
Abstract
Traumatic brain injury (TBI) produces several cellular changes, such as gliosis, axonal and dendritic plasticity, and inhibition-excitation imbalance, as well as cell death, which can initiate epileptogenesis. It has been demonstrated that dysfunction of the inhibitory components of the cerebral cortex after injury may cause status epilepticus in experimental models; we proposed to analyze the response of cortical interneurons and astrocytes after TBI in humans. Twelve contusion samples were evaluated, identifying the expression of glial fibrillary acidic protein (GFAP) and calcium-binding proteins (CaBPs). The study was made in sectors with and without preserved cytoarchitecture evaluated with NeuN immunoreactivity (IR). In sectors with total loss of NeuN-IR the results showed a remarkable loss of CaBP-IR both in neuropil and somata. In sectors with conserved cytoarchitecture less drastic changes in CaBP-IR were detected. These changes include a decrease in the amount of parvalbumin (PV-IR) neurons in layer II, an increase of calbindin (CB-IR) neurons in layers III and V, and an increase in calretinin (CR-IR) neurons in layer II. We also observed glial fibrillary acidic protein immunoreactivity (GFAP-IR) in the white matter, in the gray-white matter transition, and around the sectors with NeuN-IR total loss. These findings may reflect dynamic activity as a consequence of the lesion that is associated with changes in the excitatory circuits of neighboring hyperactivated glutamatergic neurons, possibly due to the primary impact, or secondary events such as hypoxia-ischemia. Temporal evolution of these changes may be the substrate linking severe cortical contusion and the resulting epileptogenic activity observed in some patients.
Introduction
A
Although the mechanisms involved in the generation of epilepsy after TBI are not clearly understood, the damage to superficial layers of the cerebral cortex could be a common factor in several forms of trauma that involves subdural or epidural hematoma and/or depressed skull fracture (Frey, 2003; D'Ambrosio and Perucca, 2004). Pitkänen and associates (2007) indicated that epilepsy after TBI may develop in three phases: lesion, epileptogenesis, and spontaneous recurrent seizure generation. During the epileptogenesis step several cellular changes occur, such as gliosis, neurogenesis, axonal and dendritic plasticity, matrix rearrangement, angiogenesis, and/or cell death. These cell alterations may be accompanied by physiological and biochemical alterations in the cerebral cortex that produce, among other changes, a decrease in the frequency of inhibitory currents and in the synaptic release of GABA (D'Ambrosio and Perucca, 2004).
Chu and colleagues (2003), Salin and Prince (1996), and Yang and Benardo (1997, 1998) all reported that a decrease in inhibition generated by an acute cortical lesion is primarily due to loss of inhibitory neurons on the superficial layers. In their model of acute cortical lesion Yang and associates (2007) propose that after a superficial cortical lesion, dendritic processes coming up from infragranular layers are sectioned, and consequently an alteration of inputs to layer I are generated. They suggest that layers IV and V, which are supposedly controlling net synchronized activity, are thus deprived of the inhibitory inputs originated in the superficial layers.
Since inhibitory control of the cerebral cortex depends upon interneurons and management of the microenvironment by astrocytes, and physiologically it has been demonstrated that their dysfunction after cortical lesion generates an epileptic state, we proposed to analyze the response of cortical interneurons and astrocytes after TBI in humans. For this purpose we utilized antibodies against calcium-binding proteins (CaBPs): calbindin (CB), parvalbumin (PV), and calretinin (CR), which specifically stain the GABAergic subpopulations of interneurons in the cerebral cortex, and glial fibrillary acidic protein (GFAP), to study the astrocytic reaction to the lesion.
Methods
All the experiments described here were approved by the Ethical Committee of the Universidad del Valle, Cali, Colombia (acta no. 045, 2004). Control tissue was obtained from 12 subjects from the Instituto de Medicina Legal and Ciencias Forenses, Cali, with a mean post-mortem interval of 10 h, the time period recommended for this type of procedure that enabled us to discriminate the complete cytoarchitectural organization of the cerebral cortex. These subjects did not have previous reports of neurological or psychiatric disease, trauma, edema, or any other alteration, according to the forensic criteria (Table 1). Traumatic tissue was obtained from 11 patients subjected to decompressive hemicraniectomy with evacuation of the hemorrhagic contusion of the brain and dural graft. The tissue samples corresponded to the contusion area that the surgeon usually removes in these procedures (Table 2). No additional tissue was removed for this study. Patient data were obtained from the surgical clinical reports at the Hospital Universitario del Valle. All samples were washed for 10 min in saline solution (10%), and fixed in paraformaldehyde-lysin-sodium periodate (pH 7.4) for 6–10 days at 4°C.
PMI, post-mortem interval (hours); M, male; LPC, lateral prefrontal cortex; LTC, lateral temporal cortex; HG, homicide by gunshot; HS, homicide by stabbing; RA, ruptured aortic aneurysm.
HPPF, hypoperfusion; HPX, hypoxia; GCS-A, Glasgow Coma Scale score at emergency room admission; GCS-B, Glasgow Coma Scale score at surgery; DIS, days between injury and surgery; GOS, Glasgow Outcome Scale score; M, male; F, female.
