
Abstract
Sports concussion is a major problem that affects thousands of people in North America every year. Despite negative neuroimaging findings, many athletes display neurophysiological alterations and post-concussion symptoms such as headaches and sensitivity to light and noise. It is suspected that neurometabolic changes may underlie these changes. In this study we investigated the effects of sports concussion on brain metabolism using 1H-MR spectroscopy by comparing a group of 12 non-concussed athletes with a group of 12 concussed athletes of the same age (mean 22.5 years) and education (mean 16 years). All athletes were scanned 1–6 days post-concussion in a 3T Siemens MRI, and were administered a symptom scale to evaluate post-concussion symptomatology. Participants also completed a neuropsychological test battery to assess verbal memory, visual memory, information processing speed, and reaction time, and no group differences were detected relative to controls. Concussed athletes showed a higher number of symptoms than non-concussed athletes, and they also showed a significant decrease in glutamate in the primary motor cortex (M1), as well as significant decreases in N-acetylaspartate in the prefrontal and primary motor cortices. No changes were observed in the hippocampus. Furthermore, the metabolic changes in M1 correlated with self-reported symptom severity despite equivalent neuropsychological performance. These results confirm cortical neurometabolic changes in the acute post-concussion phase, and demonstrate for the first time a correlation between subjective self-reported symptoms and objective physical changes that may be related to increased vulnerability of the concussed brain.
Introduction
D
For all their associated symptoms (e.g., headache, light sensitivity, and sleep disturbances) (McCrea, 2008), sports concussions have yet to reveal any sort of consistent injury pattern that is detectable using neuroimaging (Johnston et al. 2001; McAllister, et al., 2001; Mendez et al., 2005). Both CT and MRI have yielded limited and inconsistent results. Of the few CT studies available, all conducted on boxers, the vast majority of scans came back negative for any sort of lesions or damage, with only a few of the most severe cases showing cerebral atrophy (Jordan et al., 1992; Jordan and Zimmerman, 1988; Ross et al., 1987). MRI data follow a similar pattern of results, with very few lesions being detected despite its precision, which is superior to that of CT (Jordan and Zimmerman, 1990; Newton et al., 1992).
Functional imaging techniques have fared somewhat better. Using fMRI, Chen and colleagues found different activation patterns on a working memory task for symptomatic concussed athletes compared to uninjured athletes; furthermore, they found that atypical activation patterns are highly related to symptomatology months after the injury (Chen et al., 2007, 2004, 2008). Lovell and associates (2007) and Jantzen and associates (2004) confirmed these findings in athletes imaged in the acute (within 7–10 days) post-concussion phase. PET and SPECT have yet to be used to study sports concussions specifically, but their use in mild traumatic brain injury (mTBI) has garnered mixed results, with frontotemporal regions exhibiting the most consistent impairment in patients with persistent symptomatology (Abu-Judeh et al., 1999, 2000; Bonne et al., 2003; Chen et al., 2003; Korn et al., 2005; Umile et al., 2002; Varney et al., 2001).
The inconsistency of neuroimaging findings is in stark contrast to the electrophysiological alterations that have been detected in concussed athletes using event-related potentials (ERP) and transcranial magnetic stimulation (TMS) paradigms. In symptomatic and asymptomatic athletes alike, concussions produced auditory and visual processing alterations well after the acute post-injury phase that may be reflective of brain injury (Dupuis et al., 2000; Gaetz and Weinberg, 2000; Gosselin et al., 2006). Gaetz and colleagues (2000) and De Beaumont and colleagues (2007a), respectively, demonstrated the cumulative effects of concussions on latency and amplitude changes in the P300 wave in concussed athletes months after the last incident. Another study using TMS found altered intracortical inhibition in concussed athletes that was positively correlated with the number of concussions sustained, suggesting long-term and cumulative effects in the motor system (De Beaumont et al., 2007b).
How can the concussed brain demonstrate altered electrophysiology (ERP and TMS), and detectable though variable changes in functional response (fMRI, PET, and SPECT) in the face of a brain that appears uninjured and asymptomatic according to anatomical imaging (CT and MRI)? Almost all other examples of altered function are accompanied by either a known lesion/abnormality or chronic pathological condition, including epilepsy (Goyal et al., 2008; Krendl et al., 2008; Mazerolle et al., 2007; Moran et al., 2008), Alzheimer's disease (Chapman et al., 2007; Heckemann et al., 2008; Levy-Cooperman et al., 2008; Olichney et al., 2008), or Parkinson's disease (Bokura et al., 2005; Hallevi et al., 2008; Li et al., 2005; Nandhagopal et al., 2008). It is likely that there may be micro-structural and/or metabolic changes associated with concussions that underlie the persistent alterations in electrophysiology. The subtlety of these changes has evaded traditional imaging techniques, leaving concussion poorly understood, while the underlying neural changes, be they anatomical or metabolic, have yet to be defined, particularly in the acute phase.
Magnetic resonance spectroscopy (MRS) allows the detection and quantitation of several brain metabolites in vivo, including but not limited to creatine/phosphocreatine (Cr), a general energy marker; choline-containing compounds (Cho), markers of neuronal damage and membrane turnover; myo-inositol (mI), a glial marker; glutamate (Glu), the principal excitatory neurotransmitter; and N-acetylaspartate (NAA), a marker of neuronal integrity. MRS is thus able to detect neuronal damage and changes in the chemical make-up of the brain. It is particularly useful for corroborating evidence of diffuse axonal injury, because damage-related changes to neurons are manifest not only in their physical structure, but also in their composition (Toga and Mazziotta, 2002). MRS has been used to look at the neurometabolic changes after moderate and severe TBI (Babikian et al., 2006; Garnett et al., 2000; Groswasser et al., 1987; Holshouser et al., 2006). Studies from outside of the sports literature have also demonstrated significantly depressed NAA concentrations in mTBI patients months to years after injury, despite normal findings on conventional MRI (Cecil et al., 1998; Cohen et al., 2007; Garnett et al., 2000; Govindaraju et al., 2004; Kirov et al., 2007; Signoretti et al., 2008).
