
Abstract
Taurine has multiple functions in the central nervous system (CNS), serving as an osmoregulator, antioxidant, inhibitory neuromodulator, and regulator of intracellular Ca2+ flux. Since the role of taurine in traumatic spinal cord injury (SCI) is not fully understood, the present study was conducted with C57 black/6 mice (18–20 g) who underwent severe SCI at the Th-8 level using a weight compression device. Taurine was injected intraperitoneally at doses of 25, 80, 250, and 800 mg/kg within 30 min after SCI. Controls were injected with saline. The contusional cord segments were removed 6 h after SCI, and concentrations of interleukin-6 (IL-6) and myeloperoxidase (MPO) were measured using ELISA kits. Phosphorylation of STAT3, which is activated by IL-6, and expression of inducible cyclooxygenase-2 (COX-2) were also compared between the taurine treatment group (250 mg/kg) and the control group by Western blot analysis. Morphological changes were evaluated with H&E-stained sections. Taurine significantly decreased IL-6 and MPO levels in a dose-dependent manner, significantly reducing the phosphorylation of STAT3 and expression of COX-2 after SCI compared to controls. A reduced accumulation of neutrophils, especially in the subarachnoid spaces, and secondary degenerative changes in gray matter were also noted, and motor disturbances were significantly attenuated with taurine treatment (250 mg/kg). These findings indicate that taurine has anti-inflammatory effects against SCI, and may play a neuroprotective role against secondary damage, and thus it may have therapeutic potential.
Introduction
S
Taurine, 2-aminoethane sulfonic acid, is conditionally essential in humans and is ubiquitously expressed in many tissues. It acts as an antioxidant, osmoregulator, calcium regulator, and membrane stabilizer (Huxtable, 1992), and has been shown to be beneficial in the treatment of congestive heart failure (Azuma, 1994), ischemia-reperfusion injury (Milei et al., 1992), pulmonary fibrosis (Giri and Wang, 1992), and endothelial cell cytotoxicity (Wang et al., 1996b). Neutrophils contain high concentrations of taurine in their cytosol (Fukuda et al., 1982), and they play important roles in phagocytosis after SCI as a host defense mechanism against infection. However, the effects of taurine under pathophysiological circumstances, for example after SCI, have hitherto remained unclear (Gupta et al., 2006).
The purpose of the present study was thus to investigate any protective effects of exogenous taurine in a mouse spinal cord compression model. To elucidate its influence on inflammatory responses, the IL-6 concentration, the degree of phosphorylation in an IL-6 signal transduction pathway, expression of inducible cyclooxygenase-2 (COX-2), and myeloperoxidase (MPO) level were all measured in injured spinal cords. We also examined the impact of taurine on hindlimb motor function after SCI.
Methods
Materials
Unless otherwise specified all chemicals were from Sigma Chemicals Co. Ltd. (St. Louis, MO).
Animals
Female mice (C57BL/6NCrj; Charles River Japan, Inc., Yokohama, Japan) 8–10 weeks of age (weight 18–20 g) were housed two or three per cage and kept at a temperature of 24°C with free access to water and food before and after surgery. The experiments were carried out in accordance with the National Institutes of Health guidelines for the care and use of laboratory animals, with approval of the appropriate committee of Nagoya University Graduate School of Medicine. All efforts were made to minimize the number of animals used and their suffering.
Spinal cord injury model and drug treatment
The mice were anesthetized with 1.5% halothane and maintained on 1.25% halothane in an oxygen/nitrous oxide (30%/70%) gas mixture. Temperature was monitored with rectal probes and maintained between 36.5° and 37.5°C with heating pads and lamps. The mice were fixed with a stereotaxic apparatus and subjected to severe spinal cord compression as previously described by Farooque (2000; Supplementary Fig. 1s) (see online supplementary material at
Analysis of the IL-6 concentration and myeloperoxidase level
Spinal cord tissue samples for enzyme-linked immunosorbent assay (ELISA) were prepared using the following buffer: 10 mmol/L Tris-HCl (pH 8.0), 150 mmol/L sodium chloride, 1 mmol/L EDTA, 1 mmol/L PMSF, 1% Triton X-100, 10 μg/mL aprotinine, and 1 μg/mL pepstatin. Homogenates were centrifuged at 18,000g at 4°C for 15 min, and protein concentrations of the supernatants were determined by the method of Bradford using bovine serum as the standard. The concentration of IL-6 and MPO level were measured using sandwich ELISA kits (R&D Systems, Inc., Minneapolis, MN, and HyCult Biotechnology, Uden, The Netherlands, respectively), according to the manufacturer's instructions.