Immunohistochemical procedure
All samples were coronally cut into 50-μm sections with a vibratome. In each case the first section was used for rapid staining with toluidine blue in order to verify the appropriate plane of section. For each sample, consecutive sections were incubated for 18 h at room temperature with the following antibodies: NeuN (mouse anti-neuronal nuclei monoclonal antibody MAB377 diluted 1:2500; Chemicon International,, Temecula, CA) to identify the whole neuronal population, excluding the glial cells; parvalbumin (monoclonal anti-parvalbumin clone PARV-19 P-3088 diluted 1:5000; Sigma, St. Louis, MO); calbindin (monoclonal anti-calbindin D-28K clone CB-955 G9848 diluted 1:5000; Sigma), and calretinin (polyclonal anti-calretinin clone AB-149 diluted 1:5000; Chemicon International), to selectively identify the GABAergic populations, and GFAP (monoclonal anti-GFAP clone G-A-5; Sigma), to identify the intermediate filaments of the astrocyte population. After incubation with primary antibodies, the sections were washed with PBS and processed with the avidin-biotin HRP complex (Vectastain Elite ABC; Vector Laboratories, Southfield, MI). Control sections were processed in the same way except that the primary antibody was omitted.
Sections were photographed, at 10× magnification with a Canon Power Shot A85 (Canon, Tokyo, Japan) adapted to an Olympus CH-2 optical microscope model CHS (Olympus, Tokyo, Japan). Images from each cortical lamina were fused using the Canon Photo Stitch program version 3.1 2000. Fusion of consecutive cortical images from pial to white matter led to obtaining the whole cortical column. Microphotographs were calibrated with a 1-mm stage objective micrometer (type A, MBM11100l; Nikon, Tokyo, Japan), with 100 subdivisions of 10 μm each. Boundaries between each cortical lamina were established according to the densities and cell forms characteristic of each layer (Barbas and Pandya, 1989; Gittins and Harrison, 2004), and the independent criteria of three observers. For high-resolution images a BX-60 Olympus microscope coupled to a 750L Cool CCD camera (Optronics, Goleta, CA) and a PC running Image Pro Plus (Media Cybernetics, Silver Spring, MD) were used. Multiple focal planes were added to create mosaics of the desired microscopic field, and the function “Enhanced Depth of Field” of the software was used to yield the final images.
Zones with laminar and zones without laminar preservation (patches) in posttraumatic tissue