There are two sports concussion studies using MRS in the acute phase, both of which report a decrease in NAA, though there is some variability in their findings (Cimatti, 2006; Vagnozzi et al., 2008). Cimatti's study looked at six different cases (four boxers) and compared baseline scan data to post-trauma scan data. The results of the study, although interesting, are variable, as changes in NAA are reported in some but not all cases. The results are also rendered difficult to interpret by a lack of specific details regarding the participant demographics. Indeed, with only six participants in the study, and the fact that previous head injuries are not accounted for, it is difficult to draw conclusions about either the state of the post-concussion brain or the utility of MRS in assessing concussion.
Vagnozzi and colleagues (2008) included the initial post-injury scan and two follow-up scans, that allowed for the time course of metabolic recovery to be charted in 14 concussed athletes of both genders ranging in age from 21–35, 10 of whom were injured while participating in a contact sport (boxing and kickboxing). Their main finding is centered on a decrease in the concentration of NAA in a single region of interest in each hemisphere of the frontal lobes. They report that concussions open a temporal window of metabolic imbalance that returns to normal levels within 30 days, but can be further exacerbated by subsequent injury within the restorative time frame.
The results of these two studies suggest that there may be structural damage in sports concussions despite the negative findings with more traditional imaging techniques (Cimatti, 2006; Vagnozzi et al., 2008). MRS is thus able to detect damage-related changes to the physical structure and the composition of neurons following a concussion; however, with only two studies investigating sports concussion with MRS on a grand total of 20 subjects, we are still prevented from making any definitive statements about the application of MRS in sports concussion, either for diagnostic or research purposes. Furthermore, the studies by Cimatti (2006) and Vagnozzi and colleagues (2008) leave many questions unanswered. The MRS protocol used in these two studies favors a less specific, but more robust, spectrum in a single region of interest (Toga and Mazziotta, 2002). Also, the control group used by Vagnozzi and colleagues was not composed of athletes, and the study population contained both men and women with a sizeable age range, and no mention of education level. To overcome these limitations, the current study used an all-male university athlete study population with a narrow age range, in recognition of the fact that concussions exert different effects on men and women (Farace and Alves, 2000; Granito, 2002; Roof et al., 1993). The current study also included more regions of interest, to investigate the potential for different effects in different brain regions, and a more elaborate, though lest robust, spectral sequence in order to investigate different metabolites.
The current study looked at the acute effects of sports concussion (within 7 days) on the chemical composition in the primary motor cortex (M1), the dorsolateral prefrontal cortex (DLPFC), and the hippocampus, as well as neuropsychological test performance, in collegiate athletes. The short post-injury time frame was chosen in light of the fact that the vast majority of athletes return to play within the first week, and some within 2 days, post-injury (Echemendia, 2006). The primary motor cortex was chosen as the region of interest based on the alterations seen in intracortical inhibition in concussed athletes (De Beaumont et al., 2007b). Likewise, the DLPFC was chosen based on both ERP studies (Dupuis et al., 2000; Gaetz and Weinberg, 2000; Gosselin et al., 2006), and fMRI studies (Chen et al., 2007, 2004, 2008; Jantzen et al., 2004; Lovell et al., 2007). The hippocampus was chosen because of its susceptibility to damage (Barr, 2005; Kandel et al., 2000), and its involvement in the generation of the P300 wave in ERP (Picton, 1992; Polich, 2004). It is hypothesized that all regions of interest will show diminished levels of NAA, increased levels of mI, and reduced levels of glutamate. We also speculate that concussed athletes will perform more poorly on neuropsychological tests; furthermore, we expect significantly different neuropsychological test performance to correlate with neurochemical changes in the region of interest (ROI) thought to underlie the abilities assessed by the test in question.
Methods
Participants
All 24 participants in this study were active players for university level intervarsity sports teams and were recruited with help from the team physician and physiotherapists. The following exclusion criteria were applied to select the athletes who took part in this study: a history of alcohol and/or substance abuse; psychiatric illness; learning disability; neurological disorders (seizure disorder, central nervous system neoplasm, or brain tumor); and TBI unrelated to contact sports. None of the athletes who participated in this study were taking psychotropic medications at the time of testing. The study was composed of two experimental groups. The control group consisted of 12 non-concussed athletes who had no history of neurological insult. The experimental group consisted of 12 athletes who suffered a sports concussion within the 6 days prior to being scanned (mean = 81.92 h, SD = 46.74 h). Concussion severity was assessed by the team physician. It varied from grade 1 (confusion for <15 min without amnesia or loss of consciousness) to grade 3 (loss of consciousness of brief duration [seconds] or prolonged duration [minutes]) according to the parameters set out by the American Academy of Neurology Quality Standards Subcommittee and Neurology (Quality Standards Subcommittee and Neurology AAo, 1997). Additionally, all head injuries were classified as mild, with Glasgow Coma Scale scores ranging from 13–15 at the time of injury. Control and concussed athletes were equivalent with regard to age (t = 0.90, p = 0.19) and level of education (t = 0.18, p = 0.74; Table 1). A standardized concussion history form was administered to obtain detailed information about the number of previous concussions (if any), the approximate date of each concussion, descriptions of the cause, and the nature and duration of relevant post-concussion symptoms (confusion and/or disorientation, retrograde and/or anterograde amnesia, or loss of consciousness). Reported symptom scores were significantly different (p = 0.003), with the concussed athletes showing a relatively high level of symptom severity on a post-concussion symptom scale from the National Football League battery (Lovell and Collins, 1998; Table 1). Symptoms were further divided into subcategories by type, including cognitive (feeling slow, feeling foggy, difficulty concentrating, or memory problems), mood (sadness, irritability, anxiety, or increased emotionality), sleep/arousal (fatigue, difficulty falling asleep, sleep changes, or loss of energy), somatic (nausea/vomiting, balance, paresthesias, or dizziness), and cranial (headache, pressure, sensitivity to light, or sensitivity to noise) symptoms (Table 2).
Reported symptom scores were significantly different (p = 0.003).
A Bonferroni correction was used for multiple comparisons to change the significant p-value from .05 to .01.
