
Abstract
Despite reduced sympathetic activity below the level of a spinal cord injury (SCI), venoconstriction during autonomic dysreflexia increases venous return to the heart. Here, contractions of isometrically mounted tail veins from rats with spinal transection at T4 performed 8 − 10 weeks earlier are compared with those from sham-operated rats. After SCI, lumen diameter was reduced by ∼30% and the contractions evoked by electrical stimulation of the perivascular axons were larger than control. This augmentation of neurovascular transmission was not associated with enhanced sensitivity to α-adrenoceptor agonists or to adenosine-5′-triphosphate (ATP) although contractions to depolarization with K+ were larger after SCI. The percentage reduction in nerve-evoked contraction after SCI produced by the α1-adrenoceptor antagonist prazosin (10 nM) was unchanged but that by the α2-adrenoceptor antagonist rauwolscine (0.1 μM) was reduced. The relative contribution of P2-purinoceptors to nerve-evoked contractions after α-adrenoceptor blockade, revealed by adding suramin (0.1 mM), was unchanged. The greater depolarization-induced contraction and the reduced contribution of α2-adrenoceptors to nerve-evoked contraction suggest that changes in the venous smooth muscle underlie the potentiation of neurovascular transmission after SCI. Furthermore, the smaller lumen diameter after SCI will increase the pressure that the veins exert on the luminal contents when they are neurally activated.
Introduction
Loss of excitatory bulbospinal pathways controlling pre-ganglionic sympathetic vasoconstrictor pathways after SCI leads to a drop in arterial pressure that cannot be recovered by baroreceptor reflexes (Mathias and Frankel, 1999). Furthermore, the loss of baroreflex inhibition allows sensory inputs from below the lesion to activate vasoconstrictor neurones leading to autonomic dysreflexia. While this sympathoexcitation may be exacerbated by plasticity within the spinal cord (Hou et al., 2008; Weaver et al., 2006), augmentation of neurovascular transmission in arterial vessels may also contribute by producing a maintained and intense increase in peripheral resistance below the lesion (Stjernberg and Wallin, 1983; Teasell et al., 2000). In humans, the major underlying cause of this enhancement of neurovascular transmission is believed to be an increased responsiveness of vascular smooth muscle to nerve-released norepinephrine (NE) (Teasell et al., 2000). The idea that this mechanism may also be involved in enhanced venoconstriction during autonomic dysreflexia is supported by the larger responses of superficial foot veins produced by locally infused NE in SCI patients (Arnold et al., 1995). However it is not known whether SCI modifies the response of foot veins to NE released from sympathetic nerves, which is not necessarily mimicked by the contractions produced by exogenous NE (Hirst et al., 1996).
In recent experiments, we have demonstrated a marked enhancement of sympathetic nerve-evoked contractions in arteries isolated from below a chronic spinal cord transection in rats (Brock et al., 2006; McLachlan and Brock, 2006; Rummery et al., 2010; Yeoh et al., 2004a). While it is notable that the mechanisms underlying the enhancement differed between the vessels in various vascular beds, the findings in the tail artery are similar to the changes observed after decentralization of this vessel (Yeoh et al., 2004b). The simplest conclusion seems to be that decreased sympathetic activity following loss of bulbospinal inputs to the pre-ganglionic sympathetic neurons potentiates neurovascular transmission in all arteries below a spinal cord injury. Other factors, such as circulating vasoactive hormones, may play a role in discriminating between the reactions to the disturbed sympathetic activity in different vascular beds. Increased nerve-evoked contractions are likely to contribute to the autonomic dysreflexia that can be evoked by stimuli such as colorectal distension (Krassioukov and Weaver, 1995).
In the present study, we directly tested the hypothesis that a T4 spinal cord transection would result in an augmentation of neurovascular transmission in rat lateral tail veins and that this change results from a greater reactivity of the vascular muscle to neurotransmitters. The tail veins, which drain blood from the skin of the tail, are controlled by pre-ganglionic neurons located in T13-L2 spinal cord segments; spinal cord transection at T4 severs all their bulbospinal inputs without damaging the pre- or post-ganglionic sympathetic pathways. We determined the responses of segments of these veins to electrical stimulation of their perivascular nerves and the effects on these nerve-evoked contractions of the α-adrenoceptor antagonists prazosin (α1-selective) and rauwolscine (α2-selective), and the P2-purinoceptor antagonist suramin. In addition, we evaluated the responses of these veins to the α1-adrenoceptor agonists phenylephrine (PE) and methoxamine, to the α2-adrenoceptor agonist clonidine, to adenosine-5′-triphosphate (ATP), and to depolarization with K+. The reasons for studying responses to both PE and methoxamine are that only the former is a substrate for the neuronal NE transporter (NET) (Trendelenburg et al., 1970) and that decreased activity of this amine carrier contributes to the SCI-induced augmentation of nerve-evoked contractions of rat mesenteric arteries (Brock et al., 2006). The results suggest that changes in both vascular dimensions and neurovascular transmission interact to potentiate the response of veins to sympathetic activity.