Previous studies using the neuronal markers NeuN and MAP2 led us to establish the cell laminar distribution in normal human cerebral cortex (Buriticá and Pimienta, 2007). In injured tissue we described two zones: one in which the laminar organization could be observed, and another without laminar preservation, defined as patch areas (Fig. 1). Once we identified the areas to be evaluated we proceeded to measure cortical laminas and cell counts for each type of antibody on 10 × microphotographs using the program SigmaScan Pro 5.0 (SPSS Inc., Chicago, IL). The densities were calculated by counting the number of positive cells for each type of antibody, using boxes 300 μm wide and 100 μm deep located in the center of each lamina. In sections in which the laminar organization was not preserved, the grid was placed in the center of the lamina according to the laminar measures obtained in the consecutive NeuN-immunostained sections (Buriticá and Pimienta, 2007).
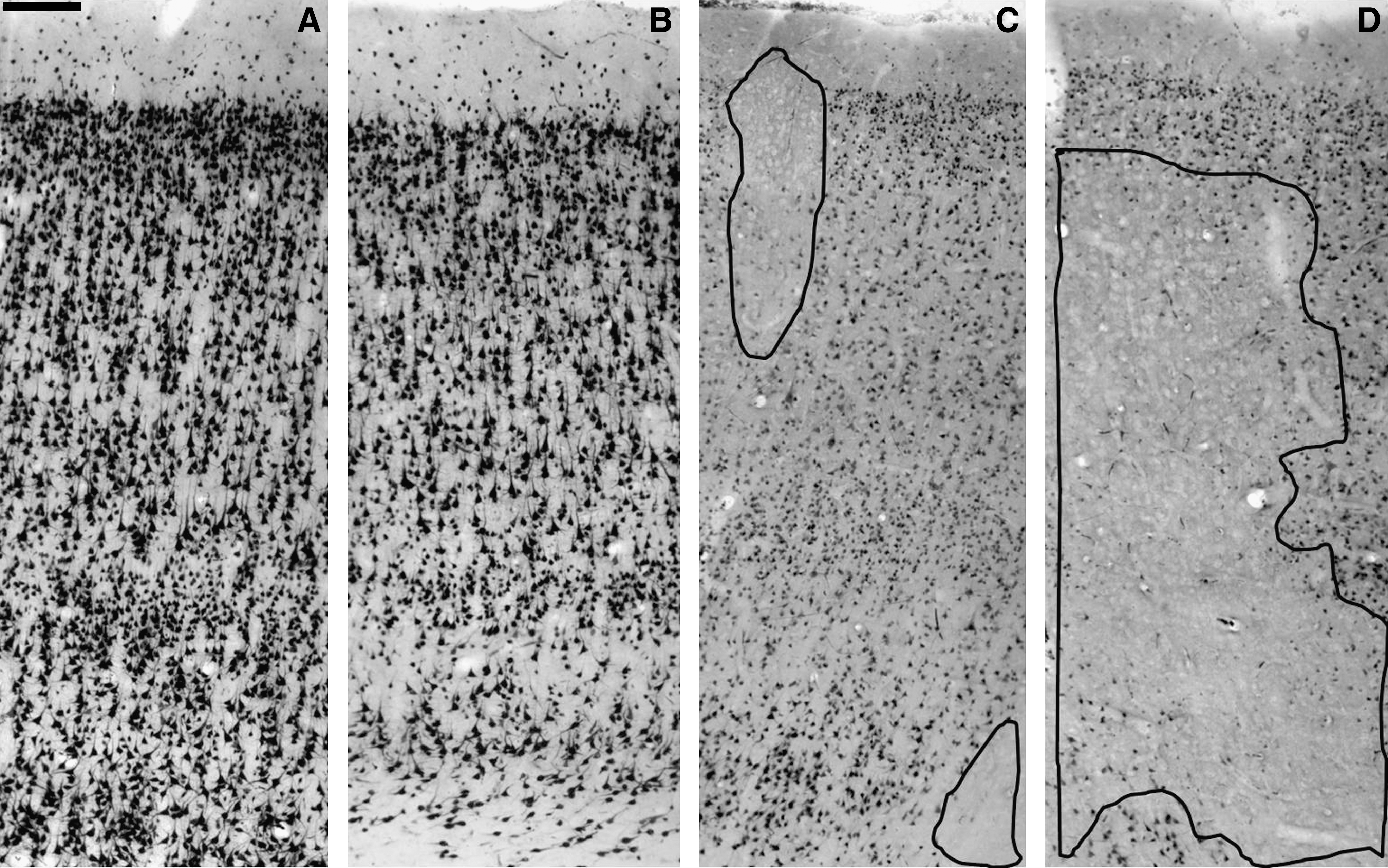
Variations in NeuN expression after human TBI. (
Quantitative analysis
The counts of PV-, CB-, and CR-positive neurons in each cortical layer are expressed as mean and standard deviation comparing control and trauma samples. In order to establish whether there were differences in the homologous cortical layers between control and trauma samples, we compared the means of the counts in each cortical layer using a one-tailed non-paired Student's t-test with statistical significance set at p < 0.05.
Results
Cortical layers in control and trauma samples were identified using NeuN antibody, according to previously established criteria (Medina and Escobar, 2002; Buriticá and Pimienta, 2007; Fajardo et al., 2008). Control samples led us to identify the entire neuronal population with well-defined cell bodies, dark nuclei, and less stained cytoplasm (Fig. 2); all cortical layers could be observed (Fig. 1A). In TBI samples, those areas with laminar conservation (Fig. 1B) showed a uniform background with NeuN-positive cells visible, but with less stained nuclei and darker cytoplasm. The borders appear interrupted and processes were not visible all the way across. These neurons were homogeneously distributed, with a slight tendency to appear more often in supragranular layers (Fig. 2). Areas without laminar conservation, or patch areas, showed loss of NeuN-IR, particularly in the transition between layers II–III and areas with decreased IR (Fig. 1C–D and Fig. 2).
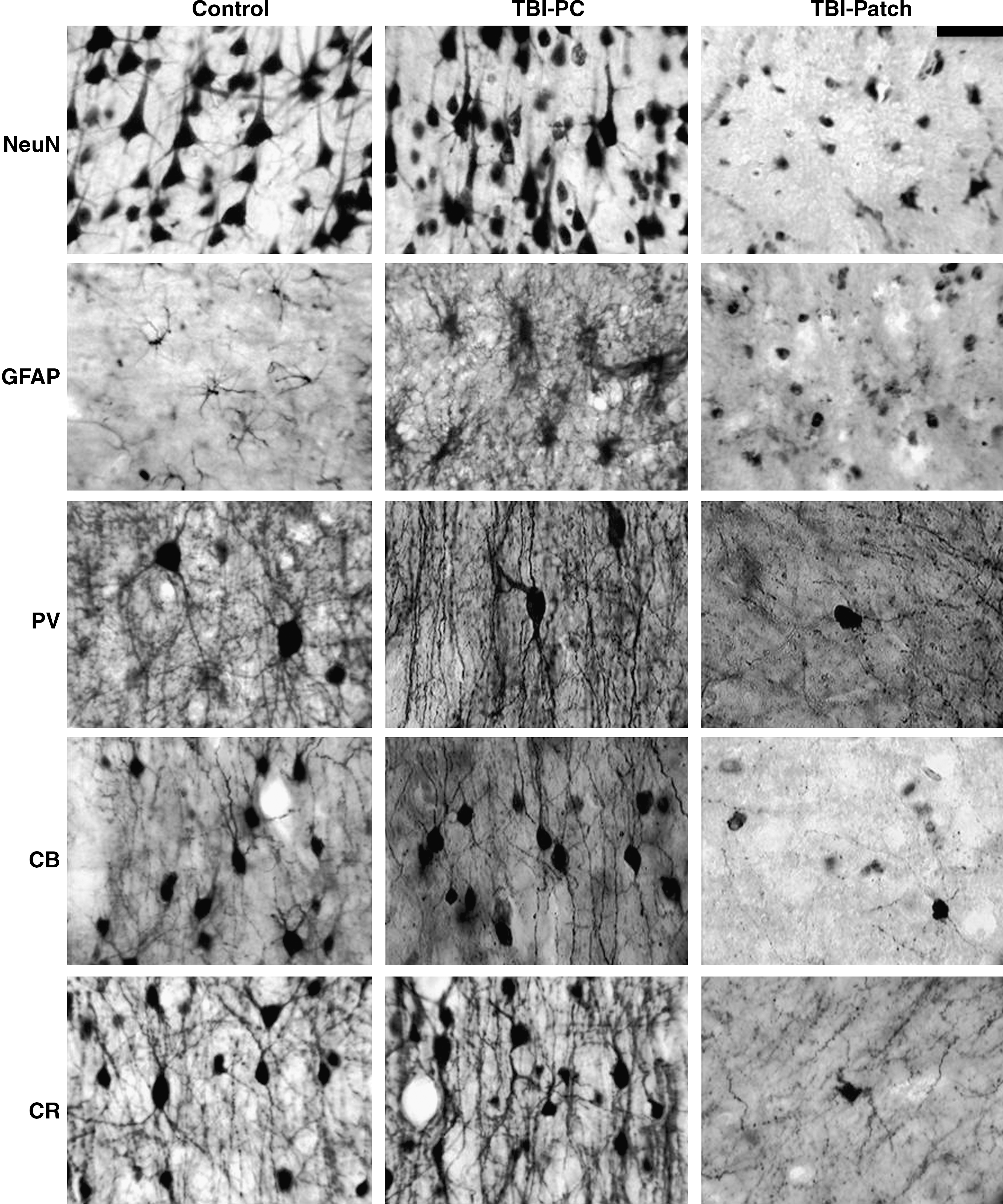
Changes in NeuN, GFAP, PV, CB, and CR expression after human TBI. Images in the left column correspond to post-mortem tissue with no brain injury, the middle column corresponds to TBI tissue sectors with preserved cytoarchitecture (TBI-PC), and the right column corresponds to TBI tissue sectors with patches of loss of immunoreactivity (TBI-Patch) (scale bar = 50 μm).