Neuropsychological testing
Neuropsychological tests from the National Hockey League neuropsychological testing program were used to assess multiple aspects of cognitive functioning (Echemendia and Julian, 2001). This battery includes classic neuropsychological tests (Lavoie et al., 2004; Lovell and Collins, 1998) selected to evaluate retrograde and anterograde amnesia (orientation questions), attention processes (Pennsylvania State University cancellation task), visual scanning and mental flexibility (Color Trails A and B and Symbol Digit Modality Test), visual memory (Brief Test of Visual Memory and incidental memory recall of Symbol Digit Modality Test), verbal memory (Hopkins Verbal Learning Test), and speech fluency (verbal and phonemic fluency). Standardized and uniform neuropsychological testing procedures were carried out by a trained neuro-psychology student for all participants.
Neuroimaging
MR imaging
All studies were performed at the Unité de Neuroimagerie Fonctionelle (UNF) of the Centre de Recherche, Institut Universitaire de Gériatrie de Montréal, using a Siemens 3-T whole-body MRI system (Siemens, Erlangen, Germany). This study was approved by the Research and Community Ethics Boards at the UNF and the Université de Montréal, and was done in compliance with the code of ethics as stated in the Declaration of Helsinki. All subjects gave informed consent following careful screening for MRI compatibility.
MR spectroscopy
We positioned our regions of interest using a rigorous anatomical localization protocol. The subjects were placed in the scanner and underwent a localizer scan prescribed parallel to the hippocampus (anterior commissure to posterior commissure [AC-PC]). Voxels were then prescribed for the hippocampus (20 × 40 × 16 mm), DLPFC (16 × 16 × 16 mm), and M1 (16 × 20 × 32 mm) of the left and right hemispheres (Fig. 1). All voxels were placed on an AC-PC–oriented oblique axial slice corresponding to the region of interest, first on a sagittal view, and then confirmed using coronal and axial views, to ensure adequate distance from ventricles, fatty tissue, and bone. Single-voxel 1H-MRS spectroscopic measurements were performed using a PRESS (Point RESolved Spectroscopy) sequence (TE [echo time] =30 msec, TR [repetition time] = 1500 msec, 256 acquisitions, 1200-Hz bandwidth, 1024 points, duration 6.5 min) on a 12-channel head coil. We opted for a moderate TR and shorter TE to balance between T1- and T2-associated signal losses and scan time, and to ensure that all six regions of interest could be captured within a reasonable period to ensure the comfort of the participants while in the scanner. Outer-volume suppression bands contiguous with the PRESS-selected volume were automatically placed in all three dimensions based on the voxel size of each ROI.
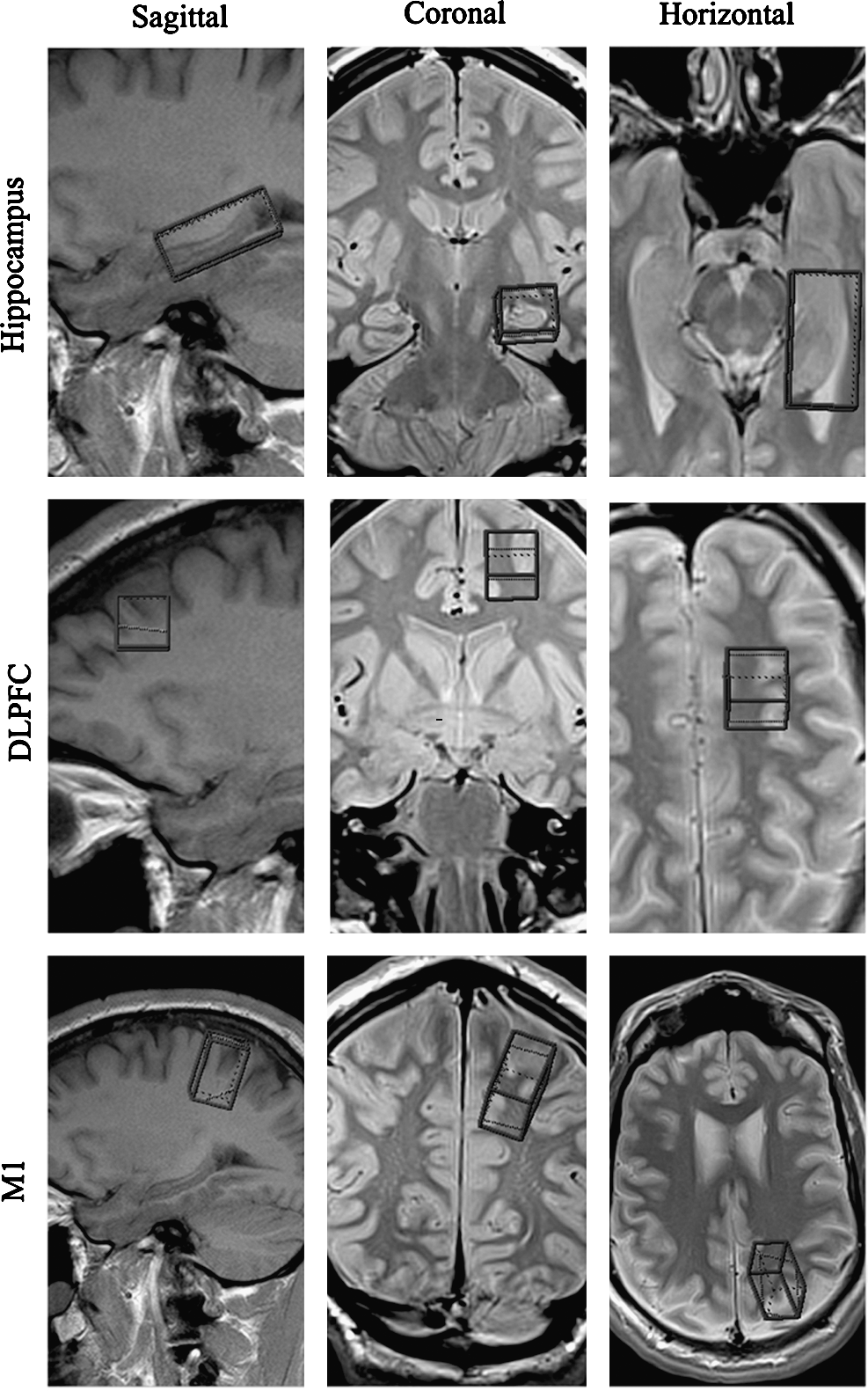
Regions of interest (ROI) for magnetic resonance spectroscopy (MRS) data acquisition depicted in the sagittal, coronal, and axial planes, in the hippocampus (top; 20 × 40 × 16 mm), dorsolateral prefrontal cortex (DLPFC, middle; 16 × 16 × 16 mm), and primary motor cortex (M1, bottom; 16 × 20 × 32 mm). Spectra were recorded in both the left and right hemispheres.