Analysis of phosphorylation of signal transducers and activators of transcription 3 (STAT3) and expression of COX-2 by Western blot analysis
Spinal cord tissue samples of each group were collected at 6 h after SCI and homogenized on ice in 300 μL of buffer and phosphatase inhibitor solution: 50 mmol/L Tris base/HCl (pH 7.5), 0.2 mmol/L EGTA (pH 7.5), 0.2 mmol/L EDTA (pH 8.0), 0.2 mmol/mL phenylmethylsulfonyl fluoride (PMSF), 1 μg/mL pepstatine, 0.2 μg/mL aprotinine, 2 μg/mL leupeptine, 0.1 mmol/L dithiothreitol, 1 mmol/L sodium orthovanadate (Na3VO4), 50 mmol/L sodium fluoride (NaF), 2 mmol/L sodium pyrophosphate (Na4P2O7 · 10H2O), and 1% Nonidet P-40, for Western blot analysis. The homogenates were then centrifuged at 18,000g at 4°C for 15 min, and protein concentrations of the supernatants were determined by the method of Bradford using bovine serum as the standard. Crude supernatant samples containing 25 μg of protein were subjected to 7.5% SDS-PAGE, and the proteins were transferred to polyvinylidene difluoride (PVDF) membranes and incubated with primary antibodies against phosphorylated (p)- Tyr705-STAT3 (Cell Signaling Technology, Inc., Beverly, MA) at a dilution of 1:500, actin at a dilution of 1:3000, and COX-2 (BD Biosciences Pharmingen, Franklin Lakes, NJ) at a dilution of 1:500, for 1 h at room temperature. After washing, the membranes were incubated with goat anti-rabbit polyclonal or anti-mouse monoclonal IgG conjugated to horseradish peroxidase at a dilution of 1:3000 for 30 min at room temperature. Reactions were developed with ECL (GE Healthcare, Buckinghamshire, U.K.). Phosphorylated-Tyr705-STAT3 immunoblots were stripped from PVDF membranes and reblotted with primary antibodies against STAT3 (BD Biosciences Pharmingen) at a dilution of 1:500 for 1 h at room temperature. Exposure to the secondary antibody, goat anti-mouse monoclonal IgG, was at a dilution of 1:3000 for 30 min, followed by color development. Band intensities were quantitated by densitometric scanning using the NIH IMAGE program.
Histological examination of the spinal cord
Mice killed 6 h or 4 weeks after SCI were transcardially perfused with a phosphate-buffered solution of 10% formaldehyde. The spinal cord at Th-8 was then immediately removed and immersed overnight in the same solution. Transverse semi-serial paraffin-embedded sections 10 μm thick were prepared and stained with hematoxylin and eosin (H&E). An experienced pathologist blinded to study group assessed the sections.
Behavioral analysis
To investigate the effects of taurine on SCI, we measured recovery of hindlimb motor function using the Basso mouse scale (BMS) motor rating scale. Severe SCI mice were treated with taurine (n = 6; 250 mg/kg IP within 30 min after induction of SCI, and then once a day from day 1 until day 7), or saline (n = 7; IP on the same schedule). Two investigators observed each animal for 5 min and scored (0–9 scale) each hindlimb at 1, 2, 4, 7, 10, 14, 17, 21, 24, and 28 days after SCI. The scores were averaged to obtain a single value per animal for each time point.
Statistical analysis
Data are expressed as mean ± SEM values. The statistical significance of the differences between the groups was assessed by one-way ANOVA with the Fisher's post-hoc test. Densitometric ratios of p-STAT3:STAT3 and COX-2:actin were compared between groups using the Mann-Whitney U test. Statistical significance was set at p < 0.05.
Results
Effects of taurine on IL-6 concentrations after SCI
We reported earlier that severe SCI induced a significant increase of IL-6 from 3–24 h after SCI (Yamauchi et al., 2006). In the present study, the concentrations of IL-6 in intact control spinal cords and injured spinal cords treated with saline at 6 h after SCI were 3.0 ± 0.7 and 396 ± 28 pg/mL, respectively. Animals with laminectomy without compression treated with taurine had slightly induced expression of IL-6. Taurine caused significant decreases at the doses of 80, 250, and 800 mg/kg compared to mice treated with saline (Fig. 1).