Immunoreactivity for GFAP, PV, CB, and CR were also analyzed qualitatively in both zones. Quantitative analysis for CaBPs was made only in areas with laminar conservation. The laminar distribution of CaBPs seen in control samples was similar to that described by others (Marco et al., 1996; DeFelipe, 1997, 1999; DeFelipe et al., 1999; Tooney and Chahl, 2004).
GFAP expression
In control samples few GFAP-IR cells were observed (Fig. 2). Occasionally some cell bodies showed abundant thin processes commonly associated with blood vessels, or in the layer VI–white matter transition. On layer I multiple astrocyte varicose processes were observed.
In general, TBI samples showed an increase in GFAP-IR, and cell bodies were bigger with more vigorous processes (Fig. 2). Layer I and white matter showed the densest IR processes, with some hypertrophic bodies in the latter. Some of these processes were bifurcated and perpendicular to the blood vessels or to the gyrus crest.
Those sectors corresponding to laminar preservation had thicker hyperreactive processes in layer I with no apparent varicosities as seen in control tissue (Fig. 2). Additionally, we observed an increase in hypertrophic astrocytes sometimes associated with blood vessels, especially surrounding patch areas and in the layer VI–white matter transition.
Sectors around microhemorrhages (coincident with NeuN patch areas) showed cavitations with some pale areas devoid of cell somas or processes, and other neighboring sectors showed small GFAP-IR astrocytes without processes and some hypertrophic cells around blood vessels (Fig. 2). Cell processes in layer I were also decreased. When patches were located in infragranular layers, hypertrophic astrocytes appeared to be located under sectors devoid of patches.
Parvalbumin expression