Metabolite quantitation was done using a linear combination (LC) model (Provencher, 1993), an operator-independent spectral analysis software program that estimates metabolite concentrations and their ratios relative to Cr using a set of reference spectra acquired from the individual metabolites using our MR instrument. Values were retained when uncertainties estimated as Cramer-Rao lower bounds (%SD) were <20%. The LC model operator was blind to group membership. Figure 2 displays a typical LC model output spectrum from a control participant. Values of NAA/Cr, Cho/Cr, mI/Cr, and Glu/Cr were analyzed.
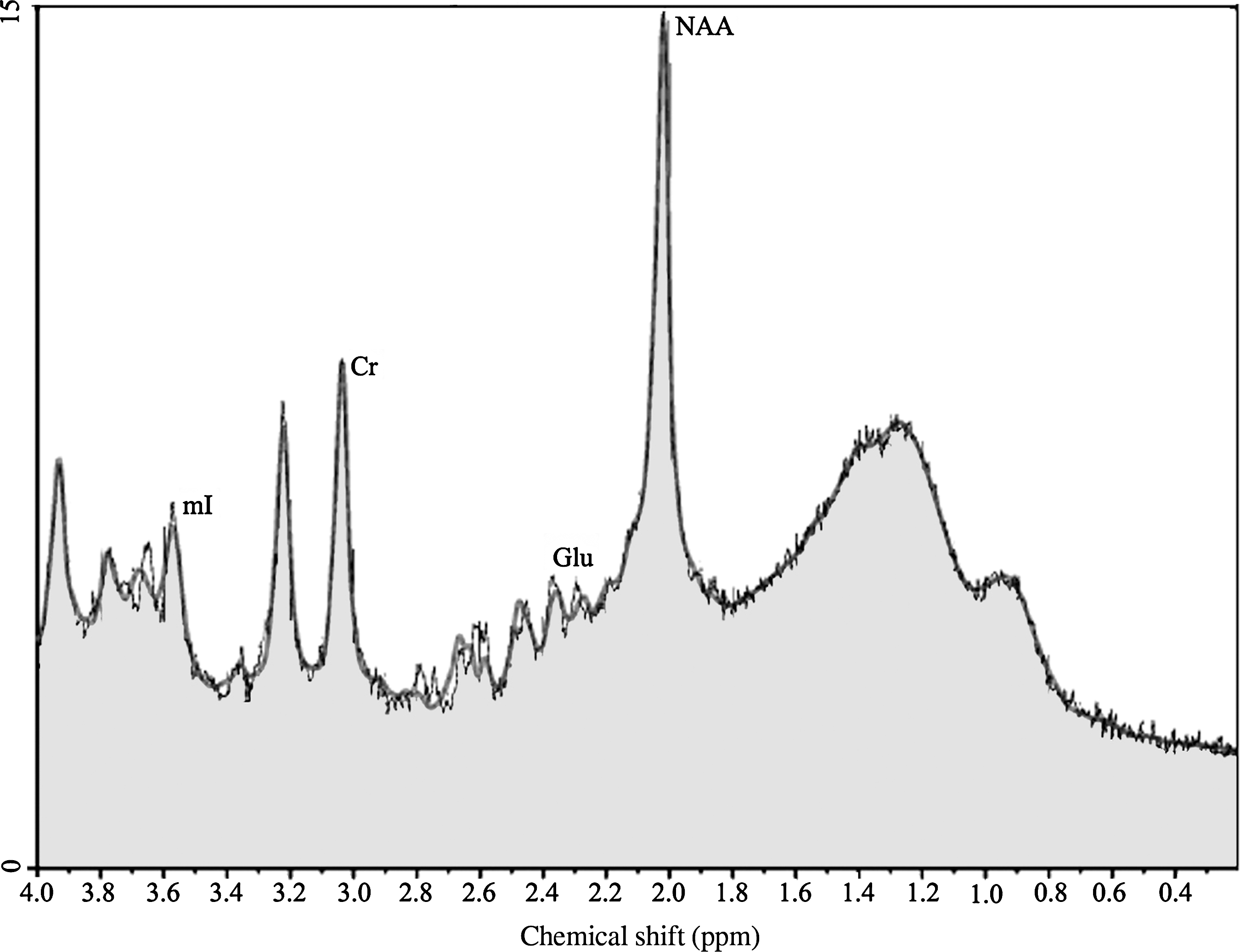
Proton spectrum of a control subject in the primary motor cortex (M1), showing the peaks corresponding to the metabolites of interest: creatine (Cr), myo-inositol (mI), glutamate (Glu), and N-acetylaspartate (NAA). Concentrations are derived from the areas under the peaks.
Statistical analysis
Statistical analyses were done using SPSS (PC version 15.0; SPSS, Inc, Chicago, IL). Coefficients of variance (CV) were calculated for each metabolite. All subsequent statistical analyses were performed only for metabolite ratios that had overall CV values <20% (i.e., NAA/Cr, Glu/Cr, and mI/Cr). The values from the left and right hemispheres were averaged for all three ROIs. The rationale for averaging the ROIs across the hemispheres is twofold. First, the literature suggests that there are no lateralization effects in the regions examined in this study (Zimmerman et al., 2008; King et al., 2008; Szentkuti et al., 2004; Geurts et al., 2004). Second, it is not known if the effects of a concussion are greater at the site of impact, or if any resulting changes are distributed diffusely regardless of impact site. As such, because it is impossible to determine the exact location of impact without the use of helmets equipped with sensors, it was decided to combine the spectra from both hemispheres within each ROI. This was confirmed in our control group across regions of interest and metabolite ratios (hippocampi: Glu/Cr t(20) = 1.06, p = 0.15; mI/Cr t(22) = −0.59, p = 0.72; NAA/Cr t(22) = 0.86, p = 0.20) (DLPFC: Glu/Cr t(22) = −0.18, p = 0.57; mI/Cr t(22) = 0.0, p = 0.50; NAA/Cr t(22) = −1.45, p = 0.92); (M1 Glu/Cr t(22) = −2.18, p = 0.98; mI t(22) = 0.05, p = 0.48; NAA t(22) = 0.26, p = .40). Though the Glu/Cr and NAA/Cr levels are near significance in the hippocampi, this is not considered to be suggestive of a lateralization effect, but rather reflective of three control participants who are driving the nonsignificant differences. It should be noted that one participant was excluded from the Glu/Cr ratio in the hippocampus, as he had an abnormally high value on the left side (>3.29 SD). The different metabolites in any given voxel are unrelated in principle and are not correlated (Braun et al., 2002). As such, the metabolite ratios of the two groups were compared using Student's independent t-tests.