Methods
All experimental procedures conformed to the Australian Code of Practice for the Care and Use of Animals for Scientific Purposes and were approved by the University of New South Wales Animal Care and Ethics Committee.
The spinal cords in female Wistar rats (8 − 9 weeks of age) were transected under anesthesia with ketamine (60 mg kg−1) and xylazine (10 mg kg−1) injected i.p. After infiltrating the back muscles with 0.3 mL of the long-acting local anesthetic Marcain (5 mg mL−1 bupivacaine hydrochloride; AstraZeneca, Melbourne, Australia), the spinal cord was exposed by a laminectomy over the T4 segment and cut with fine scissors. A piece of hemostatic gelatine foam was placed between the cut surfaces and another over the laminectomy site before applying oxytetracycline powder and closing the incision. Five mL of warm saline (0.9% NaCl) was injected i.p., and oxytetracycline (100 mg kg−1) and benzylpenicillin (90 mg kg−1) were injected s.c. The urinary bladder was emptied before and at frequent intervals after spinal transection. The animals were operated on a heating blanket; respiration and rectal temperature were monitored frequently until they recovered consciousness. Following surgery, the animals recovered quickly, moving about their cages within 2 h, and eating and drinking the following day. Urinary bladders were emptied manually at least four times a day for the first 1 − 2 weeks until they emptied automatically, and then the animals were monitored at least twice a day. The animals were caged in groups of two to four and maintained for 8 − 10 weeks (9.4 ± 0.1 weeks [mean ± SD], n = 6). In sham-operated controls, which were maintained for 8 weeks (8.1 ± 0.2, n = 6), the laminectomy was performed to expose the spinal cord at T4 and the dura was opened across the dorsal surface of the cord. All other treatment was identical to the spinally transected rats except for post-operative bladder care.
On the day of the experiment, animals were exsanguinated under deep anesthesia (80 mg kg−1 pentobarbitone, i.p.). The lateral veins on either side of the tail were carefully dissected from 2 − 5 cm below the base of the tail. The veins were maintained in physiological saline with the following composition (in mM): Na+, 150.6; K+, 4.7; Ca2+, 2; Mg2+, 1.2; Cl-, 144.1; H2PO4 -, 1.3; HCO3 -, 16.3; glucose, 7.8. This solution was gassed with 95% O2/5% CO2 (to pH 7.25) and warmed to 35–36°C.
Mechanical responses
Segments of vein ∼1.5 mm long were mounted isometrically between stainless steel wires (50 μm in diameter) in two four-chambered myographs (Multi Myograph model 610M, Danish Myo Technology, Aarhus, Denmark). Each chamber of the myographs contained 6 mL of physiological saline. The output from each myograph was recorded using a PowerLab recording system and the program Chart (ADInstruments, Bella Vista, Australia).
Because of the changes in diameter after SCI (see Results section), the basal conditions were normalized using Laplace's equation by converting the measured force to the effective transmural pressure (ETP), which scales the force to the lumen surface area of the vessel segment (mN/mm2) (Mulvany and Halpern, 1977). Responses to nerve stimulation and to agonists were also converted to ETP in the same way. To illustrate the effect of reducing lumen diameter, the contractions to nerve stimulation are also presented in Figure 2 as increases in wall tension (force/2 × artery segment length) (Mulvany and Halpern, 1977).
In preliminary experiments, it was confirmed that the increase in ETP produced by raising the extracellular K+ concentration to 60 mM (equimolar substitution of KCl for NaCl) was near maximal at the basal ETP of ∼2 mN/mm2 (∼15 mmHg; Fig. 1); this was used in all the experiments.

The relationship between basal effective transmural pressure (ETP) and the increase in ETP produced by increasing K+ concentration to 60 mM in tail veins (n = 5).
Electrical stimulation
Four vein segments from each rat were used to assess contractions to activation of the perivascular nerves. Electrical stimuli (0.2 msec pulse width, 15 V) were applied through platinum plate electrodes mounted along either side of the vein. In preliminary experiments, these stimuli produced supramaximal responses that were abolished by 0.5 μM tetrodotoxin, indicating that they were entirely due to the release of neurotransmitters from the perivascular nerves.
To construct a frequency-response curve, the veins were initially stimulated with single trains of 25 stimuli at 0.1, 0.3, 0.5, and 1 Hz. To assess the effects of neurotransmitter antagonists, the responses of the veins to 10 stimuli at 1 and 10 Hz and 100 stimuli at 1 Hz were compared before and during the application of the antagonists; the antagonists were in contact with the tissue for at least 20 min before changes to the responses were determined. Two vein segments were used to assess the contribution of NE to nerve-evoked contraction. Following measurement of responses in the absence of drugs, the effect of prazosin (10 nM; Sigma Chemical, Castle Hill, Australia) was assessed on one of these segments and the effect of rauwolscine (0.1 μM; Tocris Bioscience, Bristol, United Kingdom) on the other. Subsequently, the other adrenoceptor antagonist was added and their concentrations increased 10-fold (prazosin [0.1 μM] plus rauwolscine [1 μM]) to ensure blockade of all α-adrenoceptors. To assess the contribution of ATP to nerve-evoked contraction, another vein segment was treated with suramin (0.1 mM; Sigma) after the responses in the absence of drugs. Subsequently, prazosin (0.1 μM) and rauwolscine (1 μM) were applied and the effect of all three antagonists together was assessed. The fourth vein segment was used to determine time-dependent changes in the electrically evoked responses.