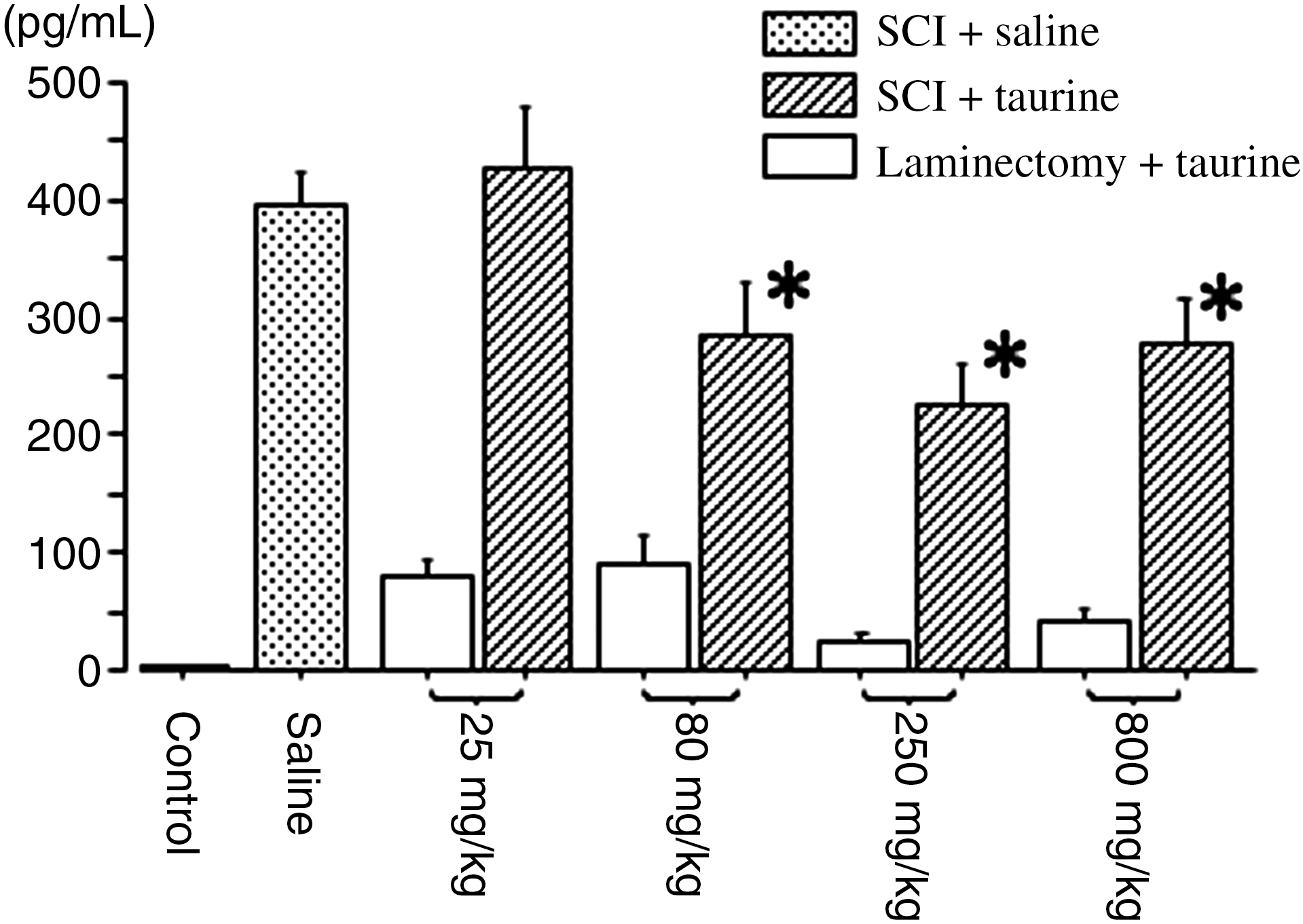
Concentrations of interleukin-6 (IL-6) in spinal cord at 6 h after severe spinal cord injury (SCI) or sham surgery. Taurine was injected within 30 min after induction of severe SCI (n = 7 per group), or laminectomy without compression (n = 6 per group), at doses of 25, 80, 250, and 800 mg/kg. The concentrations of IL-6 in spinal cord were measured using an ELISA kit. Mean ± SEM values of the data are shown (Saline, mice suffering from severe SCI and treated with saline [n = 7]; Control; intact control spinal cord). Concentrations of IL-6 in the intact control spinal cord were 3.0 ± 0.7 pg/mL (n = 6; *p < 0.05 indicates significant difference between saline and taurine treatment as assessed by analysis of variance followed by the Fisher's post-hoc test; ELISA, enzyme-linked immunosorbent assay; SEM, standard error of the mean).
Effects of taurine on activation of STAT3 following SCI
We have also reported that severe SCI is associated with activation of the JAK/STAT3 signaling pathway, which transduces signals from the cell surface to the nucleus in response to IL-6 (Yamauchi et al., 2006). Western blot analysis revealed equal levels of actin and STAT3 in crude samples (Fig. 2 lower panel). Densitometric analysis of bands of p-Tyr705-STAT3/STAT3 revealed significant suppression in those treated with taurine (SCI + taurine) compared with those treated with saline (SCI + saline) (Fig. 2).