Student's t-tests were carried out for each neuropsychology test. Also, in order to look at the neuropsychological performance of the control athletes and the concussed athletes, all test scores were converted into Z-scores to derive an overall neuropsychology battery composite score. Pearson correlations were performed on significant neurometabolic findings and significant clinical findings, with a Bonferroni correction for multiple comparisons. The effect sizes of all inter-group comparisons are reported as Cohen's d.
Results
Neuropsychological testing
Performance was equivalent between control and concussed athletes across all neuropsychological tests used to assess the various cognitive domains (Table 3). Due in part to a lack of statistical power, no between-group differences reached significance across all neuropsychological tests used to assess the various cognitive domains (Table 3).
Neurological testing did not reveal any significant differences across seven different tests, including measures of learning/memory (Hopkins Verbal Learning Test, Brief Test of Visual Memory [BTVM], and incidental memory recall of the Symbol Digit Modality Test [SDMT]), attention (the Pennsylvania State University [PSU] cancellation task), visual scanning and mental flexibility (Color Trails A and B), and speech fluency (verbal and phonemic fluency).
Magnetic resonance spectroscopy
Overall spectroscopy values across groups were quite stable with very little variance, due in large part to the relative quantification technique used. There were no significant differences between control and concussed subjects in the hippocampi. Glu/Cr was statistically equivalent (t(22) = 0.55, p = 0.59), as were mI/Cr (t(22) = −1.176, p = 0.25) and NAA/Cr ratios (t(22) = 0.45, p = 0.66) (Fig. 3A). Results from the DLPFC (Fig. 3B) revealed a mixed pattern of changes. While there were no differences in Glu/Cr (t(22) = 0.40, p = 0.70) and mI/Cr (t(22) = 0.28, p = 0.78), there was a significant difference in NAA/Cr (t(22) = 2.36, p = 0.03, d = 0.99), suggesting at least a partial metabolic disruption in the acute phase. M1 revealed an even stronger pattern of metabolic alterations (Fig. 3C). Glu/Cr levels were lowered significantly (t(22) = 3.47, p = 0.004, d = 0.83). mI/Cr levels remained unaffected in M1 (t(22) = 0.18, p = 0.86), but similarly to the change seen in the DLPFC, NAA/Cr ratios were significantly affected, with concussed athletes showing reduced levels (t(22) = 2.912, p = 0.008, d = 1.16) (Fig. 3C).
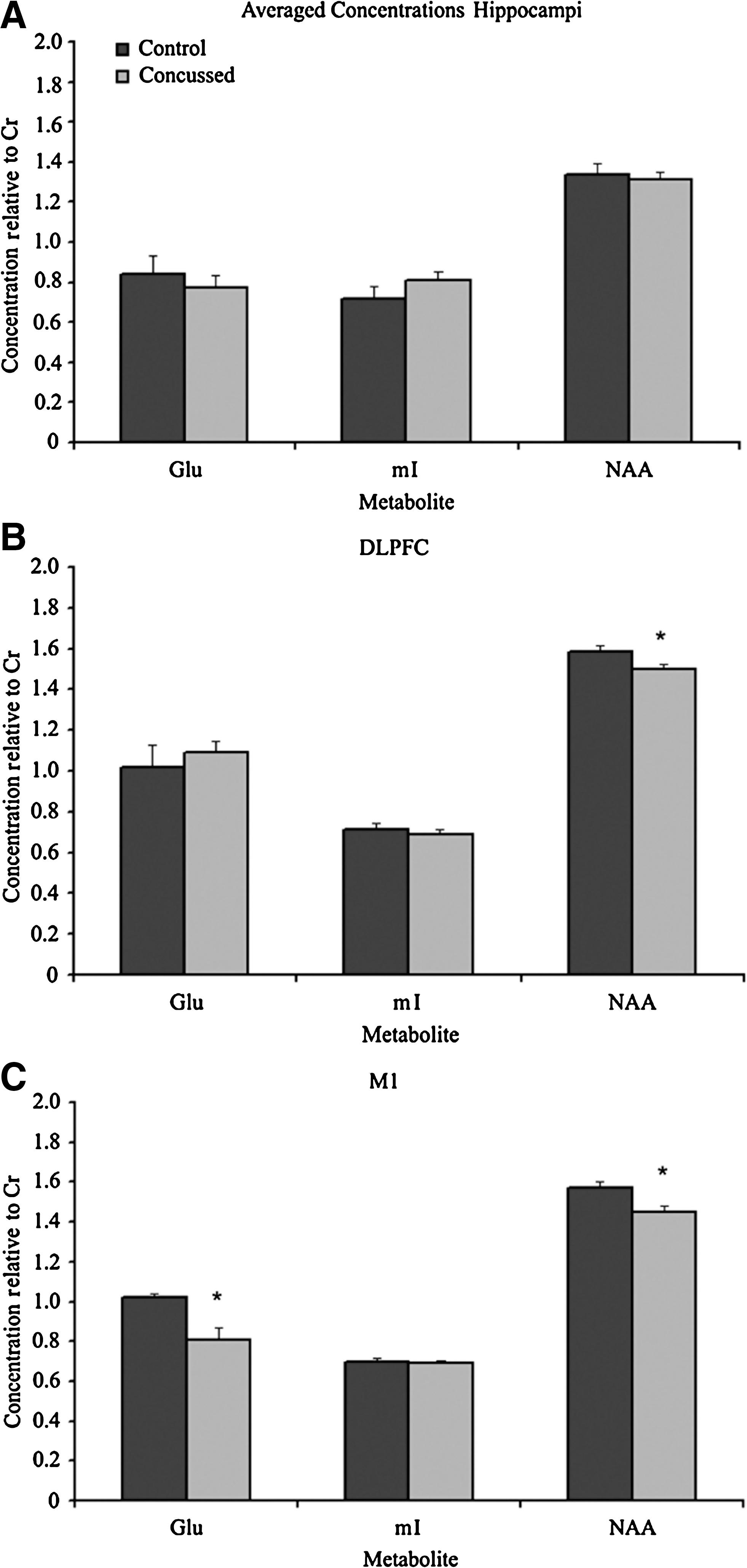
Bar graphs of the mean Glu/Cr, mI/Cr, and NAA/Cr metabolite ratios for control (black bars, n = 12) and concussed (gray bars, n = 12) athletes in (
Correlations between significant metabolite changes and symptom severity were carried out, as symptom severity was the only clinical variable distinguishing concussed from non-concussed athletes. Correlations between NAA/Cr and self-reported symptom scores were not significant in the DLPFC (r = −0.132, p = 0.559). However, NAA/Cr in M1 was significantly negatively correlated with self-reported symptom scores (r = −0.554, p = 0.008) (Fig. 4A). Similarly, Glu/Cr in M1 also showed a significant negative correlation with symptom severity (r = −0.637, p = 0.001) (Fig. 4B). Two athletes were excluded from the correlation analysis, one control and one concussed. The control athlete was excluded for an unusually high symptom score that was related to somatic pain and cognitive stress related to an impending knee surgery. The concussed athlete was also excluded for an abnormally high symptom score that had him more than three standard deviations from the group norm.