Chemical stimulation
For each rat, another four vein segments were used to assess changes in post-junctional reactivity to α-adrenoceptor agonists. Noncumulative concentration-response curves for the α1-adrenoceptor agonists PE (0.01 − 30 μM; Sigma) and methoxamine (0.01 − 30 μM; Sigma), and the α2-adrenoceptor agonist clonidine (0.001 − 3 μM; Sigma) were determined in separate vein segments by applying concentrations that increased by half-log increments, with the tissue exposed to each concentration for 5 min and then washed for 7 min before the addition of the next concentration. Sensitivity to PE was assessed both in the absence and in the presence of desmethylimipramine (DMI, 30 nM) to block NET. In addition, after washing out the highest concentration of α-adrenoceptor agonist studied, each tissue was exposed to ATP (0.1 mM; Sigma) for 2 − 4 min, this drug being washed out shortly after the contraction had peaked. In veins from unoperated rats, this concentration of ATP produces approximately 80% of the maximum contraction to this agent.
Following these chemical stimulation protocols, contractions to 60 mM K+ were recorded with prazosin (0.1 μM) and rauwolscine (1 μM) present to prevent the excitatory actions of NE released from the sympathetic nerves by depolarization.
Histological assessment of lumen diameter
Tail veins were dissected from three 8-week SCI rats and three unoperated rats that had been perfusion-fixed. These animals were deeply anesthetized with pentobarbitone (100 mg/kg i.p.) and perfused through the descending thoracic aorta at 100 mmHg with normal saline containing heparin (10,000 IU/L) and sodium nitrite (10 mg L−1) followed by Zamboni's fixative. Tissues were dissected and post-fixed overnight before washing in phosphate-buffered saline (PBS) and infiltrating with 30% sucrose in PBS. The tissues were then embedded in Optimal Cutting Temperature compound (Tissue Tek, Elkhart, IN) and frozen until used (-80°C). Transverse sections of the vessels (20 μm thick) were cut on a cryostat, mounted on gelatinized slides, and stained with hematoxylin and eosin. Digital images were taken using a Spot camera (Diagnostic Instruments, Sterling Heights, MI) on an Olympus microscope, and the lumen circumference and vessel wall thickness were measured using ImageJ software (
Data analysis
In this study, groups of vein segments from rats with spinal cord transection are referred to as “SCI veins” and those from sham-operated rats are referred to as “control veins.” The peak amplitudes of the contractions to electrical and chemical stimulation were measured. Because the amplitude of the nerve-evoked contractions in untreated veins declined with time during the experiment (<15%), the % blockades produced by the antagonists were calculated by comparing the relative changes produced in the drug-treated veins with those observed at the same time points in untreated veins. For contractions evoked by 10 stimuli at 1 and 10 Hz, the rise time was the interval between 10 and 90% of the peak contraction and the 50% decay time was the interval between the peak and 50% of the peak contraction during the decay phase. The 50% decay time of contraction to 60 mM K+ was measured from the time this stimulus was removed until the amplitude of the contraction had fallen to 50% of its value at the start of washout. The half maximal effective concentration (EC50) for each of the α-adrenoceptor agonists was estimated by fitting their concentration-response curves to the Hill equation using Igor Pro (Wavemetrics, Lake Oswego, OR).
All statistical comparisons were made using SPSS 17 (SPSS Inc., Chicago, IL). Prior to comparison, the data sets were checked for homogeneity of variance using Levene's test. Where the variance did not differ between the data sets, the data are presented as mean and standard error of the mean (SEM) and comparisons are made with Student paired t tests (within group comparisons) or unpaired t tests (between group comparisons). Where the variance differed significantly between the data sets (p < 0.05), the data are presented as median and interquartile range (IQR) and comparisons are made with Mann-Whitney U tests (between group comparisons). P values <0.05 were considered to indicate a significant difference. When multiple comparisons were made, the false discovery rate procedure was used to adjust the significance level (Curran-Everett, 2000). In all cases, the value n is the number of animals studied.