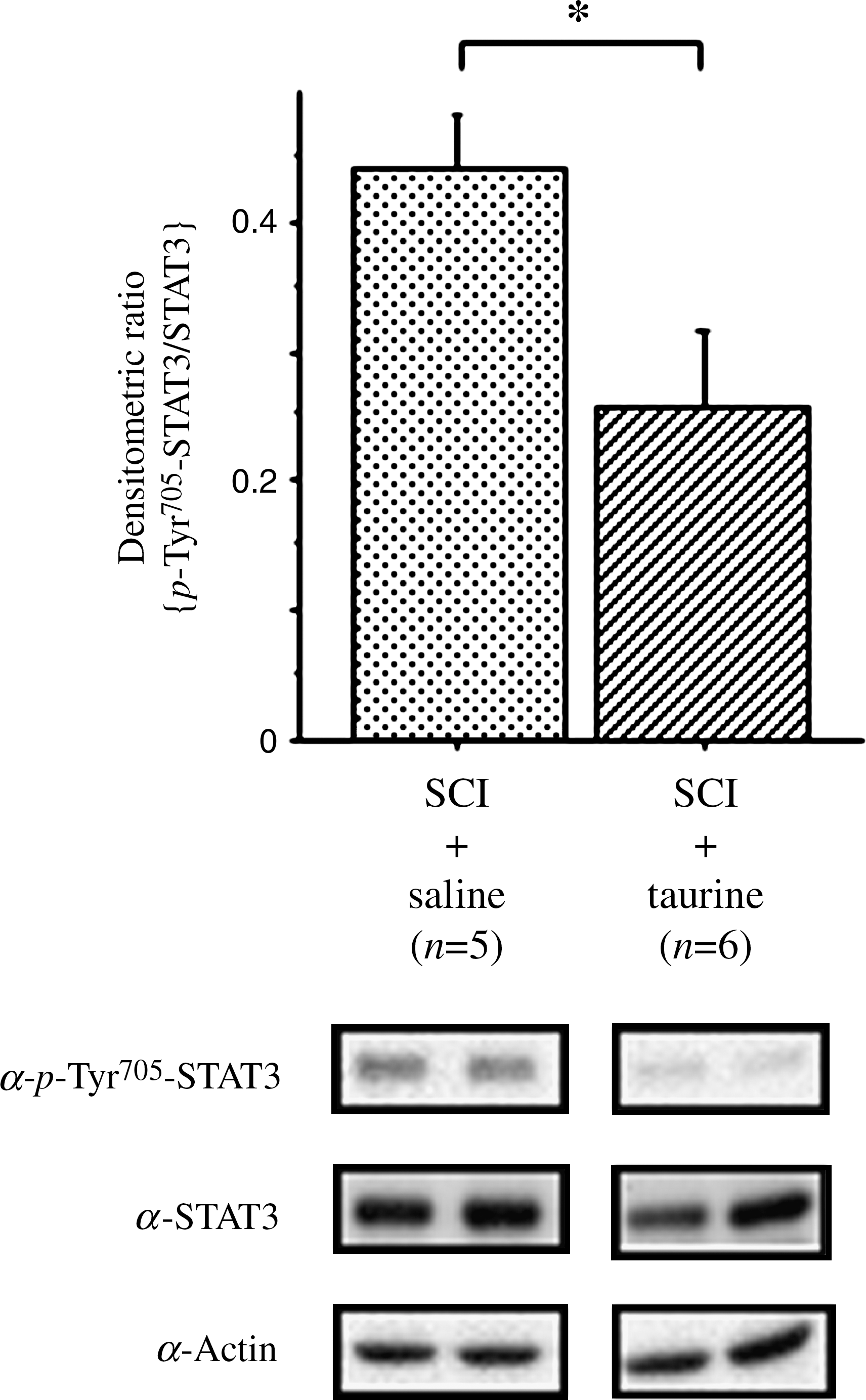
Effects of intraperitoneal injection of taurine on phosphorylation of the signal transducer and activator of transcription 3 (STAT3) at Tyr705. Mice were post-treated with taurine (250 mg/kg) or saline within 30 min after induction of spinal cord injury (SCI). Crude samples were subjected to Western blotting with anti-phosphorylated STAT3 at Tyr705 (α-p-Tyr705-STAT3), anti-STAT3 (α-STAT3), and actin (α-actin) antibodies. Two representative Western blots are shown in the lower panels. The histogram shows the amount of α-p-Tyr705-STAT3 relative to that of α-STAT3. Numbers of animals are given in parentheses. An asterisk indicates a significant difference by the Mann-Whitney U test (p < 0.05).
Expression of COX-2
Immunoblot analysis revealed constant detection of actin in the two groups, and a significant decrease in COX-2 after treatment with taurine was noted compared with the saline group (Fig. 3).