In control samples PV-IR was present from layers II to VI (Fig. 3). Neuronal morphology included multipolar forms with ovoid somata, and 3–4 processes radially distributed. Some large multipolar cells were also observed. According to Kawaguchi and Kubota (1997), DeFelipe (1997), and Gonzalez-Albo and associates (2001), these two cell types correspond to basket and chandelier cells. In layers V and VI, PV-IR cells were scant, with predominantly bipolar forms with their processes oriented vertically.
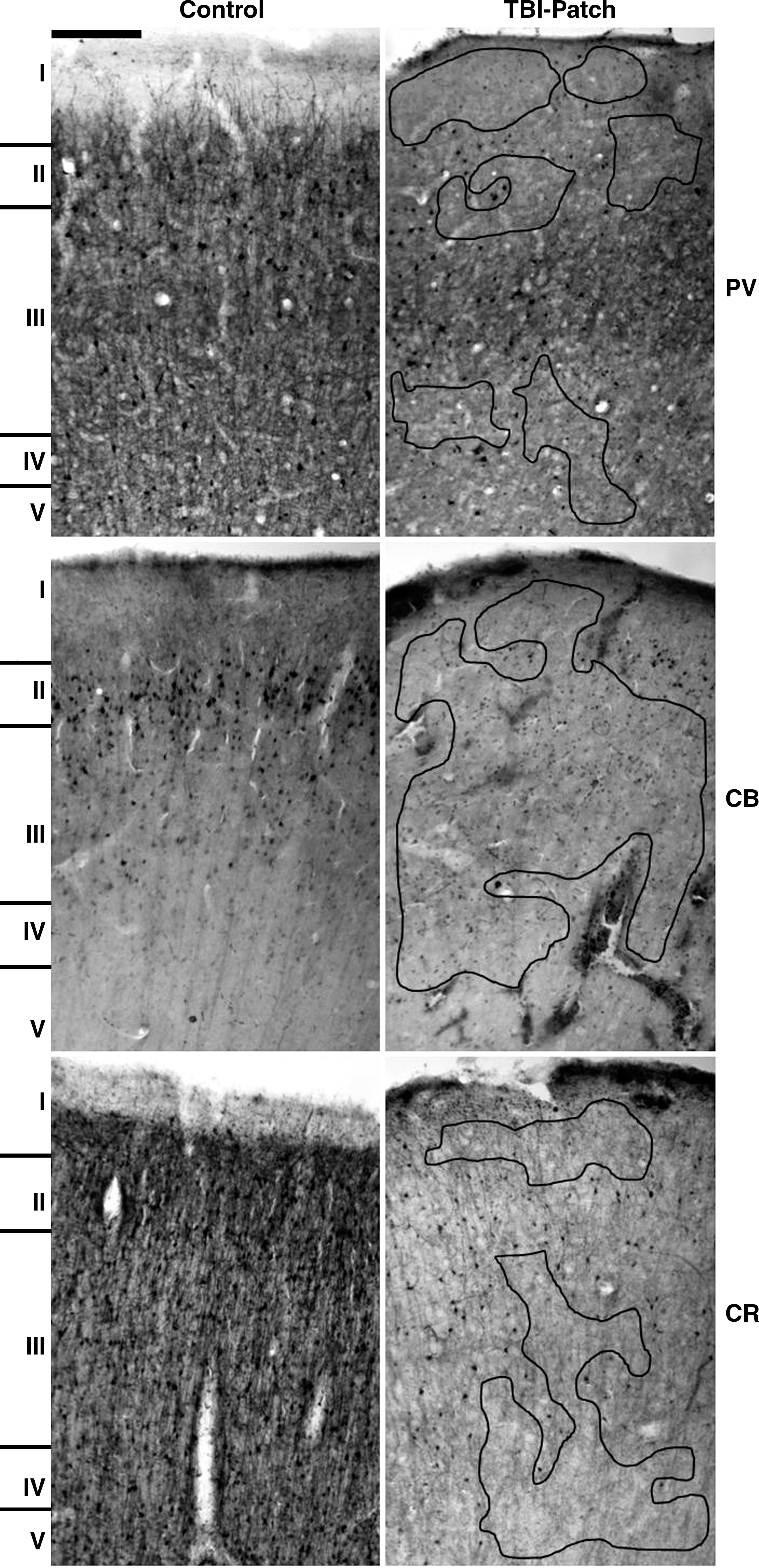
Low-magnification images (10 × ) comparing control and TBI-Patch areas for each calcium-binding protein. The areas bound by the black lines correspond to areas devoid of staining (patches) (PV, parvalbumin; CB, calbindin; CR, calretinin; scale bar = 100 μm).
In trauma samples areas with laminar preservation showed IR neurons from layers II–VI, with a remarkable decrease in PV-IR neurons, especially in layer III. Cell somas appear ovoid in shape, and their size was slightly reduced compared to controls. Dendritic processes from cell bodies located in layer III had decreased IR. When processes were seen, they appear short, discontinuous, and varicose (Fig. 4). Neuropil, clearly seen in control samples between layers III–IV and in layer I, was absent in trauma samples (Fig. 3).
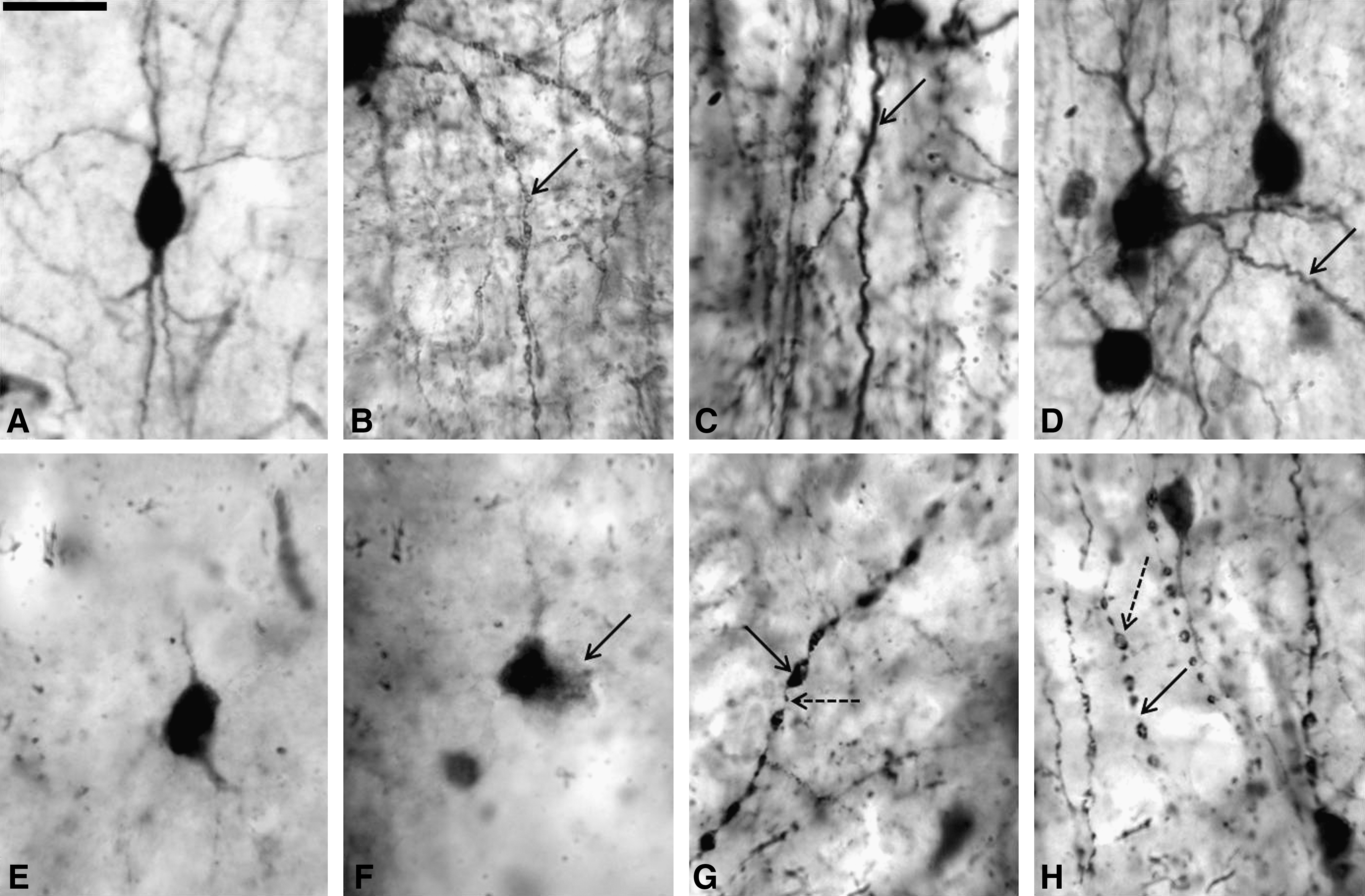
High-magnification (100 × ) morphological details of CaBPs-positive neurons in control and trauma samples. (
Sectors without laminar preservation showed patches of loss of IR, and these areas mainly corresponded to supragranular layers (Fig. 3). IR neurons on layers II–III were not homogeneous, exhibiting a gradation from moderate to mild to absent (Fig. 2). In these sectors cell somas were small in size without processes, and when visible they appear dilated and varicose (Fig. 4). Decreases in process density in layer I arising from layers II–III were also observed (Fig. 2). Tangential processes on layer I were also decreased.