Results
Basal conditions
Table 1 shows the lumen diameter and basal ETP for control and SCI tail veins (n = 6 for both groups). Whereas the lumen diameter of SCI veins was ∼30% smaller than control, the basal ETP did not differ significantly between the two groups. The effect of spinal cord transection on lumen diameter of tail veins was confirmed in transverse sections of these vessels obtained from perfusion-fixed 8-week SCI and age-matched unoperated control rats (control, 705 ± 13 μm, n = 3; SCI, 474 ± 21 μm, n = 3; p < 0.01). The lumen diameter of both sets of fixed veins was smaller (∼25%) than that estimated for veins mounted in the wire myograph (Table 1), which can be explained by tissue shrinkage during processing and the possibility that the transmural pressure at the time of fixation was less than 2 kPa (15 mmHg). Nevertheless, as for the myograph mounted vessels, lumen diameter after SCI was ∼30% smaller than control. Vessel wall thickness measured in transverse sections did not differ between control (14.7 ± 1.5 μm) and SCI (14.0 ± 0.8 μm) veins (p = 0.78).
Lumen diameter is presented as median and interquartile range (in parentheses), and effective transmural pressure
Statistical difference between lumen diameter of control and SCI veins by Mann Whitney U test, p < 0.01.
Nerve-evoked increases in ETP were potentiated after SCI
The reduction in lumen diameter will make the transmural pressure that the SCI veins generate larger for a given increase in wall tension. To illustrate this effect, the contractile responses to trains of 25 stimuli at 0.1 to 1 Hz are presented both as increases in ETP (Fig. 2A,B) and as increases in wall tension (Fig. 2C). From a functional point of view, changes in transmural pressure, which determine the lumen cross-sectional area of peripheral veins and therefore their volume (Rowell, 1977), are more important than increases in wall tension and, because of the changes in vessel diameter after SCI, all contractions described below are presented as increases in ETP. The increases in ETP produced by these trains of stimuli were about twice as large in the SCI veins as in control veins at all frequencies of stimulation studied.
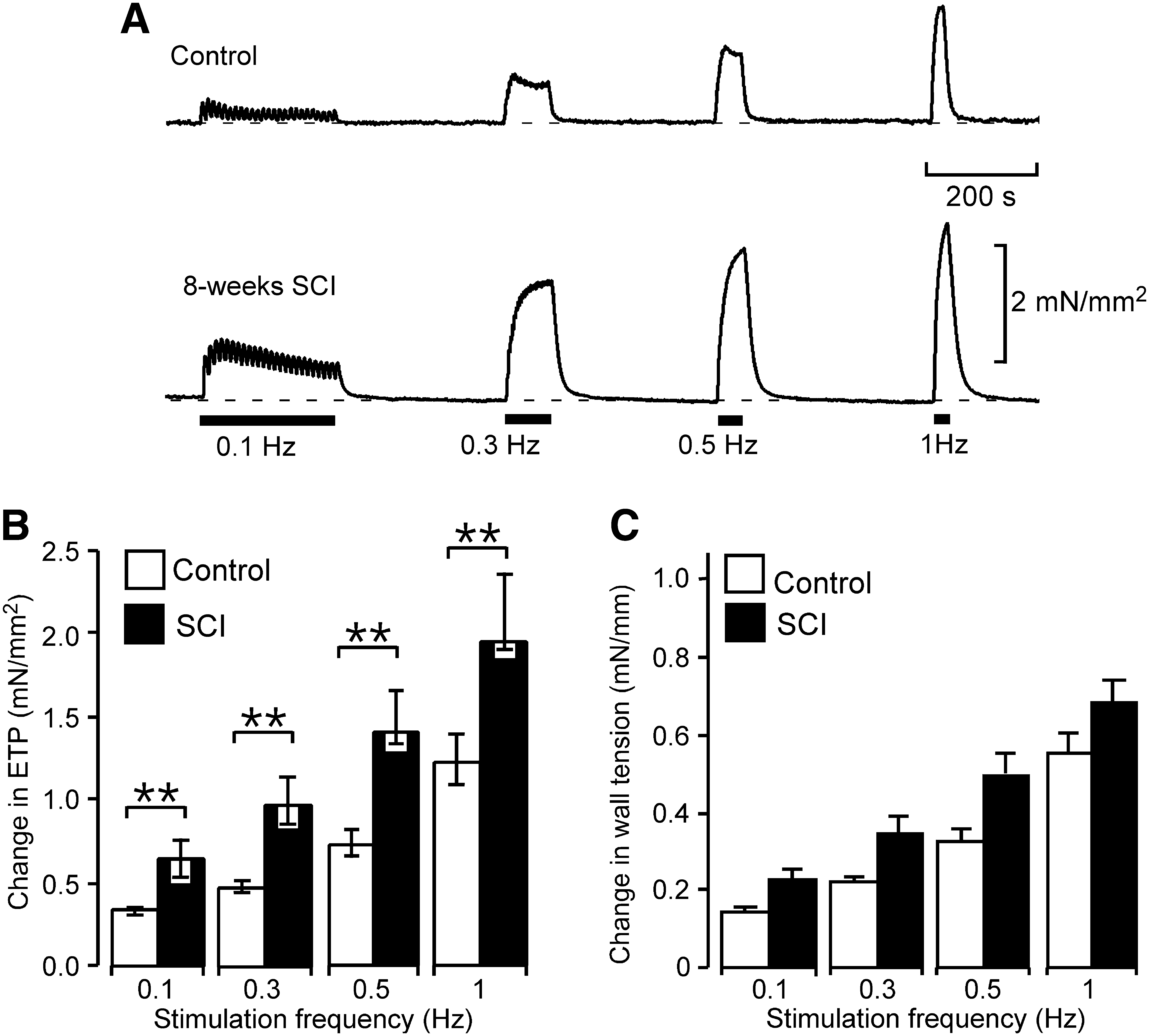
Contractions to perivascular nerve stimulation in tail veins were enhanced after a T4 spinal cord transection. (
Spinal cord transection did not change time course of nerve-evoked contractions
In tail arteries, SCI not only increased the amplitude of nerve-evoked contractions evoked by short trains of stimuli but also prolonged their time course (Yeoh et al., 2004a). In SCI veins, the increases in ETP produced by trains of 10 stimuli at 1 and 10 Hz were larger than those in controls but the 10 − 90% rise time and the 50% decay time of contractions did not differ from that in controls (Table 2).