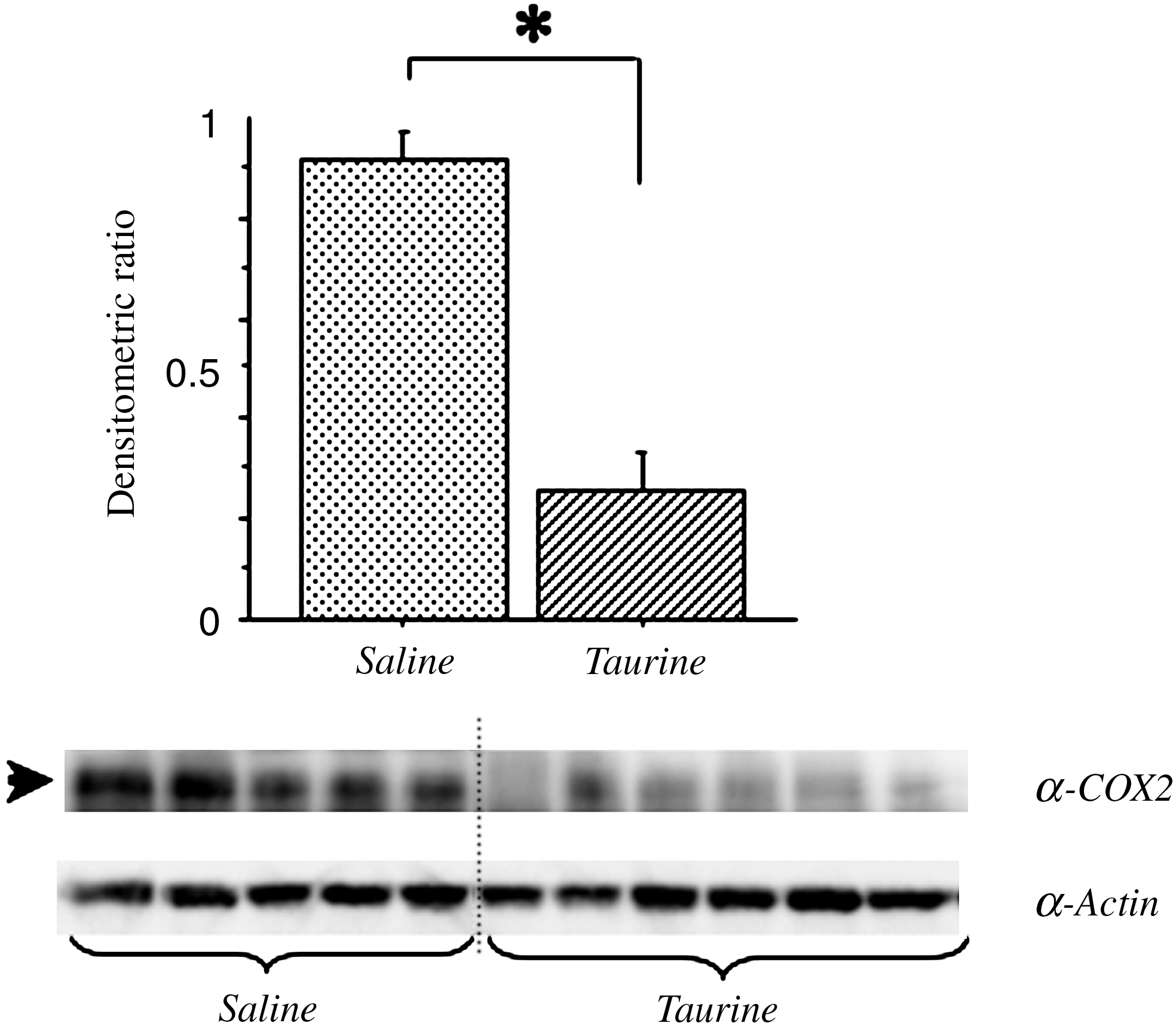
Immunoblot analysis of inducible cyclooxygenase-2 (COX-2) in injured spinal cord 6 h after severe spinal cord injury (SCI). Mice were treated with intraperitoneal injection of taurine (250 mg/kg; n = 6) or saline (n = 5) within 30 min after induction of severe SCI. Crude samples (25 μg of protein each) were subjected to Western blot analysis with anti-actin (α-actin) and anti-COX-2 (α-COX-2) antibodies. The histogram shows the amount of α-COX-2 relative to that of α-actin in the membrane. Mean ± SEM values are shown. Note the significant decrease in expression of COX-2 with taurine treatment. An asterisk indicates a significant difference by the Mann-Whitney U test (p < 0.05; SEM, standard error of the mean).
Effects of taurine on MPO after SCI
In intact control and injured tissues treated with saline at 6 h post-SCI, the mean concentrations of MPO in the spinal cord tissue were 3.2 ± 0.7 and 202 ± 16 ng/mL, respectively. Animals with laminectomy without compression treated with taurine had only slightly induced expression of MPO. Taurine caused significant decrease in a dose-dependent manner at 80, 250, and 800 mg/kg, compared to mice treated with saline (Fig. 4).