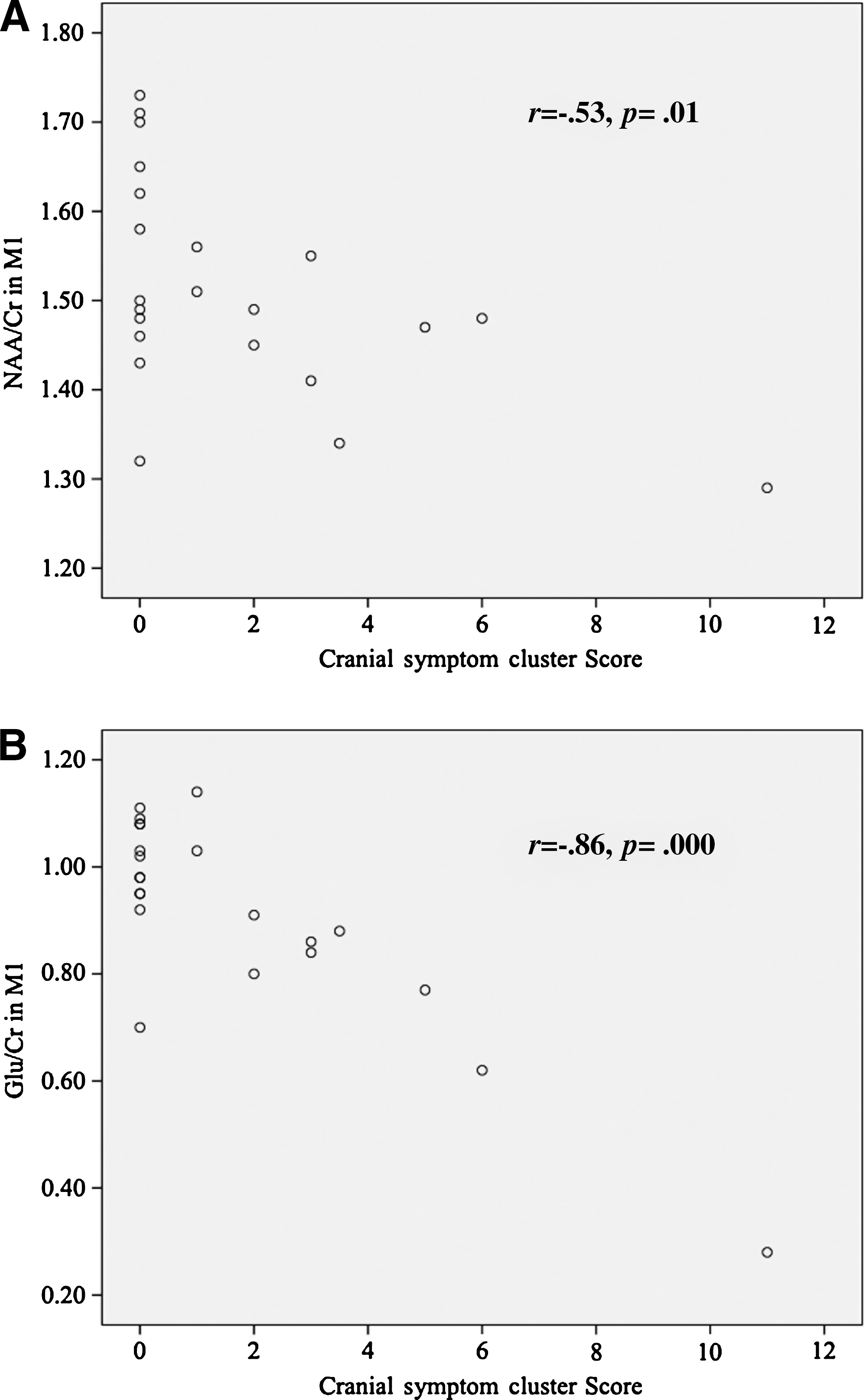
(
Further post-hoc correlations were performed on neurometabolites and symptom clusters, including cognitive (feeling slow, feeling foggy, difficulty concentrating, and memory problems), mood (anxiety, sadness, irritability, and increased emotionality), sleep/arousal (fatigue, difficulty falling asleep, sleep changes, and loss of energy), somatic (nausea/vomiting, balance, paresthesias, and dizziness), and cranial (headache, pressure in the head, sensitivity to light, and sensitivity to noise) categories (Table 2) using a Bonferroni correction for multiple comparisons. The post-hoc symptom-neurometabolite correlations in M1 did not reveal significant correlations for NAA/Cr and cognitive symptoms (r = −00.3, p = 0.18), mood symptoms (r = −0.24, p = 0.28), sleep/arousal symptoms (r = −0.36, p = 0.10), or somatic symptoms (r = −0.43, p = 0.05). The correlation between NAA/Cr and cranial symptoms in M1 was equal to the cut-off value after multiple comparisons (r = −0.53, p = 0.01). The same pattern of results was found for Glu/Cr and cognitive symptoms (r = −0.491, p = 0.02), mood symptoms (r = −0.179, p = 0.43), sleep/arousal symptoms (r = −0.047, p = 0.84), and somatic symptoms (r = −0.37, p = 0.09), with only the Glu/Cr-cranial symptoms correlation in M1 being significant after correcting for multiple comparisons (r = −0.86, p = 0.000).