Changes in effective transmural pressure (ETP) are presented as median and interquartile range (in parentheses), and rise and decay times as mean and SEM.
p < 0.05, **p < 0.01––statistical differences between control and SCI veins by Mann Whitney U-test.
Contribution of α2-adrenoceptors to nerve-evoked contractions was reduced after SCI
Because the effects of blocking α-adrenoceptors and/or P2-purinoceptors on neurally evoked contractions were similar for the different patterns of stimulation, only the results for contractions to trains of 100 stimuli at 1 Hz are presented. The increases in ETP produced by these trains of stimuli were larger in SCI veins than in their controls (control, 1.28 mN/mm2 [IQR 1.07 − 1.39]; SCI, 2.27 mN/mm2 [IQR 2.01 − 2.77]; Mann Whitney U test, p < 0.01). Figure 3A − C shows the percentage of the nerve-evoked contractions remaining in the presence of the antagonists. For prazosin (10 nM), this did not differ between SCI and control veins (p = 0.49; Fig. 3A). However, the residual contraction in the presence of rauwolscine (0.1 μM) was greater than control in SCI veins (Fig. 3B). In combination, prazosin (0.1 μM) and rauwolscine (1 μM) reduced the amplitude of nerve-evoked contractions by approximately ∼75% in both groups of vessels (Fig. 3C; control vs. SCI, p = 0.86). These higher doses of prazosin (0.1 μM) and rauwolscine (1 μM) individually produced a slightly bigger reduction in ETP (results not shown) but these effects, in each case, could reflect antagonism of the other α-adrenoceptor subtype (Bylund et al., 1992; Timmermans et al., 1984).

The percentage of the nerve-evoked contraction sensitive to blockade of α2-adrenoceptors was reduced in tail veins after SCI. The contractions in the presence of prazosin (
In both control and SCI veins, blockade of P2-purinoceptors with suramin (0.1 mM) did not change the nerve-evoked increases in ETP (paired t test, p = 0.52 and 0.50, respectively). However, in both groups of veins, the remaining nerve-evoked contraction in the presence of antagonists to both α-adrenoceptors (with 0.1 μM prazosin and 1 μM rauwolscine) and P2-purinoceptors (with 0.1 mM suramin) was smaller than that in the presence of the antagonists of α-adrenoceptors alone (see Fig. 3C,D; paired t test, p < 0.05 for control veins and p < 0.01 for SCI veins). There was no difference in the remaining contraction (∼10%) produced by this combination of antagonists between control and SCI veins (p = 0.73). As TTX (0.5 μM) abolished all electrically evoked contractions (see Methods section), the small residual contraction remaining when both α-adrenoceptors and P2-purinoceptors were blocked may be due to nerve-released neuropeptide Y, which is a potent contractile agent in tail veins when applied exogenously (Tsurumaki et al., 2002).
Spinal cord transection did not increase sensitivity to α-adrenoceptor agonists or change NET activity
Table 3 shows the EC50 values for PE, methoxamine, and clonidine in control and SCI veins. Sensitivity to PE was assessed in both the absence and presence of the NET inhibitor DMI (30 nM). The EC50 for methoxamine was slightly greater in SCI veins compared to that in control veins but the EC50 values for both PE and clonidine did not differ between these groups of vessels. Furthermore, DMI produced a similar decrease (2- to 3-fold) in the EC50 for PE in both control and SCI veins (p = 0.16), indicating that there was no change in the activity of NET produced by SCI.
Sensitivity to phenylephrine was assessed in both the absence and presence of the NET inhibitor desemthylimipramine (DMI; 30 nM). EC50 values for clonidine are presented as median and interquartile range (in parentheses). All other data are presented as mean and SEM.
p < 0.05 - Statistical difference between control and SCI veins by unpaired t test.
There were no significant differences in the maximum increase in ETP produced by the α-adrenoceptor agonists between control and SCI veins (Fig. 4A – C).