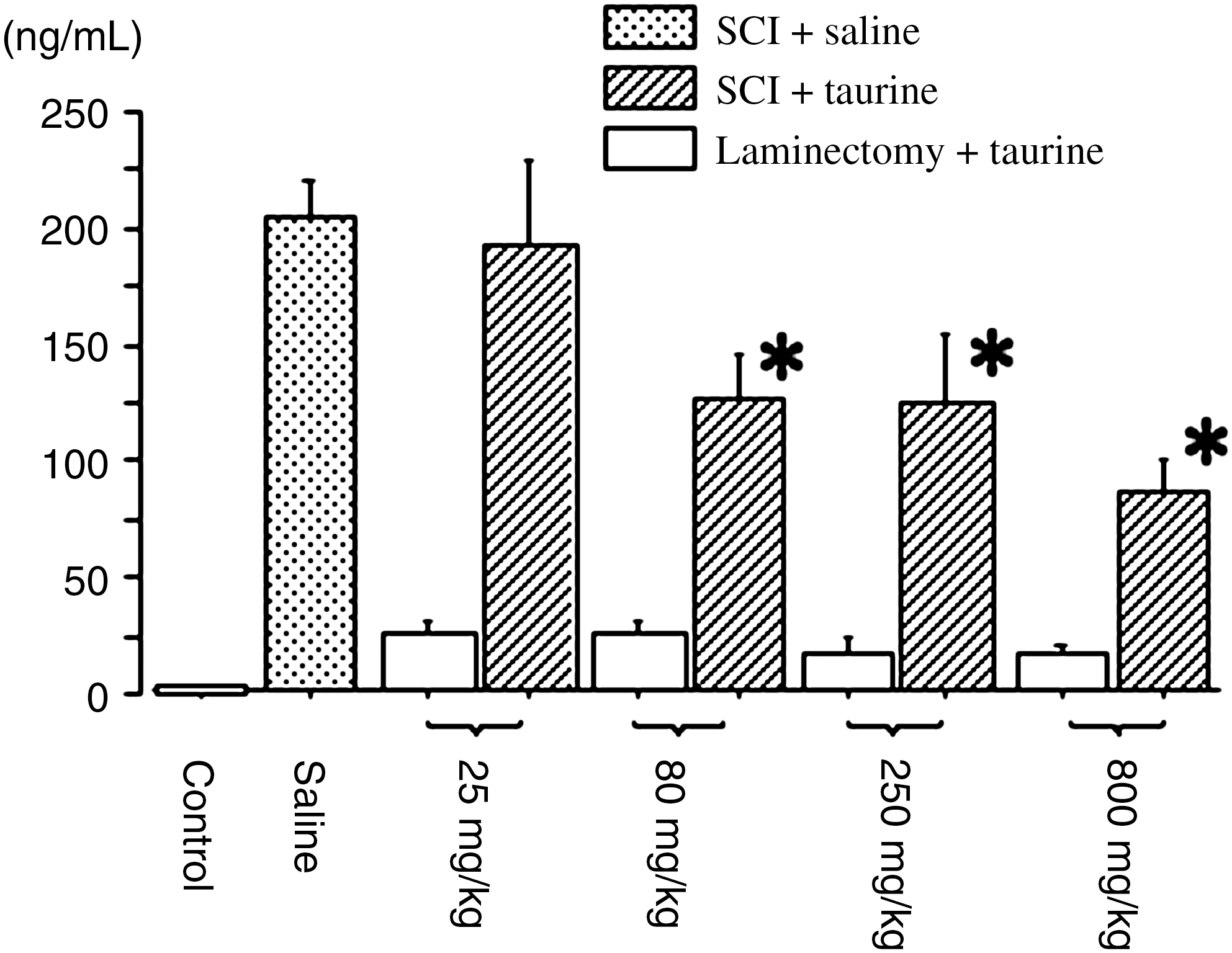
Myeloperoxidase (MPO) levels in the spinal cord 6 h after severe spinal cord injury (SCI) or sham surgery. Taurine was injected within 30 min after induction of severe SCI (n = 7 per group) or laminectomy without compression (n = 6 per group) at doses of 25, 80, 250, and 800 mg/kg. The MPO levels in spinal cord were measured using an ELISA kit. Mean ± SEM values of the data are shown (Saline, mice with severe SCI treated with saline [n = 7]; Control, intact control spinal cord). The MPO level in intact control spinal cord was 3.2 ± 0.7 ng/mL (n = 6; *p < 0.05 denotes a significant difference between saline and taurine treatment by analysis of variance followed by Fisher's post-hoc test; SEM, standard error of the mean; ELISA, enzyme-linked immunosorbent assay).
Histological observations
Histological examination of the injured spinal cord 6 h after SCI revealed intramedullary hemorrhage (Fig. 5A and B), with pronounced accumulation of neutrophils within subarachnoid spaces around the dorsal root nerve in mice treated with saline (Fig. 5C). This was reproducibly reduced by taurine treatment (250 mg/kg, Fig. 5D).

