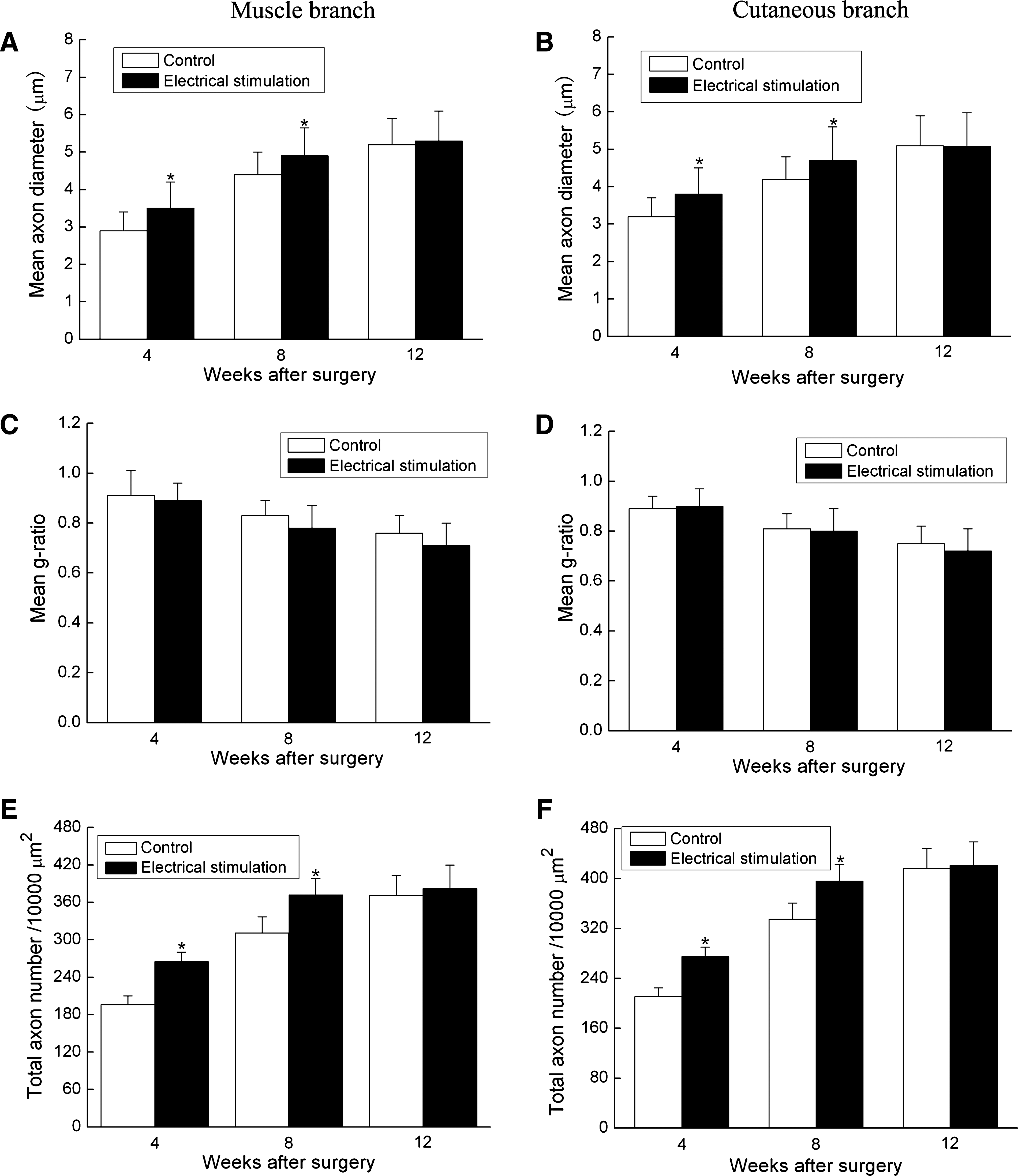
Summary of histomorphometry of distal portions for the control rats (open bars) and the electrically stimulated rats (black bars) at 4, 8, and 12 weeks after femoral nerve gap lesion and electrical stimulation. (

TEM of motor/cutaneous branch distal to autograft at 4 weeks (
Further investigations showed that ES significantly increased the number of myelinated axons in both the muscle branch and cutaneous branch 4 weeks and 8 weeks after surgery. However, 12 weeks after surgery, the number of myelinated axons was similar in both the electrically stimulated and the control groups (382 ± 27 vs. 371 ± 32, Fig. 4E; Fig. 5). Although the beneficial effect of ES on axons was as described above, it was noteworthy that the degree of myelination estimated by the g ratio (axon to fiber diameter) was similar in both groups at the predefined time points (4, 8, and 12 weeks after surgery, Fig. 4C and D; Fig. 5).
Discussion
Severe traumatic injury or invasive surgical procedures on a peripheral nerve can result in a gap between two nerve stumps. Autologous transplantation is presently the most common treatment for peripheral nerve lesions. Although autograft transplantation has been considered the clinical “gold standard” for nerve repair, it generally fails to achieve significant functional recovery after months of nerve regeneration. Over the past decades, multiple therapeutic strategies have been presented to promote nerve regeneration including magnetic field (Kelleher et al., 2006), ultrasound (Michlovitz, 2005), and electrical stimulation (Al Majed et al., 2000a, 2000b, 2004; Brushart et al., 2002, 2005; Geremia et al., 2007).