Maximum increases in effective transmural pressure (ETP) produced by both α1- and α2-adrenoceptor agonists were unchanged in tail veins after SCI. Maximum increases in ETP produced by phenylephrine (
Increase in ETP produced by ATP did not change after SCI
The increase in ETP produced by ATP (0.1 mM) did not differ between control and SCI veins (Fig. 5A).
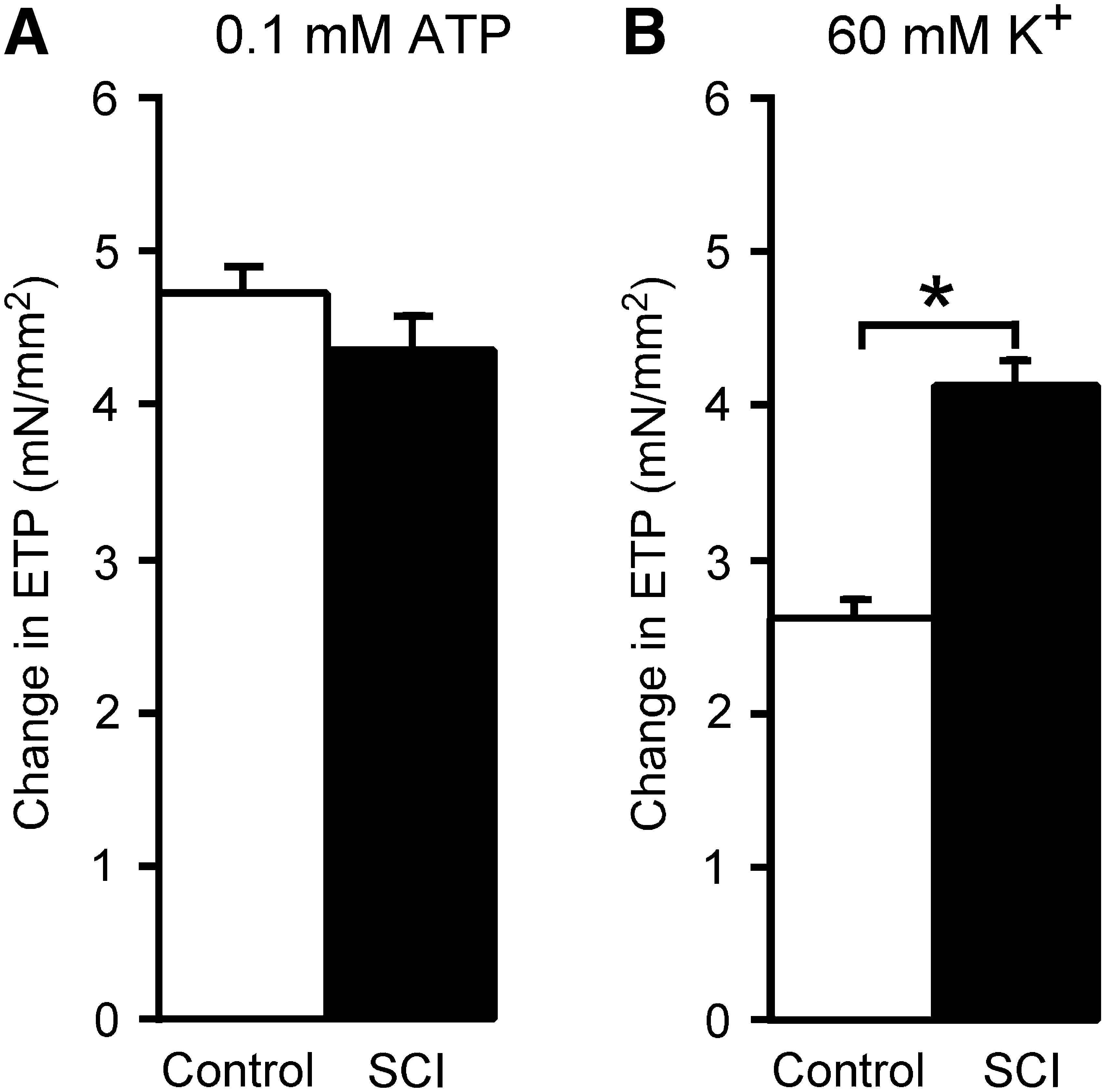
Increases in effective transmural pressure (ETP) produced by ATP were unchanged but those produced by 60 mM K+ were augmented in SCI veins. Increases in ETP by 0.1 mM ATP (
Increase in ETP produced by raised [K+] was greater but not more prolonged after SCI
The increases in ETP produced by raising the K+ concentration to 60 mM were greater in SCI veins than in control veins (Fig. 5B). In addition, there was a positive correlation between the response of veins to 60 mM K+ and their responses to all the patterns of nerve stimulation used (Spearman's rank coefficient ρ > 0.8, p < 0.01 for all comparisons).
The time to 50% decay of high K+-evoked contractions following washout was prolonged in tail arteries from rats after SCI relative to controls (Yeoh et al., 2004a). In SCI veins, the 50% decay time (24.4 ± 1.8 sec) did not differ significantly from that in controls (28.1 ± 3.9 sec, p = 0.22).