Discussion
In this study we investigated the neuropsychological and neurometabolic differences between 12 non-concussed athletes and 12 concussed athletes of similar age and education level. Neuropsychological testing did not reveal any significant differences between control and concussed participants. Despite normal neuropsychological test results and anatomical MRI results, concussed athletes reported post-concussion symptoms and had altered neurometabolic profiles. The neurometabolic alterations, as measured using 1H-MR spectroscopy, were not seen in all ROIs. There were no changes detected in the hippocampus within the acute post-injury time frame. However, there were significant alterations in cortical areas. Depressed levels of NAA were seen in the DLPFC and M1. Furthermore, glutamate levels were depressed in M1, though a similar depression was not seen in the DLPFC. These results are important in light of the typical recovery window (7–10 days) and return-to-play protocols commonly used by athletic teams for sports concussion. This study demonstrates that even though an athlete may perform at typical neuropsychological test levels, this does not mean that the brain has returned to its pre-injury state. The MRS results of the current study correlate with the symptoms reported by the concussed athletes. Vagnozzi and associates (2008) concluded that the closure of metabolic brain imbalance (as measured by MRS) did not correlate with the closure of clinical symptoms. By contrast, in this study we report a correlation between the MRS findings and the self-reported symptoms.
Indeed, our results did not reveal any significant between-groups differences in neuropsychological test performance. The absence of differences is common to many sports concussion studies (De Beaumont, et al., 2006), especially using paper-pencil testing (Broglio et al., 2007; Grindel et al., 2001), a testament to the elusiveness of the pathology, as well as to the insensitivity of the majority of available test batteries. Another reason why we did not find any neuropsychological deficits is the mild nature of the injuries, in concert with the education levels of the athletes (Grindel et al., 2001). This is in sharp contrast with the subjective report of post-concussive symptoms, which were found to correlate with metabolite changes in cortical areas. Without doubt, the major problem with respect to the current study and the neuropsychological test results is the sample sizes. Indeed, those studies in which neuropsychological differences are found generally have sample sizes three or four times larger than that used in the current study (Echemendia and Julian, 2001; Gosselin et al., 2006; Collins et al., 2003; Lau et al., 2009). With a more sensitive battery and a larger sample size, neuropsychological differences are more likely to emerge in a pattern consistent with what is reported in studies in which neuropsychological testing is the focus (Echemendia and Julian, 2001; Gosselin et al., 2006; Collins et al., 2003; Lau et al., 2009). While the small sample size means that our neuropsychological test results should be taken with a grain of salt, it also underscores the strength of the effect of the neuroimaging data.
Given the susceptibility of the hippocampus to damage due to its histological properties (Barr, 2005; Kandel et al., 2000), and its role as a P300 wave modulator (Fushimi et al., 2005), we expected to see changes in the hippocampus; however, from a biomechanical standpoint it is not surprising that we did not find any changes in this ROI in the acute phase. The anatomical location of the hippocampus is further removed from any initial impact delivered to the head at the time of injury relative to the other two ROI. By virtue of its histology and location, pathological changes reach the hippocampus at a different rate than the cortex (Elkin and Morrison, 2007). This means that the brain is not uniform and that different regions respond differently to the same mechanical forces. This is corroborated by the nature and extent of hippocampal injury in moderate to severe TBI. According to recent animal research using a fluid percussion model, pathological changes in the hippocampus correlate with the severity of the injury (Golarai et al., 2001; Hellmich et al., 2005), and injury to the hippocampus increases with the force of the impact. It is therefore less likely for damage to be detectable in the hippocampus with less severe injuries. The hippocampus is still capable of being injured by a mTBI, but the relative mildness of a sports concussion makes it less likely than an injury to the neocortex. However, it must also be noted that other researchers (Yoshino et al., 1991, 1992) have found metabolic changes in the hippocampus subsequent to fluid percussion injury. Yoshino and colleagues (1991) found that cells in areas not mechanically damaged had dramatically altered functioning in terms of their glucose metabolism, and that this was seen bilaterally, though it was more pronounced ipsilateral to the injury. Of note, these changes were more pronounced in the neocortex and the hippocampus. Yoshino and associates (1992) further discovered that these metabolic changes were directly related to glutamatergic projections within the hippocampus. We did not find a similar depression of glutamatergic metabolism within the hippocampus. This may be due to differences between sports concussion and the fluid percussion model. It may also be due to biomechanical differences related to the differences in head-cervical anatomy between bipedal and quadruped animals, or it may be due to limitations inherent to the imaging technique used in this study. The nonsignificant increase in mI may be indicative of gliosis, a common feature of neural scarring, a process that can take years to appear in TBI patients (Diaz-Arrastia et al., 2000; Golarai et al., 2001; Swartz et al., 2006).