ES has been used to enhance peripheral nerve regeneration in the animal model of nerve injuries for some time (Brushart et al., 2002; Angelov et al., 2007). Applying depolarizing current to the damaged nerve has proven beneficial for regeneration after nerve injury in rabbits, rats, and mice (Nix and Hopf, 1983; Pockett and Gavin, 1985; Al Majed et al., 2000a, 2000b, 2004; Brushart et al., 2002, 2005; Geremia et al., 2007). However, the functional recovery was seldom addressed in those studies. In addition, we have to acknowledge that animal models of nerve injury have only included crush injury, transected injury, or a short nerve gap. The effect of ES on motor functional recovery over a long nerve defect has not been investigated in a comprehensive manner thus far.
In the present study, we investigated the impact of ES on motor functional recovery using a single-frame motion analysis (Irintchev et al., 2005; Ahlborn et al., 2007) after a 10 mm femoral nerve defect in rats. The single-motion analysis allows precise assessment of knee extensor function of the quadriceps muscle and is valuable for evaluating motor functional recovery after femoral nerve injuries (Irintchev et al., 2005; Ahlborn et al., 2007). We found that brief ES (1 h, 20 Hz) of the proximal stump leads to accelerated motor functional recovery in adult rats. When a 10 mm femoral nerve defect was bridged with an autologous nerve graft, the motor functional recovery in control rats reached maximum recovery by 10–12 weeks after surgery. However, 1 h of ES dramatically accelerated motor functional recovery by at least 4 weeks, as shown in the present study.
Electrophysiological study was performed to further investigate motor functional recovery after ES. Since the CAMP amplitude is directly proportional to the number of nerve fibers innervating the target muscle, this allows a further evaluation of motor functions (Chew et al., 2007). Here we recorded the CAMP peak amplitude, CAMP latency of onset, and NCV after femoral nerve stimulation. The electrically stimulated rats had a significant shorter CAMP latency (4 weeks), faster NCV (4 weeks and 8 weeks), and higher CAMP peak amplitude (4 weeks and 8 weeks) than the control rats. The electrophysiological data further confirmed that ES accelerated functional recovery when 1 h of ES (20 Hz) was applied immediately after nerve lesion and surgery.
ES has been shown to enhance nerve regeneration in animal models of crush injuries, transected injuries, or nerve gap lesions (Brushart et al., 2002; Angelov et al., 2007; Lu et al., 2008). In the present study, we found that ES not only enhanced axonal regeneration, but also accelerated the speed of nerve regeneration after a 10 mm nerve defect. The acceleration of nerve regeneration occurred with time concurrent with the accelerated motor functional recovery, and may partially explain the ES accelerated motor functional recovery in the present study. Although accelerated nerve regeneration was observed in the present study, we have to realize that the final outcome of nerve regeneration was not significantly enhanced by ES in the present study. It is possible that the inherent regenerative capacity of the nerve in rats was efficient enough to cross the 10 mm nerve gap when it was bridged with autologous nerve grafts. Thus, a larger gap in rats or a nerve gap in larger animals was needed to illustrate the impact of ES on final outcome of nerve regeneration and functional recovery.
In the present study, brief ES (1 h, 20 Hz) of the proximal nerve stump significantly accelerated the rate of motor functional recovery and nerve regeneration over a 10 mm femoral nerve defect. The mechanism underlying the accelerated motor functional recovery by ES is a complex process. It has been reported that brain-derived neurotrophic factor (BDNF) and its tyrosine kinase B (TrkB) receptors, as well as expression of the regeneration-associated proteins Talpha 1-tubulin and growth-associated protein-43 (GAP-43) were upregulated in motoneurons after brief ES (Al Majed et al., 2000a, 2004). These studies have indicated that TrkB-mediated neurotrophin signaling, argumented by membrane depolarization and subsequent Ca2+ influx into the cell bodies, and externalization of both TrkB and BDNF, also plays an important role in mediating the effects of ES (Brushart et al., 2002). The combined findings indicate that TrkB-related effects may be one possible explanation for the acceleration of both axonal regeneration and functional recovery by ES (Al Majed et al., 2000a, 2004; Brushart et al., 2002; Eberhardt et al., 2006; English et al., 2007). Despite this knowledge, we have to acknowledge that the mechanisms responsible for accelerated motor recovery may be more complicated. Identifying the differentially expressed regeneration-associated genes or proteins in the presence or absence of ES may help us better understand the mechanism for the beneficial effect of ES on nerve regeneration and motor functional recovery; this work is now being undertaken in our laboratory and will be summarized in subsequent reports.
In the present study, we showed that ES significantly accelerated motor functional recovery and nerve regeneration of a 10 mm femoral nerve defect in rats. Although the final outcome in functional recovery was not considerably improved by ES, the accelerated motor functional recovery may be of therapeutic importance in clinical settings. In comparison to small laboratory animals, reinnervation of peripheral nerves in humans is a slower process that requires months to be completed. Motor functional recovery in humans is limited by slow regeneration of axons across nerve lesion, progressive decline in regenerative capacity of axotomized neurons, and progressive deterioration of the distal growth environment (Brushart et al., 2002; Fenrich and Gordon, 2004). If the speed of nerve regeneration is accelerated by ES in humans, as described in rats in the present study, the regenerated axons may enter the distal stump and reach their target organs before the growth potential of the axotomized neurons and the permissive growth environment of the distal nerve stump deteriorate. Thus, it can be expected that accelerated nerve regeneration and functional recovery by ES may improve the final outcome of autograft-bridged nerve gap lesions to a considerably greater extent in larger animals, including humans.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (grants no. 30770571) and the National Hi-Tech Research and Development Program of China (863) (grant no. 2002AA216101). We thank technicians Lan Lifeng for her excellent technical assistance.
Author Disclosure Statement
No competing financial interests exist.