Discussion
This study demonstrates that chronic spinal cord transection potentiates nerve-evoked increases in ETP in rat lateral tail veins. This enhancement of reactivity was associated with a 30% reduction in lumen diameter and a lesser proportion of nerve-evoked contraction that was blocked by the α2-adrenoceptor antagonist rauwolscine. There was no increase in sensitivity of the venous smooth muscle to α1- or α2-adrenoceptor agonists or in the effectiveness of the NET. Our findings also indicate that responsiveness of the venous smooth muscle to ATP was not changed by SCI. The potentiation of nerve-evoked contractions seems to involve changes in post-receptor mechanisms in the smooth muscle and probably in NE release.
In individuals with long-standing SCI, both venous capacity and compliance are reduced in the lower limbs (Hopman et al., 1994; Wecht et al., 2000). These findings have been suggested to result from adaptations of the venous vasculature to inactivity of skeletal muscles. Accordingly, the diameter of the popliteal vein measured at the same distending pressure was ∼45% smaller in a group of subjects with motor complete SCI (ASIA A) than that in able-bodied subjects (de Groot et al., 2005). As this change could not be attributed to nerve-mediated venoconstriction, the change in diameter was most likely due to structural remodeling of the vein. The lumen diameter of the common femoral artery is reduced by about ∼30% in humans with SCI in comparison with that of able-bodied humans, a change that occurs rapidly after injury (de Groot et al., 2006; Olive et al., 2003). In rats with spinal cord transections, we have observed reductions in the lumen diameter of tail arteries (∼15%) (Yeoh et al., 2004a) and saphenous arteries (∼20%) (Rummery et al., 2010), suggesting that vascular remodeling may be widespread following SCI. Remodeling of arterial vessels may be an adaptation to the reduction in peak levels of vessel wall stress and endothelial shear stress that occurs after SCI (de Groot et al., 2006). A chronic reduction in blood flow through mesenteric arteries has been demonstrated to produce both a decrease lumen diameter and thinning of the vessel wall (Pourageaud and De Mey, 1997). In this case, the change in blood flow initially reduces both circumferential wall stress and endothelial sheer stress, with remodeling restoring wall stress, and probably shear stress, to normal values. It is possible that the smaller lumen diameter of the SCI tail veins is also an adaptation to a reduction in blood flow, although in the present study we did not detect a change in wall thickness.
Because of the smaller lumen diameter of SCI veins, the change in ETP produced by a given increase in wall tension is enhanced. Importantly, the maximum increases in ETP to α-adrenoceptor agonists did not differ significantly between the control and SCI veins. This implies that the contractile capacity of the venous smooth muscle scales with the difference in vascular dimensions after SCI. Furthermore, this finding indicates that the greater magnitude of the nerve-evoked increases in ETP in SCI veins must involve enhancement in neurovascular transmission.
Our data show that in control tail veins nerve-evoked contractions are due to NE acting at both α1- and α2-adrenoceptors on the vascular smooth muscle. When both these receptor subtypes were blocked by the combination of prazosin and rauwolscine, reducing the response by 75%, ATP acting at P2-purinoceptors contributed to the remaining contractions. Furthermore, the proportional blockade produced by rauwolscine alone (∼70%) was only slightly less than that produced by the combined blockade, whereas prazosin alone reduced the contractions of control tail veins by nearly 50%. These non-additive effects of the antagonists suggest synergistic interactions between the effects of activating α1- and α2-adrenoceptors in tail veins, similar to those we have previously reported in the control tail artery (Yeoh et al., 2004a).
Our findings suggest that SCI reduces the synergistic interactions between the effects of activating α1- and α2-adrenoceptors. In SCI veins, the proportional blockade produced by the combination of prazosin and rauwolscine (73%) was close to the sum of the blockades produced individually by prazosin (38%) and rauwolscine (45%). In tail arteries, we have observed a similar reduction in the component of contraction that is mediated by co-activation of α1- and α2-adrenoceptors after SCI (Yeoh et al., 2004a).
The degree of blockade produced by the combination of α1- and α2-adrenoceptor antagonists (∼75%) did not differ between control and SCI veins, indicating that the relative contribution of NE to neural activation of the vascular smooth muscle was not changed after SCI. Furthermore, as antagonism of both α-adrenoceptors and P2-purinoceptors also produced similar degrees of blockade (∼90%) in SCI and control veins, the relative contribution of nerve-released ATP was also unaffected by SCI.
Activation of α2-adrenoceptors in the tail artery augments contractions to α1-adrenoceptor agonists through a mechanism that involves an increase in Ca2+ influx into smooth muscle cells through voltage-gated Ca2+ channels (Xiao and Rand, 1989). As SCI increases the contractions to depolarization with K+ in both tail veins and tail arteries (Yeoh et al., 2004a), perhaps depolarization-induced Ca2+ entry increases after SCI. Alternatively SCI may selectively sensitize the contractile mechanism to Ca2+ entering from outside the cell. Importantly, in both tail veins and tail arteries (Yeoh et al., 2004a), the contractions to 60 mM K+ were positively correlated with those to nerve stimulation, suggesting that the mechanism involved in increasing the responses of SCI vessels to depolarization with K+ also contributes to the augmentation of nerve-evoked contractions.