Mice subjected to severe spinal cord injury (SCI) were perfused with 10% formaldehyde in a phosphate-buffered solution 6 h after induction of SCI. Slices 10 μm thick were stained with hematoxylin and eosin. Mice were treated with saline (
At 4 weeks after severe SCI, H&E staining showed that normal gray matter in the lesion epicenter had been replaced by a connective tissue scar (Fig. 6A and C). Taurine treatment markedly attenuated these changes in the gray matter (Fig. 6B and D).

Mice subjected to severe spinal cord injury (SCI) were perfused with 10% formaldehyde in a phosphate-buffered solution 4 weeks after induction of SCI. Slices 10 μm thick were stained with hematoxylin and eosin. Mice were treated with saline (
Behavioral assessment
The BMS scores for hindlimb motor function of mice treated with saline improved from 0.7 ± 0.4 on day 1 to 6.7 ± 0.4 on day 28. The scores of mice treated with taurine demonstrated greater improvement, from 1.6 ± 0.4 on day 1 to 8.8 ± 0.2 on day 28 after SCI, the difference being significant after day 7 (Fig. 7).

Recovery of hindlimb motor function after spinal cord injury (SCI) was assessed using the Basso mouse scale (BMS) motor rating scale. Mice treated with taurine (n = 6; 250 mg/kg intraperitoneally within 30 min after induction of SCI, and then once a day from day 1 until day 7), or saline (n = 7 intraperitoneally on the same schedule) were examined for hindlimb motor function at 1, 2, 4, 7, 10, 14, 17, 21, 24, and 28 days after SCI. Mean ± SEM values of the data are shown. Asterisks indicate a significant difference between the two groups by the Mann-Whitney U test (p < 0.05; SEM, standard error of the mean).
Discussion
In the present exploration of the effects of taurine on local inflammatory responses and neurologic outcome after severe SCI, taurine significantly inhibited the expression of the proinflammatory cytokine IL-6, with an associated decrease in the phosphorylation of STAT3 at Tyr705 and expression of COX-2. Taurine also decreased the MPO level at the site of injury in a dose-dependent manner, and immunohistochemistry showed reduced neutrophil accumulation. Moreover, taurine clearly improved functional recovery in hindlimbs after SCI.
In the normal spinal cord the concentration of taurine in gray matter is significantly higher than that in white matter (Benton et al., 2001). Taurine-like immunoreactivity is most prominent in laminae I and II in the dorsal horn, suggesting an inhibitory neurotransmitter role (Lee et al., 1992). As an osmoregulator, taurine is well known to prevent brain swelling after cerebral ischemia or brain trauma (Pasantes-Morales and Schousboe, 1989). Taurine has multiple other functions, and is a regulator of Ca2+ influx with effects on neuromodulation and neuronal regeneration (Huxtable, 1992). After SCI, the concentration of taurine is reported to be temporarily elevated in acute stages, and then returns to basal levels (Diaz-Ruiz et al., 2007; Farooque et al., 1996; McAdoo et al., 1999). There has been at least one report of elevated concentrations of taurine in the chronic stages after SCI (Benton et al., 2001). The former is in line with the role of taurine as an osmoregulator, and the latter is associated with an increase in reactive astrocytes, which are known to contain large amounts of taurine (Lake, 1992; Lehmann and Hansson, 1987). These data thus suggest that taurine plays important roles in the complex cascade of the pathophysiological processes occurring after SCI.
The early inflammatory response after SCI may be initiated by neutrophils that infiltrate the lesion site after injury, and also into the subarachnoid space surrounding the cord (Carlson et al., 1998). P-selectin and cytokine-induced neutrophil chemoattractant-1 (CINC-1) are rapidly expressed on endothelial cell surfaces after SCI, where they mediate interactions between activated neutrophils and endothelial cells (Taoka et al., 1997; Tonai et al., 2001). The degree of neutrophil infiltration is generally assessed quantitatively by measuring MPO activity (Barone et al., 1991), and is proportional to the magnitude of trauma in spinal cord injury (Xu et al., 1990). Severe oxidative stress occurs within 1 h of SCI (Kamencic et al., 2001), and activated neutrophils release inflammatory mediators such as neutrophil elastase and reactive oxygen species, which damage adjacent endothelial vascular integrity, followed by increases in vascular permeability and secondary spinal cord injury. Prevention of neutrophil accumulation by anti-P-selectin monoclonal antibodies and neutrophil elastase inhibitors attenuates the motor disturbance seen after SCI in rats (Taoka et al., 1997, 1998). Taurine, through its antioxidant activity and regulation of intracellular calcium flux, can prevent endothelial cell dysfunction and cell death mediated by activated neutrophils (Wang et al., 1996b). Our results also revealed that taurine clearly prevented neutrophil accumulation, although the exact mechanism of this effect remains to be elucidated.