When comparing homologous cortical laminas between controls and trauma samples, we found no statistically significant differences in the amount of PV-IR neurons in layers I, III, IV, V, and VI. Only layer II of the trauma samples showed smaller numbers of PV-IR neurons (p < 0.05) (Fig. 5). Comparing the entire neuronal population in the six cortical layers of the cortical column, we found no differences between control and trauma samples. When comparing numbers of infragranular and supragranular PV-IR neurons, no differences were observed between control and trauma samples.
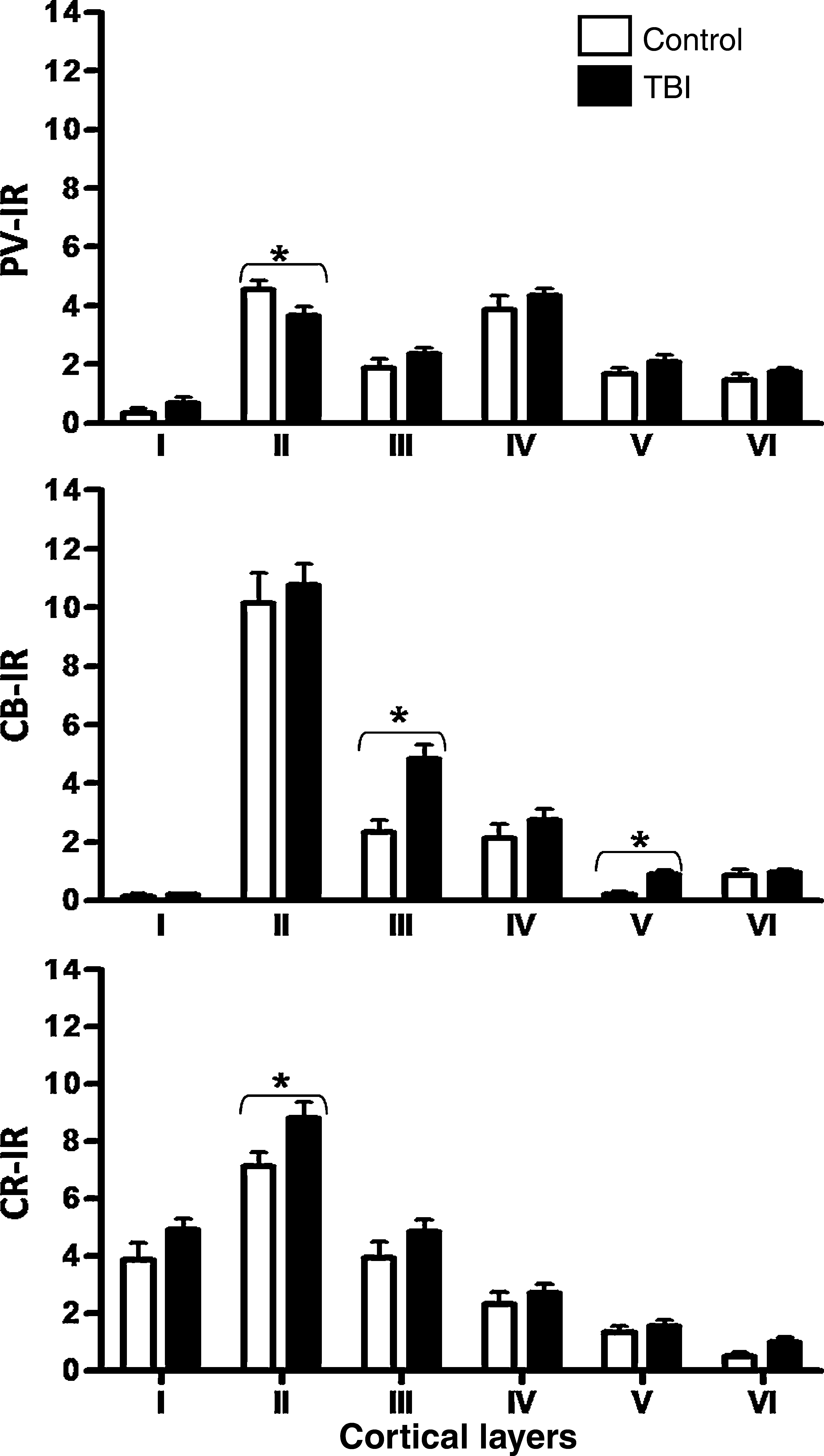
Quantification of PV-, CB-, and CR-positive neurons in sectors with preserved cytoarchitecture after human TBI. Counting was done using boxes 250 × 150 μm in size located in the center of each cortical layer. Asterisks indicate statistically significant differences (p < 0.05).