Changes to DLPFC function are perhaps the best-documented in the sports concussion literature, with changes reported in neuropsychology (Broshek and Freeman, 2005; Gosselin et al., 2006), electrophysiology (De Beaumont et al., 2007a; Gosselin et al., 2006), and functionality (Chen et al., 2004; Lovell et al., 2007). Consistent with the changes seen in the DLPFC and the existing literature (Cimatti, 2006; Vagnozzi et al., 2008), our results showed a decrease in NAA/Cr in M1. The decrease in NAA levels in M1, as in the DLPFC, indicates a change in the metabolism of neurons. Understanding the biosynthesis of NAA further supports the notion that the cost of NAA is a decrease in ATP production (Moffett et al., 2007). When the brain is in homeostatic balance, this cost is not high, and proper levels of NAA are maintained. However, when the brain is injured, in this case by a concussive blow, that metabolic balance is shifted. Concurrent with the initial spike and subsequent decrease in cerebral blood flow (Nilsson and Nordstrom, 1977a; Yuan et al., 1988), concussive injuries also cause a hyperexcited brain state via a mechanically-elicited neuronal discharge resulting in excitatory activity (Nilsson and Nordstrom, 1977b; Zimmerman and Putnam, 1947), that is not restricted to mechanoreceptors (Hu and Lewin, 2006; Morris, 1990; Sachs, 1991). This hyperexcited state puts a high metabolic demand on restoring the resting state, a key aspect of which is the production of ATP, resulting in a decrease of NAA. Vagnozzi and associates (2008) indirectly support this by charting the recovery of NAA levels, which return to typical levels at 30 days post-injury.
Though limited to M1, we are the first to report a decrease in Glu/Cr in sports concussion. The Glu/Cr decrease is corroborated by the prolongation of the cortical silent period (CSP) in M1 reported by de Beaumont and colleagues (2007b). The CSP is a pause in an otherwise continuous series of electrophysiological events. This suggests that the receptors of intracortical inhibitory interneurons of the motor system may be particularly vulnerable to the effects of sports concussions. Though diminished Glu/Cr levels do not speak to changes in intracortical inhibition, they do further suggest that M1 as a ROI is affected by concussive blows despite normal neuropsychological performance. The augmentation of the inhibitory action speculated to underlie the prolongation of the CSP may be further bolstered by the diminished levels of glutamate, which is chiefly responsible for excitatory activity in the brain.
It is not clearly understood why Glu/Cr levels are so drastically affected in M1, while they remain statistically equivalent in the DLPFC. Histological differences between M1 and DLPFC may account for some of the difference. Betz cells are giant pyramidal cells found only in layer V of M1 (Barr, 2005), and are thought to be excitatory. Thus while both M1 and DLPFC possess layer V pyramidal cells, only M1 is equipped with the giant Betz cells that are adapted to facilitate excitatory transmission to descending pathways. It is possible that the pushing of the top of the brain against the cranial vault that occurs during rotational forces in a concussion (Bayly et al., 2005) may affect the biosynthesis of glutamine in astrocytes or its hydrolyzation into glutamate in excitatory neurons. The downstream effect may be manifest as diminished levels of glutamate, as seen in M1. Also, the absence of Betz cells in the DLPFC may be involved in its exemption from glutamatergic alterations.
The breadth of the metabolic changes in M1 (decreases in NAA/Cr and Glu/Cr) in the concussed athletes might also be explained by the biomechanics of head injury (Bayly et al., 2005; Denny-Brown and Russell, 1940; Holbourn, 1945). The rotational and linear forces at work that produce a concussion (Bayly et al., 2005; Denny-Brown and Russell, 1940; Greenwald et al., 2008; Holbourn, 1945; Mihalik et al., 2007) suggest that M1 is consistently vulnerable to the white matter injury of shear strain. The differential effects on Glu/Cr in M1 versus the DLPFC are further corroborated by changes detected using diffusion tensor imaging, namely that patients who had suffered a mTBI demonstrated reduced fractional anisotropy in the corticospinal tract, indicating diffuse axonal injury, but no such injury pattern was found in the frontal regions (Kraus et al., 2007).
Interestingly, the post-hoc correlations by symptom type (cognitive, sleep/arousal, mood, somatic, and cranial) revealed a significant correlation of both NAA/Cr and Glu/Cr with cranial symptoms, including headache, pressure in the head, sensitivity to light, and sensitivity to noise. Within this symptom cluster, feeling pressure in the head, sensitivity to light, and sensitivity to noise, can all be considered symptoms of headache (IHS HCCot., 1997). Headaches themselves are common as a post-traumatic symptom (Gibbs, 1994; Matthews, 1972; Williams and Nukada, 1994a, 1994b). Indeed, post-traumatic cranial symptoms may be related to the pathophysiology of concussion (Alves et al., 1986), such as diffuse axonal injury or excitotoxic amino acid release and subsequent neural injury (Giza and Hovda, 2001). Furthermore, altered cerebral hemodynamics and slowed cerebral circulation have been described in the post-concussion syndrome (Gilkey et al., 1997), and are likely due to vasomotor instability (Alves et al., 1986), resulting in a decrease in regional cerebral blood flow (Gilkey et al., 1997).
Significant correlations between changes in NAA/Cr and symptoms, as well as Glu/Cr and symptoms in M1 are clinically important. Indeed, the diagnostic and prognostic importance of post-concussion symptoms underscores the need to establish a less subjective metric to measure recovery without the confounds of either lying about symptom resolution to return before it is safe to do so on one extreme, to embellishing symptoms for litigious or compensatory reasons on the other extreme. Neurometabolic changes in M1 may provide such a means. The results of this study demonstrate a link between self-reported symptoms, even in the relatively minor injury of sports concussion, and neurometabolic alterations. Though these correlations do not prove causation between metabolic depression and experienced symptoms, they do lend credibility to the relevance of self-reported symptoms. It is important to note that the neurometabolic changes reported in the current study, and the subsequent correlations with reported post-concussion symptoms, were found using a small sample size. However, the effect sizes are robust, and indicate a consistent metabolic alteration that is in accordance with the accepted pathophysiology of concussion (Giza and Hovda, 2001). Changes in neurometabolic balance also open the door for questions about the use of pharmaceuticals in the management and perhaps even prevention or attenuation of concussions. These changes in neurometabolism represent the first step in characterizing the physical consequences of suffering a blow to the head, and constitute a step forward in understanding the pathophysiology of concussion.
Footnotes
Acknowledgments
This research was funded by a CIHR Banting and Best CGS-D to LCH as well as a CIHR research chair to ML and CIHR research funds to DE. Special thanks to A. Khiat for his expertise in analyzing the MR specta.
Author Disclosure Statement
No conflicting financial interests exist.