Taurine chloramine (TauCl) is produced from taurine by the halide-dependent myeloperoxidase (MPO) system, and traps chlorinated oxidants (HOCl). TauCl thus reduces HOCl toxicity and decreases production of inflammatory mediators such as TNF-α, nitric oxide, prostaglandin E2, IL-6, and superoxide anion (•O2–) (Kim et al., 1996; Kontny et al., 1999; Marcinkiewicz et al., 1995, 1998). One mechanism for this is through decreased activity of major transcriptional regulators of nuclear factor-κB and AP-1 (Georganas et al., 2000; Kontny et al., 2000). TauCl also attenuates the increased permeability of endothelium caused by neutrophil oxidants (Tatsumi and Fliss, 1994). Taurine inhibits at higher concentrations and to a lesser extent than TauCl (Kim et al., 1996; Marcinkiewicz et al., 1998). It may be that the effects of taurine are in fact partially attributable to the production of TauCl (Kim et al., 1996). Our finding that high doses of taurine are necessary to reduce the production of the proinflammatory cytokine IL-6 and MPO is in agreement with this conclusion.
In injured spinal cords proinflammatory cytokines, such as IL-1, IL-6, and TNF-α, are expressed at both messenger RNA (mRNA) and protein levels in injured spinal cords (Hayashi et al., 2000; Pan et al., 2002; Streit et al., 1998; Wang et al., 1996a). Immunoreactivity of IL-1β, IL-6, and TNF-α is observed in neurons as early as 0.5 h after human traumatic SCI (Yang et al., 2004). These proinflammatory cytokines activate secondary cytotoxic events and may be responsible for neural cell death. Transcriptional responses to IL-6-family cytokines involve the Janus kinase (JAK)-STAT signaling pathway (Heinrich et al., 1998), and we have previously shown that IL-6 is expressed after SCI with consequent STAT activation (Yamauchi et al., 2006). Phosphorylated STAT molecules translocate into the nucleus and bind to enhancer elements of target genes, leading to transcriptional activation. In cardiac late preconditioning, STAT3 plays an essential role in transcriptional activation of the COX-2 gene (Xuan et al., 2003). COX-2 is known to be induced in response to various inflammatory cytokines, such as lipopolysaccharide and IL-6, and they are found in endothelial cells of blood vessels 6 h after injury, but not in neurons, astrocytes, or microglia (Adachi et al., 2005). Selective inhibition of COX-2 is known to improve functional outcome following SCI (Resnick et al., 1998), and our results are in agreement with this finding. Taurine could clearly reduce the expression of COX-2 through lowered IL-6 production and JAK-STAT signal transduction in the injured spinal cord.
There are many experimental and clinical data suggesting protective roles of taurine against cardiac and cerebral ischemia. For example, it inhibited ischemia-induced apoptosis in cardiac myocytes through Akt-mediated caspase-9 inactivation (Takatani et al., 2004), and reduced lipoperoxidation and decreased reperfusion injury during coronary bypass surgery (Milei et al., 1992). Taurine modulates glutamate excitotoxicity through regulation of mitochondrial calcium homeostasis (El Idrissi and Trenkner, 1999; El Idrissi, 2008), which plays a neuroprotective role against ischemic insult. It may also protect neuronal cells via activation of both GABA and glycine receptors after cerebral ischemia (Wang et al., 2007). Inhibition of caspase-3-dependent apoptosis is also a neuroprotective mechanism of taurine after cerebral ischemia (Sun and Xu, 2008; Wang et al., 2007). Furthermore, taurine has a putative fetal brain neurotrophic function in the process of human brain development (Chen et al., 1998), and increases nerve growth in injured retinal explants (Lima et al., 1988). Other than anti-inflammatory effects, these various tissue-protective actions of taurine might play roles in preventing secondary damage and improving motor function after SCI.
There are very few clinically effective agents accepted for the treatment of SCI. Currently, only high doses of methylprednisolone are advocated (Bracken et al., 1990, 1997), and its neuroprotective effects are thought to be due to anti-inflammatory and immunosuppressive mechanisms. Interestingly, methylprednisolone treatment significantly increased spinal cord taurine levels after SCI in rats (Benton et al., 2001). Recently, Faulkner and colleagues (2004) reported that reactive astrocytes restrict inflammation, protect neurons and oligodendrocytes, and preserve motor function after SCI. Taurine is the most abundant free amino acid in astrocytes (Lake, 1992; Lehmann and Hansson, 1987).
To the best of our knowledge, this is the first demonstration of the anti-inflammatory effects of taurine in severe SCI. In light of our findings, taurine deserves consideration as a therapeutic multi-modality drug. Taurine has very strong hydrophilic and lipophobic characteristics. Further studies are necessary to explore the potential roles of taurine using more lipophilic taurine analogs (Gupta et al., 2005), and to ascertain the proper therapeutic time frame for the effective use of taurine to treat SCI.
Footnotes
Acknowledgments
We would like to express our gratitude to Dr. Malcolm Moore for critical reading of the manuscript.
Author Disclosure Statement
No conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.