Calbindin expression
In control samples CB-IR neurons were present in layers II–VI and were more abundant in layers II and III, with their processes constituting an intensely immunoreactive band (Fig. 3); occasionally some Cajal Retzius cells were observed in layer I, with their somata oriented parallel to the pial surface. CB-IR neurons were characterized by round cell bodies that were slightly ovoid with their processes oriented vertically (ascending or descending); their morphology corresponded to bipolar, double-bouquet, or bi-tufted cells (Fig. 2 and 4A). Some multipolar forms, basket, and double-bouquet cells also appeared, with their processes radiating from soma. Occasionally some neuroglia-like cells were identified according to Condé and associates (1994), and Hof and colleagues (1999). Anti-CB antibody also stained pyramidal cells (Condé et al., 1994), including small ones in layer III and larger ones in layer V.
In trauma samples sectors with laminar conservation showed an increase in CB-IR pyramidal cells in layer III (Fig. 2), and non-pyramidal forms in the layer VI–white matter transition. Occasionally some CB-IR astrocytes were observed in infragranular layers.
Sectors with no laminar preservation (patch areas) showed decreases in immunoreactivity (Figs. 2 and 3), especially in supragranular layers (layers II–IV). CB-IR cell bodies were observed in superficial layer III, and mainly corresponded to non-pyramidal cells (Figs. 2 and 4A). Deep layers V and VI showed a discrete increase in CB-IR cell somas.
The amount of CB-IR neurons in layers I, II, IV, and VI in controls versus trauma samples, did not show any statistically significant differences. The total numbers of CB-IR neurons in all cortical layers were higher in trauma samples than in control samples (p < 0.05). Layers III and V showed the highest numbers of CB-IR neurons (p < 0.01) (Fig. 5). Additionally, the infragranular layers (V–VI) showed more CB-IR neurons than their homologous layers in control samples (p < 0.05). The supragranular layers did not show any variation.
Calretinin expression
CR-IR neurons in control tissues were present in all cortical layers (I–VI); they corresponded to bipolar and less frequently multipolar forms (Figs. 2 and 3). Denser CR-IR neurons were present in layers I, II, and III, primarily in the II–III transition. Cell bodies located in the middle and deep portions of layer III were darkly stained (Fig. 3). Layers V and VI occasionally showed CR-IR cell bodies.
TBI tissue sectors with laminar preservation showed CR-IR bodies from layers I–VI, with increases in IR intensity in the supragranular layers, particularly layer II (Fig. 2). Cell somas were ovoid and smaller than in controls, having irregular borders and dilated processes (Figs. 2 and 4).
Sectors with no laminar preservation showed decreases in IR, especially in layers II–IV (Figs. 2 and 3). Around the patches, an increase in CR-IR neurons with some of their processes dilated was observed (Fig. 4); cell somas retained their ovoid and fusiform morphology, and bipolar cells with clear vertical processes were seen. Qualitatively the highest loss of CR-IR neurons was observed in layer III (both intermediate and deep portions) and in layer IV (Fig. 3).
Although the counts showed a tendency for trauma samples to have increased numbers of CR-IR neurons in layers II and VI, statistical differences were only significant for layer II compared to control samples (Fig. 5). Comparison of total numbers of CR-IR neurons showed an increased number in trauma samples (p < 0.05), primarily in supragranular layers. Numbers in infragranular layers remained constant.
Discussion
In the present study we found a heterogeneous pattern of CaBP expression in contused tissue. Some areas were devoid of IR (patches), surrounded by areas with less dense and intense IR for CB and PV, and a slight increase in CR-positive neurons. Close to these sectors the pattern of expression of PV, CB, and CR appeared similar to those seen in control specimens, but with smaller cell bodies and shorter and more fragmented processes. Since NeuN identifies all cell types in the cortex, both pyramidal and non-pyramidal, patch areas may represent cell loss, because as reported by others IR to NeuN (Escobar et al., 2008), and in the present study to PV, CB, and CR, were also absent in those sectors.
Zones with preserved cytoarchitecture showed a small decrease in NeuN-IR, but laminar organization was clearly defined. In these sectors we observed a statistically significant decrease in PV-IR neurons in layer II, and a slight but insignificant tendency to increase in the other cortical layers. CB-IR neurons were increased significantly in layers III and V, with a slight tendency to increase in the infragranular layers. CR-IR neurons were increased in all cortical layers, but were only significantly in layer II.