The changes that produce the larger nerve-evoked increase in ETP in SCI veins must involve changes in the post-receptor mechanisms in the venous smooth muscle. In the control vein, most of the contraction arises from co-activation of α1- and α2-adrenoceptors. Because a smaller percentage of the enlarged nerve-evoked response after SCI was blocked by the α2-adrenoceptor antagonist and the component due to co-activation of α1- and α2-adrenoceptors is greatly reduced, a large component of the response evoked after SCI must be generated by activation solely of α1-adrenoceptors, presumably linked to the activation of contraction by a different pathway that does not involve the α2-adrenoceptors. The details of such pathways remain to be identified.
Other factors may reduce the proportional blockade of nerve-evoked venous contraction by rauwolscine after SCI. Blockade of pre-junctional α2-adrenoceptors by this agent might be expected to increase NE release (Starke et al., 1989). We cannot exclude the possibility that the facilitatory effect of blocking pre-junctional α2-adrenoceptors on NE release is increased after SCI and that this effect contributes to the apparent reduction in the blockade of nerve-evoked contraction produced by rauwolscine.
The magnitude of the SCI-induced increase in nerve-evoked responses differs markedly between tail arteries and tail veins. In tail arteries from rats with SCI, the contractions to stimulation of tail arteries at 0.1 Hz were magnified > 25-fold but those to stimulation at 1 Hz only 3-fold. In contrast, the enhancement of nerve-evoked responses in SCI veins was much smaller and little affected by the frequency of nerve stimulation, with the increase in ETP evoked by stimulation at either 0.1 or 1 Hz being increased only ∼2-fold. The other major difference was that the tail arteries from rats with SCI had a reduced EC50 for clonidine, whereas the EC50 for clonidine in veins was unchanged by SCI. Furthermore, in rat tail arteries, the EC50 for clonidine was negatively correlated with the size of contractions evoked by nerve stimulation (Yeoh et al., 2004a) whereas in tail veins these measures were not correlated (Spearman's rank coefficient, p = 0.51). Thus the large effect of SCI on α2-adrenoceptors in tail artery does not occur in tail vein.
Pre-junctional changes in neurotransmitter release probably also contribute to the potentiation of neurovascular transmission after SCI. In tail arteries from rats with SCI, the amplitude of purinergic excitatory junction potentials was increased by ∼100% compared to that in control arteries, suggesting an increase in neurotransmitter release (McLachlan and Brock, 2006). In rabbit ear arteries that had been chronically decentralized by cutting the pre-ganglionic inputs to the post-ganglionic neurons supplying these vessels, the release of NE from the perivascular axons was enhanced (Tsuru et al., 1993), suggesting that decreases in ongoing nerve activity increase neurotransmitter release from sympathetic nerve terminals. Neurotransmitter release is also facilitated after decreases in ongoing nerve activity at other peripheral and central neuronal synapses (Gallego and Geijo, 1987; Murthy et al., 2001). However, in both tail arteries and veins, we have not managed reliably to quantify NE released by the short trains of stimuli used here to produce nerve-evoked contractions.
Conclusion
Severing the bulbospinal connections to the pre-ganglionic vasoconstrictor neurons supplying the tail vein leads to potentiated nerve-evoked contractions of this vessel, as reported for arteries below a spinal cord transection. From a functional point of view, the reduction in lumen diameter of the SCI veins is the most important change, as this structural adaptation will increase the pressure that the smooth muscle exerts on the luminal contents when the veins are neurally activated. In the tail vein, the enhanced neurovascular transmission cannot be explained by a general hypersensitivity of the vascular smooth muscle to neurotransmitters. Rather, the smooth muscle function is modified, possibly by an increased contribution of Ca2+ influx to contraction. Assuming that similar changes in vascular dimensions and neurovascular transmission occur in other cutaneous veins below the level of a spinal lesion, this would explain how they contribute to the maintenance of venous return during autonomic dysreflexia. It is worth stressing that, while neural control of venous blood flow in skin is not normally considered important in the regulation of blood pressure in able-bodied subjects (Rowell, 1977), during autonomic dysreflexia the venoconstriction is widespread (Corbett et al., 1971; Mathias and Frankel, 1999), increasing venous return and almost certainly contributing to the generation of very high levels of blood pressure that can threaten the lives of spinal cord-injured people.
Footnotes
Acknowledgments
This study was supported by a program grant from the New South Wales Spinal Cord Injury and Related Neurological Conditions Research Grants Program. J.B. is supported by a fellowship grant (no. 350904) from the National Health and Medical Research Council of Australia.
Author Disclosure Statement
No competing financial interests exist.