Changes in the expression of PV-IR neurons have been reported in experimental models such as deafferentation or sensory deprivation (Barbado et al., 2002), and after ischemia in rats (Medina et al., 2008). Decreases in PV-IR neurons have been reported in several pathologies, such as temporal lobe epilepsy (Marco et al., 1996), and in the entorhinal cortex of post-mortem brains of patients diagnosed with bipolar disorder (Pantazopoulos et al., 2007). PV-positive neurons correspond to chandelier and basket cells (De Felipe et al., 1989; Fonseca et al., 1993). In humans and non-human primates these cells predominate in supragranular layers II–III. Axons of basket cells, which have a powerful inhibitory action, run tangentially, connecting and inhibiting pyramidal cell bodies of several adjacent cortical columns. Axons of chandelier cells also inhibit pyramidal neurons, by connecting the axon initial segment of pyramidal neurons of the same column where the interneuron is located. Both types of interneurons are also targeted by other interneurons, and also receive excitatory (glutamatergic) inputs from different sources, including pyramidal cells of the same or adjacent columns. (De Felipe, 1999). Loss of PV-positive inhibitory cells, such as those reported here, in patch areas or around these areas may produce decreases in inhibitory inputs to sectors of preserved cytoarchitecture, generating a imbalance favoring excitability. At present we do not know the fate of PV-positive cells located in preserved areas whose axons cannot reach their targets in the patch zones. We have described a loss of tangential processes running in layers III–IV and layer I, and this may indicate alterations in the inhibitory control of adjacent columns. It is important that further studies address this issue more directly.
Increases in the expression of CB-IR neurons have been reported in experimental models of ischemia (Yenari et al., 2001) and deafferentiation (Barbado et al., 2002). CB-IR cells correspond to bipolar and double-bouquet cells, and these cells are present in all layers, but predominate in layers II–III (Del Río and DeFelipe, 1997). In primates some pyramidal cells are CB-positive (Hof et al., 1999). Axons of CB-positive neurons are vertically oriented and thus restricted to the column of origin. These axons contact other interneurons and dendritic processes of pyramidal neurons (Markram et al., 2004). Excitatory axons coming from pyramidal neurons locally or distantly located connect these cells. Contrary to the PV-positive neurons, whose axons run horizontally, probably reaching sectors with different degrees of injury, CB-positive neurons possessing axons restricted to the same cortical column would be less likely to be affected by cell injury in adjacent sectors. Increases in CB-IR in areas of preserved cytoarchitecture as was demonstrated in the present study are difficult to interpret, but increases in activity due to loss of PV-positive neurons in neighboring patch areas and in the same column may be a possibility to consider.
Changes in the expression of CR-IR neurons have been documented in experimental models, such as enucleation in rats with consequent CR-IR increases in the ipsilateral colliculus, but decreases in the contralateral one (Kang et al., 2002). Increases in CR-IR neurons have also been reported in neurodegenerative diseases such as Huntington's disease (Cichetti et al., 2000) and Alzheimer's disease (Hof et al., 1993), and in temporal lobe epilepsy (Blümcke et al., 1996). CR-IR neurons appear in all cortical layers, but are more numerous in layers II and III. Their somata are frequently bipolar, their dendrites ascend and descend involving several layers, and their axons are vertically oriented. There is some evidence that CR neurons mainly contact other interneurons, affecting pyramidal activity indirectly (Del Río and DeFelipe, 1997). Increases in CR-IR close to patches as shown in this study, where PV and CB were almost absent, may indicate more tolerance to an adverse microenvironment. According to Rüttimann and associates (2004), increases in CR-IR neurons may indicate a protective mechanism against deafferentation.
In the present study we found abundant reactive astrocytes close to patch areas, especially when they were located in supragranular layers. Sectors with preserved cytoarchitecture showed fewer reactive astrocytes. It is widely known that the latter express GFAP, which is essential to keep their structure, motility, and biochemical function. It has been established also that changes in GFAP expression occur after trauma and ischemia (Eng et al., 2000). The role of astrocyte activation as expressed by an increase in GFAP-IR is controversial, because though astrocytes normally remove excessive glutamate through their transporters, after injury this function is altered, and instead of removing glutamate they can release it due to the reversed function of the transporter after lesion (Phillis et al., 2000).
In conclusion this report showed changes in the expression of CaBPs due to human TBI. CaBPs have been involved in several cell processes, such as calcium buffering, calcium transport, and regulation of multiple enzyme systems (Eyles et al., 2002). Changes in the expression of CaBPs may be subject to temporal alterations and to dynamic events that occur after TBI, that may affect the neuronal and glial metabolic states. Additionally, the present study allows us to suggest that changes in the expression of CaBPs may be associated with alterations in the circuitry of the neurons or interconnections after trauma. Furthermore, our results indicate that the GABAergic cell population may contribute to secondary events generated after TBI, such as changes in plasticity or increases in activity, and their effects on general cortical activity are currently unknown.
Footnotes
Acknowledgments
The authors wish to thanks Colciencias for financial support (code 1106-04-16329). N.G.C. is a principal investigator in Fundacao de amparo á Pesquisa do estado de Sao Paulo (FAPESP) (grant no. 2007/50261-4), FAPESP-Cinapce (grant no. 2005/56447-7,) and Conselho Nacional de Desenvolvimento científico é Technológico Programa Sud-Americano de Apoyoa a las actividades de Cooperación en Ciencia y technologia (CNPq-PROSUL) Brazil Project (grant no. 490660/2007-0). E.B., M.I.E., and H.J.P. are invited collaborators at the CNPq-PROSUL-Brazil Project. Special thanks to Olagide Wagner Castro and Artur Fernandes of the Neurophysiology and Experimental Neuroethology Laboratory for their excellent technical support and training on the use of the Olympus BX-60/Optronics/Image ProPlus image capturing system to E.B.
Author Disclosure Statement
No competing financial interests exist.